漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析
2017-04-10王冬梅覃云斌吴林川
黄 端, 王冬梅, 任 远, 覃云斌, 吴林川
北京林业大学水土保持学院, 水土保持与荒漠化防治教育部重点实验室, 北京 100083
漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析
黄 端, 王冬梅*, 任 远, 覃云斌, 吴林川
北京林业大学水土保持学院, 水土保持与荒漠化防治教育部重点实验室, 北京 100083
以漓江水陆交错带为研究区,分两个条带分别量测了适生植物的5个叶性状指标:最大净光合速率(Amax)、比叶重(LMA)、单位质量叶片全氮含量(Nmass)、单位质量叶片全磷含量(Pmass)、单位质量叶片全钾含量(Kmass)。研究重度淹没带与微度淹没带不同功能型植物叶性状间的差异,分析并讨论重度淹没带叶性状间的关系与全球尺度是否存在差异,探究重度淹没带植物对水淹生境的生理响应机制。结果如下:(1)重度淹没带植物叶片的Amass、Nmass、Pmass显著高于微度淹没带。(2)乔木、灌木叶片的LMA均显著高于草本植物,而Amass、PPUE均显著低于草本植物。(3)重度淹没带草本叶性状指标的Nmass、Pmass、PNUE均显著高于微度微度淹没带,而乔木、灌木的叶性状在两个条带的差异则不显著。(4)重度淹没带植物叶性状关系与全球尺度基本一致,其植物叶片具有低LMA,高Amass、Nmass、Pmass。分析可知,重度淹没带植物在出露期提高叶片光合效率及相关营养水平可能是其适应水淹胁迫特殊生境的关键策略之一;不同功能型植物对同一环境的适应能力存在一定的差异,草本对于水淹环境的响应更为积极,适应能力更好;重度淹没带也存在叶经济谱,其植物在经济谱中属于“快速投资-收益”型物种。
漓江;水陆交错带;淹没带;叶性状关系;叶经济谱
叶片是植物体光合作用的主要器官, 是生态系统中初级生产者的能量转换器[1],针对特定环境其特性也会产生适应性变化[2]。叶片特性如形态结构、营养元素含量、光合能力等与植物碳同化、能量平衡密切相关[3],对植物的生长发育及其分布范围有重要影响。叶经济谱(Leaf economics spectrum)是指不同植物叶片的形态结构、养分含量、生理特征等性状具有相互依存、相互关联的内在联系[4],是具备协同变化关系的功能性状组合, 能够将一系列有规律连续变化的植物资源权衡策略数量化[5],叶经济谱对植物种进行了排序,从光合、呼吸速率低,叶片氮、磷含量低,叶片寿命长,比叶重高的“缓慢投资-收益”型物种(slow investment-return species)到光合、呼吸速率高,叶片氮、磷含量高,叶片寿命短,比叶重低的“快速投资-收益”型物种(fast investment-return species)[4,6- 7]。Wright等[4]研究了全球175个地区的219个科、2548个种的植物性状数据,提出全球植物叶经济谱,此后,国内外学者开展了大量相关研究,涉及中国东北温带植物[8]、中国海南热带山地雨林植物[9]、亚极地生态系统植物[10]、欧洲山地植物[11]等,均证实了叶经济谱的存在,然而关于水陆交错带生态系统植物的相关研究并不多见。
漓江位于广西壮族自治区,是喀斯特地貌的典型地带。漓江水陆交错带是漓江水体生态系统和陆地生态系统之间的界面区,植物需应对包括反复淹水在内的较为复杂的水文过程。目前针对水陆交错带植物的生理生态适应机制已有不少研究,但大多是对个别物种模拟水淹,不具备区域代表性,针对区域研究其植物功能性状可以反映该区域植被整体的生理生态过程特殊性,具有一定的灵敏性和代表性,是指示生态系统结构与功能的有效指标[12]。研究水陆交错带适生植物的叶性状及其经济谱,有利于探讨植物适应水陆交错带淹水生境的生理生态机制,为水陆交错带生态恢复提供理论依据。本研究试图以漓江水陆交错带主要适生植物为研究对象,研究目的在于:(1)探究在淹水胁迫的特殊生境下,重度淹没带植物叶性状间的关系与全球尺度下叶性状之间关系是否存在差异,验证植物叶经济谱存在与否;(2)研究重度淹没带与微度淹没带植物叶性状的差异;(3)研究水陆交错带植物叶性状在不同功能型植物间的差异。
1 研究区概况
漓江发源于广西兴安县、资源县交界处,流经兴安、灵川、桂林、阳朔,平乐,干流全长为214km,流域总面积为12 285 km2[13]。漓江流域(24°38′10″N至25°53′59″N, 110°07′39″E至110°42′57″E)为典型的喀斯特地貌,石灰石岩溶发育完整。本研究区位于漓江桂林-阳朔段水陆交错带,全长83km。该区域地处低纬度区,属湿润的中亚热带季风气候区,光照充足,年平均气温17.8—19.1℃,年降雨量1814—1941mm,年蒸发量1377—1857mm。年径流量十分丰富,但全年分布极为不均,3—8月为丰水期,径流量约为年径流量的80%,其中5—6月为年径流量的40%,9月到次年2月为枯水期,最枯月1月径流量仅为年径流量的2%。研究区主要乔木包括:枫杨(Pterocaryastenoptera)、乌桕(Sapiumsebiferum)、朴树(Celtissinensis)等,主要灌木包括荆条(Vitexnegundo)、一叶萩(Flueggeasuffruticosa)、水杨梅(Geumaleppicum)、夹竹桃(Neriumindicum)等,主要草本包括狗牙根(Cynodondactylon)、空心莲子草(Alternantheraphiloxeroides)、皱叶狗尾草(Setariaplicata)等。
2 研究方法
2.1 样地选择及研究物种
本研究于2015年5—6月沿漓江桂林-阳朔段选取了10个受人为干扰较小的典型水陆交错带样地。根据水文影响时间及相对高程[14-15],将研究区划分为重度淹没带及微度淹没带分别进行植物调查,其中重度淹没带年平均水淹胁迫历时大于7个月,相对高程(相对水平面)小于1m,微度淹没带年平均水淹胁迫历时小于2个月,相对高程大于1.4m。根据重度淹没带及微度淹没带植物群落的组成及结构特征,分别选取研究物种,并进行叶性状指标的测定。本次调查共选取43种植物,其中重度淹没带植物35 种,微度淹没带植物 39 种,重度淹没带和微度淹没带中相同的植物种有31 种。根据功能型划分为乔木、灌木、草本植物[16]。样地基本特征见表1,所取植物名录见表2。
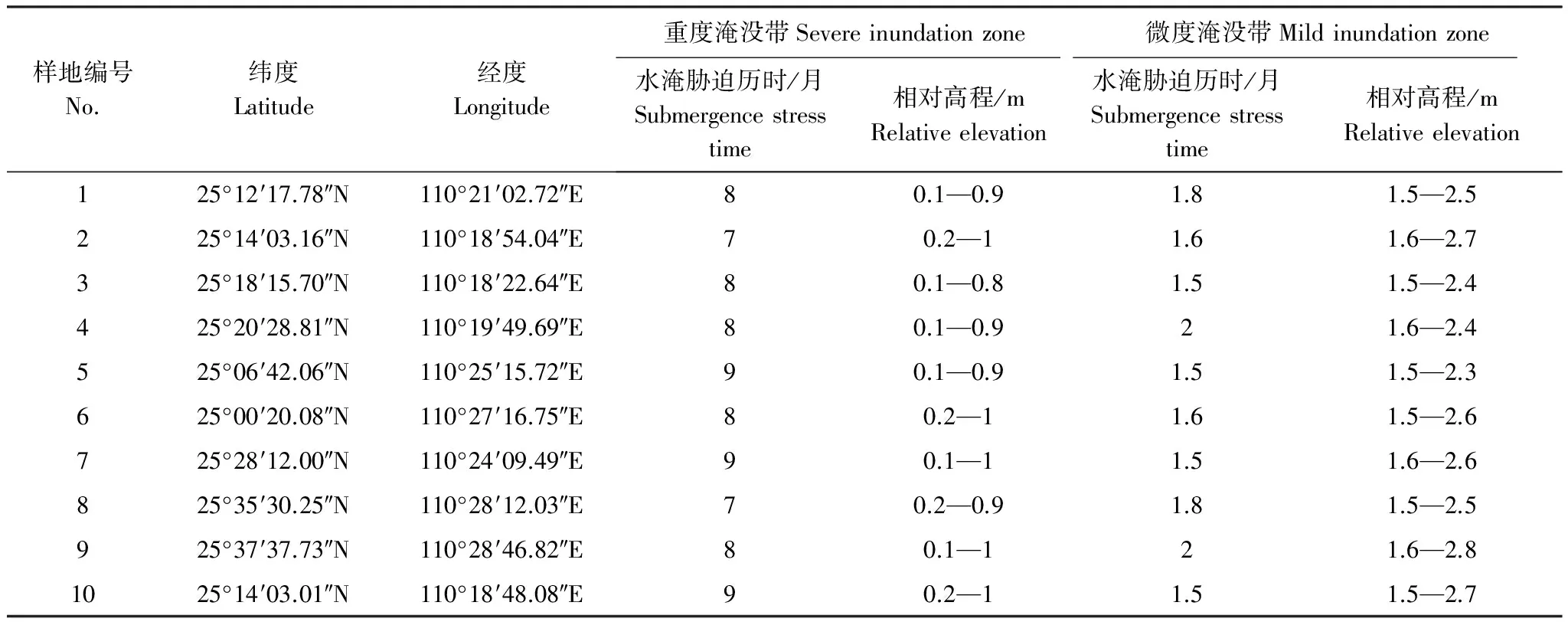
表1 样地基本特征

表2 调查植物名录
2.2 叶性状指标的测定
2.2.1 叶片气体交换参数的测定
在天气晴朗时进行野外植物叶片光合气体交换参数的测定。每种植物随机选取5—10株长势相近的成熟个体,每株植物选取3枚成熟的完全展开的未受损叶片,利用LI- 6400 便携式光合作用系统(LI-COR,Inc,USA),在天气晴朗的8:30—11:30先对叶片使用人工光源进行5—10min光合诱导,测得的光饱和速率作为最大净光合速率(Amax, μmol m-2s-1),同时测得气孔导度(GS)及胞间CO2浓度。其中,光源使用红、蓝光源,光强控制为 1 500μmol s-1m-2,叶温26—28℃,空气相对湿度50%—70%,CO2浓度(380±10)μmol s-1m-2,空气流速0. 5 L/min。计算单位质量最大净光合速率(Amass, μmol g-1s-1):Amass=Amax/LMA。
2.2.2 叶面积、干物质量及比叶重的测定
从测定气体交换参数的植株上采集10片完全展开的叶片带回实验室,用叶面积仪测量叶面积。测定结束后将叶片置于65℃烘箱中烘干48h,恒重后进行称重。计算比叶重,LMA=叶片干重(g)/叶面积(m2)。
2.2.3 叶片氮(Nmass)、磷(Pmass)、钾(Kmass)含量的测定
将采集的叶片样本烘干至恒重,碾碎后进行叶片全氮、全磷、全钾含量的测定。利用凯氏定氮法测定全氮含量Nmass(%),钼锑比色法测定全磷Pmass(%),火焰光度法测定全钾含量Kmass(%)。光合氮素利用效率PNUE(μmol g-1s-1) =Amass/Nmass,式中Amass为单位质量最大净光合速率,Nmass为单位质量叶氮含量。光合磷素利用效率PPUE(μmol g-1s-1)=Amass/Pmass,式中Amass为单位质量最大净光合速率,Pmass为单位质量叶氮含量。
2.3 数据处理与分析
采用单因素方差分析(One-Way ANOVA)分析各叶性状在重度淹没带、微度淹没带以及全球尺度 3 个点上的均值差异性,不同功能型物种叶性状间的差异;采用相关性分析及回归分析进行叶性状相互关系的参数估计。为了使数据整体符合正态分布,在处理过程中对叶性状指标进行了对数处理,所有数据分析均基于统计分析软件SPSS 17.0完成。
3 结果与分析
3.1 重度淹没带与微度淹没带共有物种叶性状的比较
由表3可知,重度淹没带与微度淹没带共有植物种31种,二者的LMA、PNUE、Kmass无显著差异;Amass、Nmass、Pmass、PPUE重度淹没带均显著高于微度淹没带(P=0.000,P=0.029,P=0.040,P=0.001)。
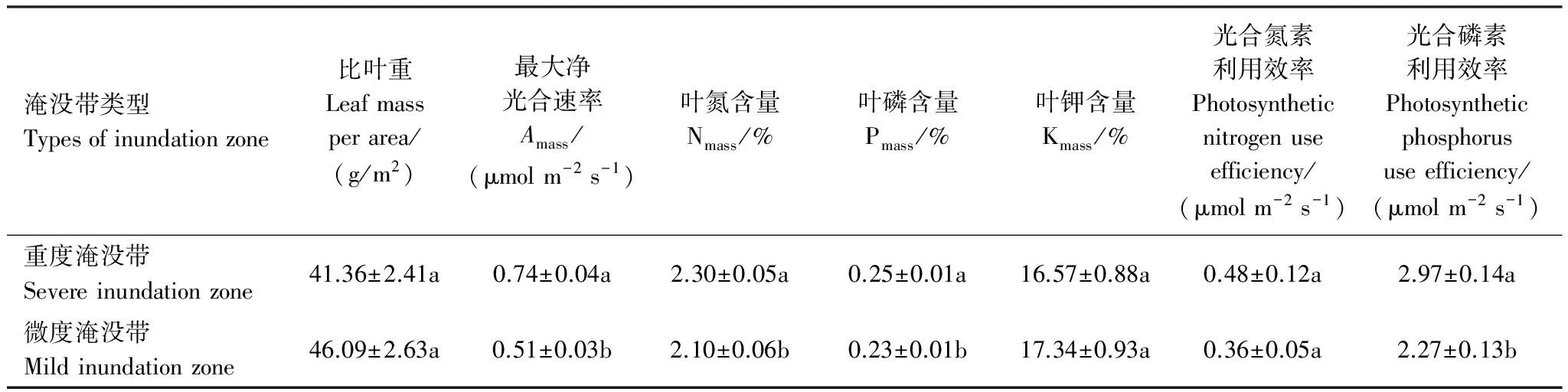
表3 重度淹没带与微度淹没带叶性状均值比较
表中数据为平均值±SE,不同小写字母表示在P<0.05水平下差异显著
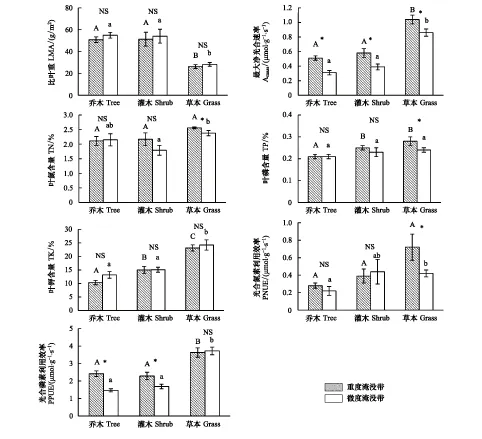
图1 重度淹没带与微度淹没带不同功能型植物叶性状比较Fig.1 Comparison among leaf traits of different plant functional types between severe and mild inundation zones不同大写/小写字母表示在P<0.05水平下差异显著;*表示重度淹没带与微度淹没带之间有显著差异,NS表示无显著性差异
3.2 重度淹没带与微度淹没带不同功能型植物叶性状的比较
由图1可知,对于重度淹没带而言,乔、灌木的LMA、Amass、PPUE较接近,但与草本植物差异显著,Kmass比较结果为:乔木<灌木<草本,且差异显著,乔木的Pmass显著低于灌木及草本植物。对于微度淹没带而言,草本的LMA、Amass、Kmass,PPUE与灌木、乔木均有显著差异。Nmass的比较结果为:灌木<乔木<草本,其中灌木与草本差异显著,PNUE的比较结果为:乔木<草本<灌木,其中乔木与草本差异显著。
图1还表明,两个条带的乔木、灌木、草本的Amass比较结果均为重度淹没带显著高于微度淹没带,而仅草本植物的Nmass、Pmass、PNUE在两个条带间有显著差异性。
3.3 重度淹没带植物叶片各性状之间的关系
由表4可知,重度淹没带植物叶片LMA与Amass、Nmass、Pmass、Kmass、PNUE、PPUE均呈现显著的负相关关系,Amass与Nmass、Pmass、Kmass、PNUE、PPUE均呈现显著的正相关关系。PNUE与PPUE呈显著的正相关关系。由表5可知,LMA、Amass、Pmass三者之间的斜率与1或-1没有显著差异。
3.4 重度淹没带与全球尺度叶经济性状的比较
如表6所示,重度淹没带植物叶片LMA为41.36 g/m2,Amass为0.74μmol g-1s-1,Nmass为2.30%,Pmass为0.25%,而在全球尺度下,植物叶片各经济性状值分别为:LMA127.70 g/m2,Amass为0.13μmol g-1s-1,Nmass为1.94%,Pmass为0.11%。分析可得,重度淹没带植物叶片的LMA显著低于全球水平,而Amass、Nmass、PNUE、Pmass、PPUE均显著高于全球水平。
4 讨论
4.1 重度淹没带与微度淹没带植物叶性状的差异性
研究结果表明,重度淹没带植物的叶性状与微度淹没带存在一定的差异。重度淹没带植物叶片净光合速率显著高于微度淹没带,说明重度淹没带植物在出露期通过提高自身的光合能力来快速合成积累碳水化合物,以应对水淹胁迫,这与郭泉水等[17]等对香根草、裴顺祥等[18]对狗牙根研究得出淹没区段植物在出露期能加强光合作用的结论一致。类似于揭胜麟[12]对三峡水库消落带植物的研究,重度淹没带植物长期处于淹水胁迫的复杂生境中,在露出水面的短期时间内通过提高自身的光合能力及相关的叶片营养物质含量,快速合成更多的光合产物并在根系进行储存,以应对深度淹水。植株体内的碳水化合物含量被发现与植株的耐淹程度及淹水后的恢复生长呈显著正相关[19-20],在淹水胁迫条件下,光合作用产物更多的会被分配到植物的根系[21]。经历水淹胁迫但在陆生条件下仍保持较高的光合速率是植物耐受水淹重要原因之一[18]。
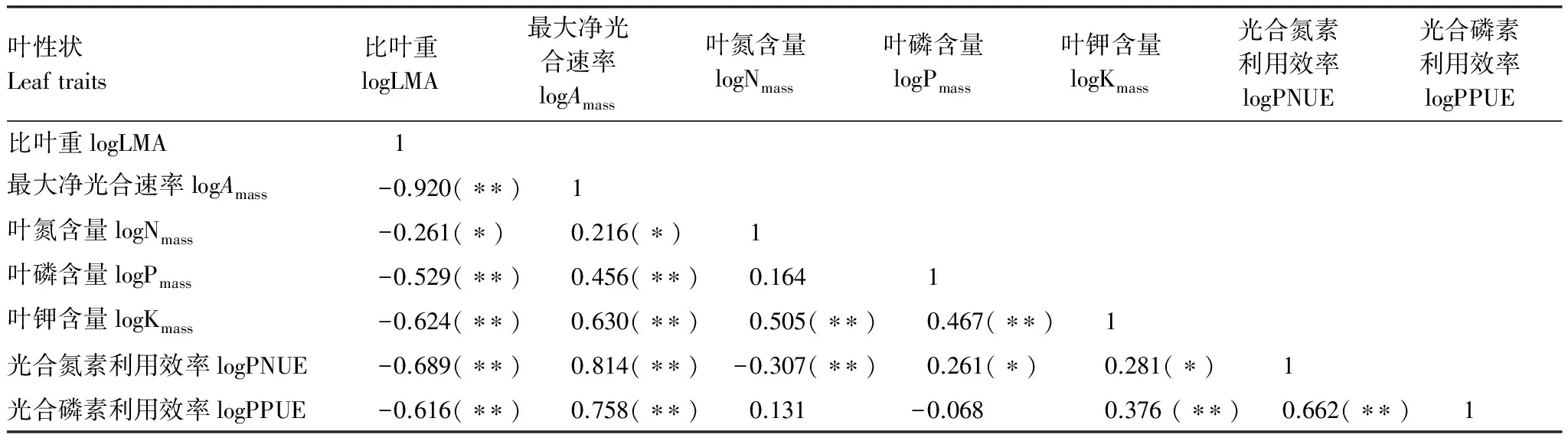
表4 重度淹没带植物叶性状之间的相关性分析
*表示在P<0.05 水平(双侧)上显著相关;**表示在P<0.01水平(双侧)上显著相关
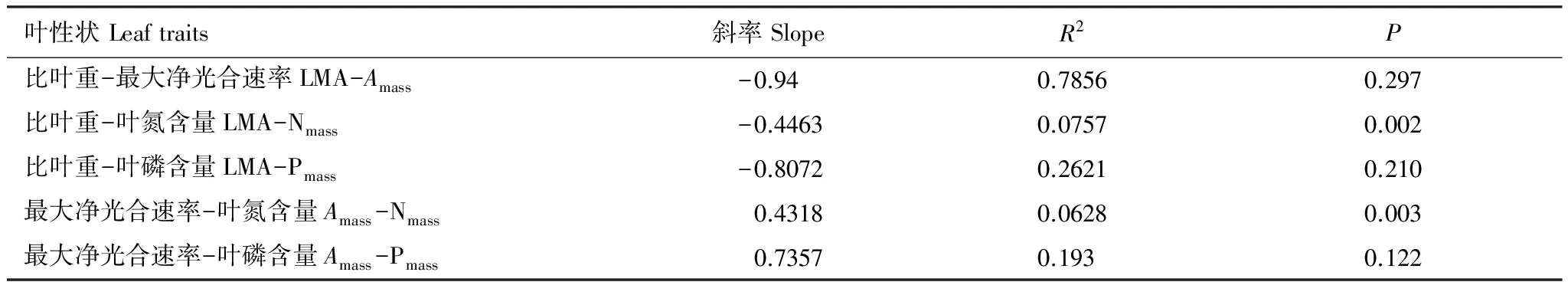
表5 重度淹没带叶性状关系斜率分析
表中P值反映回归分析斜率与1或-1是否有显著差异

表6 重度淹没带与全球尺度下叶性状均值比较
表中数据为平均值±SE,不同小写字母表示在P<0.05水平下差异显著
重度淹没带植物叶片的Nmass、Pmass、PPUE均显著高于微度淹没带。有研究表明[22],水淹土壤中氮、磷元素的有效性升高,有利于植物在缺氧环境下对氮、磷的吸收,重度淹没带植物可能通过增加出露期对土壤中氮及磷的吸收来提高自身的光合作用能力,促进有机物质的积累,增加整个植株生命活性和抗逆能力。植物的光合作用能力与其叶片的营养元素含量密切相关。氮是叶绿素的主要组成部分[16,23];磷是植物体内核酸及贮存能量的重要组成部分,也是光合过程中辅酶的成分之一,它直接参与光合过程中的光合磷酸化及碳同化过程,与ATP的合成、磷酸丙糖的转运及光合产物的运输密切相关[24]。朱根海等[25]总结了诸多学者对水稻、烟草、羊草等植物的研究,得出提高叶片的含氮量能增加叶片的光合能力。Brooks A等[26]、穆宏平等[27]对菠菜、朱砂根、山血丹的研究表明,在一定程度上增加植物叶片中的磷浓度能提高羧化效率从而提高植物的光合能力。
4.2 不同功能型植物叶性状的比较
本研究中,重度淹没带与微度淹没带的乔木、灌木叶片的Amass、PPUE均显著低于草本植物,而LMA显著高于草本植物,这说明不同功能型植物对同一环境的适应能力存在一定的差异[28],反映出草本植物具有更薄的叶片,更高的光合能力。这主要是由于不同功能型的植物对于叶氮的分配策略不同,乔木与灌木叶片主要将氮用于增加叶片厚度,增强叶片韧性,增大叶肉细胞密度,以防止强光灼伤或过度失水[29-33],因此其光合器官能分配到的氮素较为有限,以致其光合作用能力较弱,而草本植物叶片分配更多的氮到Rubisco中,提高光合速率,加快生长[4,33-34]。
重度淹没带草本叶片的Nmass、Pmass、PNUE均显著高于微度淹没带,灌木及乔木仅PPUE显著高于微度淹没带,这说明草本植物对于水淹胁迫的响应更为积极,对于涝渍环境的适应性更好。此外,根据野外调查结果,重度淹没带常见植物种为草本植物,其次是灌木,偶见乔木,这也能从实际情况反映草本植物能更好的适应水淹。
4.3 重度淹没带植物叶性状相互关系及与全球尺度的比较
重度淹没带植物叶性状与全球尺度平均水平均差异显著(见表6),重度淹没带植物叶片的Amass、Nmass、PNUE、Pmass、PPUE分别是全球尺度的5.69、1.19、8.00、2.27、2.80倍,而全球尺度下叶片的LMA则是重度淹没带的3.09倍。重度淹没带草本植物共计23种,占调查物种总数的65.7%,全球尺度下研究的草本植物共计633种,仅占研究总物种数的26.7%,草本植物叶片的LMA要显著低于乔木及灌木(图1),因此植物功能型的差异可能是导致LMA不同的原因之一。在水淹环境下,是否具备较高的光合速率及叶营养元素是植物生存与否的一个决定性因素[35],所以重度淹没带植物叶片的高Amass及高叶营养元素可能是其适应水淹生境的策略之一。
本研究还发现,重度淹没带植物叶片的Amass、Nmass、Pmass、Kmass、PNUE、PPUE均与LMA呈显著的负相关关系,而Nmass、Pmass、Kmass、PNUE、PPUE均与Amass呈显著的正相关关系,说明叶片越薄,其叶片营养越高含量越高,光合能力越强,这与前人研究结果一致[2,6,9]。此外,叶片LMA、Amass与Pmass三者之间的斜率与1或-1没有显著差异,说明LMA的增长速度同Amass、Pmass的下降速度相当,Amass与Pmass增长或减少的比例相等。Nmass与LMA、Amass之间呈现异速生长关系,Amass增加的速度要小于Nmass增加的速度(slope=0.4318),LMA下降的速度小于Nmass增加的速度(slope=-0.4463)(表5)。重度淹没带与全球尺度下植物的成对叶性状相关关系基本相同,这说明全球叶经济谱规律也存在于环境条件特殊的漓江水陆交错带,通过分析可知,重度淹没带植物在叶经济谱中属于光合速率高,叶片氮、磷含量高,比叶重低的“快速投资-收益”型。
然而有研究指出,植物叶性状相关关系中,N、P作为植物生长限制元素共同参与植物基本的生理生化过程,其含量之间呈显著正相关[36-37],本研究重度淹没带植物叶片Nmass与Pmass也呈现正相关,但并不显著。这可能是由于漓江喀斯特地貌特殊的气候条件、重度淹没带频繁受水文干扰的特殊生境、群落类型的差异或植物的遗传差异所致,具体原因可在后续研究中设计实验进行研究。
总之,本研究以漓江水陆交错带植被为研究对象,分析并验证了水淹胁迫生境下叶经济谱的存在。研究发现受水淹胁迫历时较长的重度淹没带植物叶片具有低LMA,高Amass、Nmass、Pmass,具备典型的速生种的特征,属于“快速投资-收益”型物种。重度淹没带植物会采取叶片功能性状间的权衡策略来适应长期受水淹胁迫的不利环境,这为遏制河岸带植被退化、加速生态修复、保持可持续发展提供了重要的理论基础。此外,研究发现草本植物对于淹水环境有更好的适应性,可推测采取人工措施进行植被恢复时,应优先将资源投资于草本植物及速生种。
5 结论
(1)相比微度淹没带,重度淹没带植物叶片具备更高的Amass、Nmass、Pmass。植物在出露期迅速提高叶片光合效率及相关营养水平,以抵御淹没期的深度水淹,这可能是其适应水陆交错带特殊生境的关键策略之一。
(2)不同功能型植物对同一环境的适应能力存在一定的差异,乔木、灌木叶片的LMA较为接近,且均显著高于草本植物,而Amass、PPUE均显著低于草本植物。相对乔木、灌木而言,草本植物对于水淹环境的响应更为积极。重度淹没带乔木、灌木的叶片性状指标与微度淹没带的较为接近,不具备显著差异,而草本植物的Nmass、Pmass、PNUE则显著高于微度淹没带的相应值,在进行重度淹没带植被恢复的过程中宜选择草本植物作为先锋物种。
(3)尽管重度淹没带受水淹胁迫影响较大,但其叶片性状的相互关系与全球尺度基本一致,叶经济谱依然存在。重度淹没带植物叶片具有低LMA,高Amass、Nmass、Pmass,属于经济谱中的“快速投资-收益”型物种。
(4)在对重度淹没带采取人工措施进行植被恢复时,要考虑到植物对水淹环境的适应性,建议可优先将资源投资于草本植物及速生灌木植物,以保证植物的存活率及恢复效果。
[1] 张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844- 852.
[2] 罗璐, 申国珍, 谢宗强, 喻杰. 神农架海拔梯度上4种典型森林的乔木叶片功能性状特征. 生态学报, 2011, 31(21): 6420- 6428.
[3] Ackerly D D, Knight C A, Weiss S B, Barton K, Starmer K P. Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns in species level and community level analyses. Oecologia, 2002, 130(3): 449- 457.
[4] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821- 827.
[5] 陈莹婷, 许振柱. 植物叶经济谱的研究进展. 植物生态学报, 2014, 38 (10): 1135- 1153.
[6] Reich P B, Walters M B, Ellsworth D S. From tropics to tundra: global convergence in plant functioning. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(25): 13730- 13734.
[7] Read Q D, Moorhead L C, Swenson N G, Bailey J K, Sanders N J. Convergent effects of elevation on functional leaf traits within and among species. Functional Ecology, 2014, 28(1): 37- 45.
[8] 唐艳. 东北主要温带植物叶性状的种间差异及其相关关系[D]. 哈尔滨: 东北林业大学, 2011.
[9] 刘福德, 王中生, 张明, 王文进, 安树青, 郑建伟, 杨文杰, 张世挺. 海南岛热带山地雨林幼苗幼树光合与叶氮、叶磷及比叶面积的关系. 生态学报, 2007, 27(11): 4651- 4661.
[10] Freschet G T, Cornelissen J H C, van Logtestijn R S P, Aerts R. Evidence of the ‘plant economics spectrum’ in a subarctic flora. Journal of Ecology, 2010, 98(2): 362- 373.
[11] Grigulis K, Lavorel S, Krainer U, Legay N, Baxendale C, Dumont M, Kastl E, Arnoldi C, Bardgett R D, Poly F, Pommier T, Schloter M, Tappeiner U, Bahn M, Clément J-C. Relative contributions of plant traits and soil microbial properties to mountain grassland ecosystem services. Journal of Ecology, 2013, 101(1): 47- 57.
[12] 揭胜麟, 樊大勇, 谢宗强, 张想英, 熊高明. 三峡水库消落带植物叶片光合与营养性状特征. 生态学报, 2012, 32(6): 1723- 1733.
[13] 李扬, 王冬梅, 信忠保. 漓江水陆交错带植被与土壤空间分异规律. 农业工程学报, 2013, 29(6): 121- 128.
[14] 李扬. 漓江水陆交错带水文-土壤-植被相互作用机制及植被恢复研究[D]. 北京: 北京林业大学, 2014.
[15] 李扬, 王冬梅, 信忠保, 王晶, 任远, 李青山. 漓江水陆交错带不同淹没区植物多样性与土壤特征研究. 生态学报, 2015, 35(15): 5121- 5130.
[16] 郑淑霞, 上官周平. 不同功能型植物光合特性及其与叶氮含量、比叶重的关系. 生态学报, 2007, 27(1): 171- 181.
[17] 郭泉水, 洪明, 裴顺祥, 王祥福, 聂必红, 杨永明, 梁洪海. 香根草形态性状和光合特性对三峡库区消落带水陆生境变化的响应. 西北植物学报, 2012, 32(11): 2328- 2335.
[18] 裴顺祥, 洪明, 郭泉水, 秦爱丽, 朱莉, 朱妮妮. 三峡库区消落带水淹结束后狗牙根的光合生理生态特性. 生态学杂志, 2014, 33(12): 3222- 3229.
[19] Panda D, Sharma S G, Sarkar R K. Chlorophyll fluorescence parameters, CO2photosynthetic rate and regeneration capacity as a result of complete submergence and subsequent re-emergence in rice (OryzasativaL.). Aquatic Botany, 2008, 88(2): 127- 133.
[20] Kawano N, Ito O, Sakagami J I. Relationship between shoot elongation and dry matter weight during submergence inOryzasativaL. and O. glaberrima Steud. rice cultivars. Plant Production Science, 2008, 11(3): 316- 323.
[21] 汪贵斌, 曹福亮, 张晓燕, 张往祥. 涝渍胁迫对不同树种生长和能量代谢酶活性的影响. 应用生态学报, 2010, 21(3): 590- 596.
[22] 金茜, 王瑞, 周向睿, 周志宇, 卢鑫, 赵萍, 李金辉, 周媛媛. 水淹胁迫对紫穗槐生长及营养元素积累的影响. 草业科学, 2013, 30(6): 904- 909.
[23] 曹翠玲, 李生秀, 苗芳. 氮素对植物某些生理生化过程影响的研究进展. 西北农业大学学报, 1999, 27(4): 96- 101.
[24] 于海秋, 彭新湘, 严小龙, 曹敏建. 缺磷对不同磷效率基因型大豆光合日变化的影响. 沈阳农业大学学报, 2005, 36(5): 519- 522.
[25] 朱根海, 张荣铣. 叶片含氮量与光合作用. 植物生理学通讯, 1985, (2): 9- 12.
[26] Brooks A, Woo K C, Wong S C. Effects of phosphorus nutrition on the response of photosynthesis to CO2and O2, activation of ribulose bisphosphate carboxylase and amounts of ribulose biphosphate and 3-phosphoglycerate in spinach leaves. Photosynthesis Research, 1988, 15(2): 133- 141.
[27] 穆宏平, 叶万辉, 陈贻竹, 朱建玲, 曹洪麟. 不同磷营养水平对朱砂根和山血丹光合作用及生长的影响. 武汉植物学研究, 2008, 26(5): 514- 519.
[28] 郑淑霞, 上官周平. 黄土高原地区植物叶片养分组成的空间分布格局. 自然科学进展, 2006, 16(8): 965- 973.
[29] Warren C R, Adams M A. Evergreen trees do not maximize instantaneous photosynthesis. Trends Plant Science, 2004, 9(6): 270- 274.
[30] Onoda Y, Hikosaka K, Hirose T. Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Functional Ecology, 2004, 18(3): 419- 425.
[32] Coble A P, Cavaleri M A. Light drives vertical gradients of leaf morphology in a sugar maple (Acersaccharum) forest. Tree Physiology, 2014, 34(2): 146- 158.
[33] Hikosaka K, Terashima I. Nitrogen partitioning among photosynthetic components and its consequence in sun and shade plants. Functional Ecology, 1996, 10(3): 335- 343.
[34] Galmés J, Flexas J, Keys A J, Cifre J, Mitchell R A C, Madgwick P J, Haslam R P, Medrano H, Parry M A J. Rubisco specificity factor tends to be larger in plant species from drier habitats and in species with persistent leaves. Plant, Cell & Environment, 2005, 28(5): 571- 579.
[35] Domingues T F, Meir P, Feldpausch T R, Saiz G, Veenendaal E M, Schrodt F, Bird M, Djagbletey G, Hien F, Compaore H, Diallo A, Grace J, Lloyd J. Co-limitation of photosynthetic capacity by nitrogen and phosphorus in West Africa woodlands. Plant, Cell & Environment, 2010, 33(6): 959- 980.
[36] 刘超, 武娴, 王襄平, 尹伟伦, 张淑静. 内蒙古灌木叶性状关系及不同尺度的比较. 北京林业大学学报, 2012, 34(6): 23- 29.
[37] Reich P B, Oleksyn J, Wright I J, Niklas K J, Hedin L, Elser J J. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes. Proceedings of the Royal Society of London, Series B, Biological Sciences, 2010, 277(1683): 877- 883.
Responses of leaf traits to submergence stress and analysis of the economic spectrum of plant species in an aquatic-terrestrial ecotone, the Li River
HUANG Duan, WANG Dongmei*, REN Yuan, QIN Yunbin, WU Linchuan
SoilandWaterConservationCollegeofBeijingForestryUniversity,Statekeylaboratoryofsoilandwaterconservationanddesertificationprevention,Beijing100083,China
Five leaf traits were analyzed in different plant species in an aquatic-terrestrial ecotone, Li River, to investigate differences in these traits between different types of functional plants in severe and mild inundation zones, and to explore the physiological responses of plants suffering from long-term submergence stress. By analyzing the relationships between leaf traits in plants subjected to severe inundation, differences between the results of this study and those on the global scale are also discussed. The five leaf traits analyzed were: leaf mass per area (LMA), leaf maximum net photosynthesis rate (Amax), leaf nitrogen content per mass (Nmass), leaf phosphorus content per mass (Pmass), and leaf potassium content per mass (Kmass). The results were as follows: (1) The values ofAmass, Nmass, and Pmassunder severe submergence were significantly higher than those under mild submergence. (2) The LMA value for grasses was relatively lower than that for trees and shrubs, whereasAmaxand PNUE (photosynthetic nitrogen use efficiency) were higher for grasses. (3) The Nmass, Pmass, and PNUE values for grasses in the severe inundation zone were significantly higher than those in the mild inundation zone, while there were no significant differences in leaf traits for trees and shrubs between the two zones. (4) The relationships between leaf traits in the severe inundation zone were similar to those on the global scale, and the species in the severe inundation zone had lower LMA, higherAmass, Nmass, and Pmass. These results suggest that improving photosynthetic capacity and increasing the levels of relevant leaf nutrients may be involved in the adaptation of plants to aquatic-terrestrial ecotone conditions. Compared with trees and shrubs, grasses showed stronger adaptability to submergence, which illustrates the differences in adaptability between different types of functional plants. The results of this study indicate that a spectrum of leaf economics also exists in plant species in the severe inundation zone, and that these represent species with a fast investment-return on the leaf economic spectrum.
Li River; aquatic-terrestrial ecotone; inundation zone; leaf traits relationships; leaf economics spectrum
国家“十二五”科技支撑计划项目(2012BAC16B03)
2015- 08- 28;
日期:2016- 06- 14
10.5846/stxb201508281789
*通讯作者Corresponding author.E-mail: dmwang@126.com
黄端, 王冬梅, 任远, 覃云斌, 吴林川.漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析.生态学报,2017,37(3):750- 759.
Huang D, Wang D M, Ren Y, Qin Y B, Wu L C.Responses of leaf traits to submergence stress and analysis of the economic spectrum of plant species in an aquatic-terrestrial ecotone, the Li River.Acta Ecologica Sinica,2017,37(3):750- 759.
