白细胞介素13对胆管成纤维细胞TGF-β1/Smads通路表达的影响及地塞米松的干预作用
2017-04-10李克跃石承先汤可立魏国微刘振华黎涛张帅民徐贤刚
李克跃,石承先,汤可立,魏国微,刘振华,黎涛,张帅民,徐贤刚
(贵州省人民医院 肝胆外科,贵州 贵阳 550002)
良性胆道狭窄(benign biliary stricture,BBS)是由医源性胆道损伤等良性疾病引起的胆管腔疤痕性缩窄,常表现为胆管炎、肝功能不全等并发症,严重威胁患者健康及生命[1],近年来发病率逐渐增高。报道[2-4]显示胆管成纤维细胞大量增殖并转化为肌成纤维细胞、转化生长因子β1(transforming growth factor-β1,TGF-β1)/Smads通路的异常表达在胆管疤痕修复及BBS形成过程中均发挥了重要作用。药物是疤痕治疗的选项之一,令人遗憾的是目前临床上依旧缺乏一种特效、可靠的药物[5]。白细胞介素13(interleukin 13,IL-13)具有促进成纤维细胞I型胶原表达的作用[6-7],而胶原的过度沉积是BBS及胆管疤痕的主要生物学特征之一。地塞米松(Dexamethasone,Dex)已被用于治疗皮肤疤痕性疾病[8-9]。但IL-13对胆管成纤维细胞的作用尚不清楚,Dex对IL-13处理的胆管成纤维细胞的作用也不清楚。本研究对IL-13干预的兔胆管成纤维细胞使用Dex处理,观察TGF-β1/Smads通路的表达及细胞增殖水平的变化。
1 材料与方法
1.1 材料
胎牛血清(fetal bovine serum,FBS)、达尔伯克改良伊格尔培养基(dulbecco's modified eagle medium,DMEM)、CCK-8细胞计数盒、β-actin引物、TRIzol、全蛋白提取试剂盒、UNIQ-10柱式TRIZOL总RNA提取试剂盒、反转录试剂盒(上海生工生物工程有限公司);TGF-β1、Smad3及Smad4引物(上海Introgen公司);TGF-β1上游引物序列为:5'-GGC TCA CCT TCT GCC CGT CT-3',下游引物序列为:5'-GTC TCG GTA TCC CAC GAA AGA AAC G-3';Smad3上游引物序列为:5'-CCA GTT CTA CCT CCT GTG CTG-3',下游引物序列为:5'-GGG GTC TCT GGA ATA TTG CTC-3';Smad4上游引物序列为:5'-CGC GGA TCA ACC GAG ACA TAT ACT-3',下游引物序列为:5'-GGC AGG CTG ACT TGT G-3';β-actin上游引物序列为:5'-CTC TCC ACC TTC CAG CAG AT-3',下游引物序列为:5'-TGG CTC TAA CAG TCC GCC TA-3';抗-Smad4单克隆抗体(ab187094)及抗-TGFβ1单克隆抗体(ab99562)(英国abcam公司);抗-细胞角蛋白单克隆抗体(C-1801)、抗-波形蛋白单克隆抗体(V2258)、IL-13(编号I1896)、PVDF膜(美国sigma公司);HRP标记的第二抗体及抗-β-actin单克隆抗体(北京全式金生物技术有限公司);电化学发光(electrochemical luminescence,ECL)试剂盒(美国Millipore公司);SYBR®Premix Ex Taq™(Tli RNaseH Plus)(大连宝生物工程有限公司);DAPI(瑞士Roche公司);地塞米松磷酸钠注射液(武汉华中药业有限公司,批号:20150204);家兔2只(购自贵州医科大学实验动物中心)。
1.2 动物模型的建立、细胞的获取及细胞鉴定
清洁级家兔2只,术前12 h禁食。耳缘静脉注射2.5%戊巴比妥钠(40~45 mg/kg)麻醉,取上腹部正中4~6 cm长切口,取出约2 cm长胆总管后用深麻醉处死动物。将胆总管用无菌磷酸盐缓冲溶液(PBS)清洗3次,剪为约2~3 mm3大小,然后转移到细胞培养瓶中并加入少量含有20%(v/v)FBS、100 mg/mL链霉素、100 U/mL青霉素的DMEM培养液放入37 °C、5%CO2的培养箱中培养[10]。使用差速贴壁法分离纯化细胞,细胞生长至80%左右时传代。对第3代细胞用细胞免疫荧光法检测其波形蛋白及细胞角蛋白抗体的表达进行成纤维细胞的鉴定[10-11]。本实验所用细胞为第3~5代成纤维细胞。
1.3 实验分组及各组细胞的处理
分为如下:空白对照;IL-13组(IL-13 100 μg/L)[12];低浓度Dex组(IL-13 100 μg/L+Dex 0.01 mg/mL);中浓度Dex组(IL-13 100 μg /L+Dex 0.05 mg/mL);高浓度Dex组(IL-13 100 μg /L+Dex 0.25 mg/mL)[13]。将消化好的细胞转移到96孔板或6孔板,调整细胞密度为2×104/孔(96孔板)或2×105/孔(6孔板),含10%FBS的培养基先培养4~6 h,然后再用不含FBS的培养基饥饿过夜,根据分组的不同加用对应成分试剂分别培养48 h(培养条件为37 ℃、5%CO2)。
1.4 观察指标及测定方法
1.4.1 细胞免疫荧光法鉴定成纤维细胞 将细胞接种至带有玻璃片的六孔板中(2×105/孔)培养24 h,接着进行免疫荧光检测,步骤为:4%的多聚甲醛固定(15 min);0.1% Triton-100孵 育(10 min);10%的山羊血清封闭(30 min);一抗4 ℃过夜;二抗孵育30 min(室温);加入l:100稀释的DAPI;阴性对照以PBS代替一抗。荧光显微镜下观察并拍照。
1.4.2 CCK-8法测定成纤维细胞增殖水平 具体按说明书进行。接种成纤维细胞于96孔板(2×104/孔),每孔 100 μL,做 5个复孔。37 ℃、5%CO2培养24 h,然后根据不同的分组加入含有对应成分的培养基培养48 h,每孔加入CCK-8 10 μL培养2.5 h,通过酶标仪450 nm波长测定细胞的光密度(optical density,OD)值。采用扣除本底的策略来消除误差。
1.4.3 real-time PCR测定各组细胞中TGF-β1、Smad3及Smad4 mRNA的表达 将细胞提取总RNA,用完整性高、纯度好的RNA合成cDNA。荧光定量PCR反应体系为20 µL,其中2×SYBRRPremix 10 µL,样品 cDNA 2 µL,上、下游引物各0.8 µL,双蒸水 6 µL,ROX Reference Dye(50×)0.4 µL,重复4次。设置反应条件为:95 ℃ 30 s,然后进行40个扩增循环(95 ℃ 5 s、60 ℃ 30 s)。ABI Stepone系统收集及分析结果,引物特异性好时溶解曲线为单峰,用β-actin为内参,目的基因的相对表达通过2-△△Ct方法计算。
1.4.4 Western blot检 测 组 细 胞 中 TGF-β1及Smad4蛋白表达 提取细胞总蛋白,采用二喹啉甲酸(bicinchoninic acid,BCA)法检测蛋白浓度,以聚丙烯酰胺凝胶进行电泳。蛋白经PVDF膜转膜后进行封闭,1:5 000稀释目的蛋白一抗,在4 ℃孵育过夜,三羟甲基氨基甲烷缓冲盐溶液(tris buffered saline and tween 20,TBST)缓冲液清洗PVDF膜3次。1:5 000稀释二抗,室温孵育二抗2 h,TBST摇床上洗PVDF膜3次,每次10 min。电化学发光后通过显影、定影方法扫描胶片,β-actin为内参,用Quantity one系统分析目的条带的相对灰度值。
1.5 统计学处理
实验结果用均数±标准差(±s)表示,采用SPSS 16.0软件进行统计计算,统计分析用单因素方差分析(Bonferroni法),P<0.05为差异有统计学意义。
2 结 果
2.1 成纤维细胞的鉴定
镜下可见所培养细胞含有单个核细胞,从原代的不规则形逐渐变为梭形的成纤维样细胞。第3代成纤维样细胞波形蛋白细胞免疫荧光阳性,角蛋白细胞免疫荧光阴性,符合成纤维细胞的特点(图1)。
2.2 各组细胞增殖水平的测定
胆管成纤维细胞经IL-13处理后的OD值较空白对照组明显增加(均P<0.05);使用Dex干预后,IL-13处理的胆管成纤维细胞的增殖水平受到不同程度的抑制,中、高浓度的Dex对细胞增殖的抑制具有统计学意义(均P<0.05),且呈一定的浓度依耐性(表1)。
表1 各组细胞增殖情况(±s)Table 1 Proliferation in each group of cells(±s)

表1 各组细胞增殖情况(±s)Table 1 Proliferation in each group of cells(±s)
注:1)与空白对照组比较,P<0.05;2)与IL-13组比较,P<0.05Note: 1) P<0.05 vs. blank control group; 2) P<0.05 vs. IL-13 group
组别 OD值 相对IL-13组抑制率(%)空白对照组 0.332±0.005 13.99 IL-13组 0.386±0.0081) —低浓度Dex组 0.382±0.0041) 1.04中浓度Dex组 0.370±0.0071),2) 4.15高浓度Dex组 0.357±0.0071),2) 7.51
2.3 各组细胞TGF-β1、Smad3及Smad4基因mRNA表达水平
胆管成纤维细胞经IL-13处理后TGF-β1、Smad3及Smad4的mRNA表达较空白对照组明显上调(P<0.05);使用Dex干预后,IL-13处理的胆管成纤维细胞TGF-β1、Smad3及Smad4基因mRNA表达均明显的下调(均P<0.05),且呈现浓度依耐性(图2)。
2.4 各组细胞TGF-β1及Smad4蛋白表达水平的测定
IL-13处理的胆管成纤维细胞TGF-β1及Smad4蛋白表达较空白对照组明显上调(均P<0.05);使用Dex干预后,IL-13处理的胆管成纤维细胞的TGF-β1及Smad4蛋白表达受到不同程度的下调,其中低浓度的Dex对Smad4蛋白的下调有统计学意义(P<0.05),中、高浓度的Dex对TGF-β1及Smad4蛋白的下调均有统计学意义(均P<0.05)(图3)(表2)。
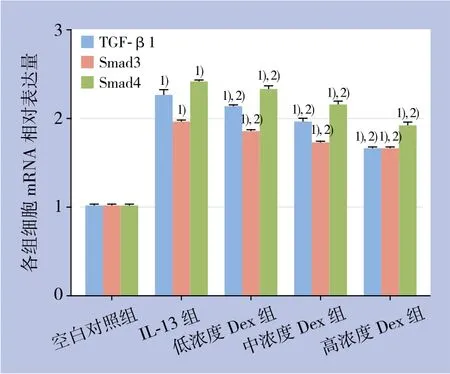
图2 各组细胞TGF-β1、Smad3及Smad4 mRNA表达水平Figure 2 Expression levels of TGF-β1, Smad3 and Smad4 mRNA in each group of cells

图3 各组细胞TGF-β1及Smad4蛋白表达检测Figure 3 Determination of TGF-β1 and Smad4 protein expressions in each group of cells
表2 各组细胞TGF-β1及Smad4蛋白表达水平(±s)Table 2 Expression levels of TGF-β1 and Smad4 protein in each group of cells(±s)
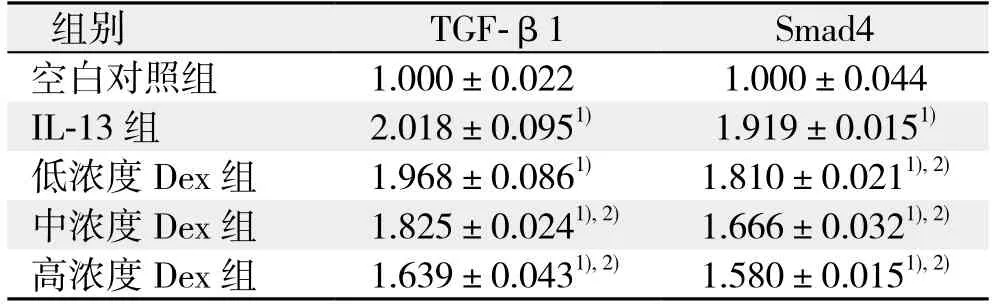
表2 各组细胞TGF-β1及Smad4蛋白表达水平(±s)Table 2 Expression levels of TGF-β1 and Smad4 protein in each group of cells(±s)
注:1)与空白对照组比较,P<0.05;2)与IL-13组比较,P<0.05Note: 1) P<0.05 vs. blank control group; 2) P<0.05 vs. IL-13 group
组别 TGF-β1 Smad4空白对照组 1.000±0.022 1.000±0.044 IL-13组 2.018±0.0951) 1.919±0.0151)低浓度Dex组 1.968±0.0861) 1.810±0.0211),2)中浓度Dex组 1.825±0.0241),2) 1.666±0.0321),2)高浓度Dex组 1.639±0.0431),2) 1.580±0.0151),2)
3 讨 论
不同程度的增生性疤痕的形成是组织损伤修复不可避免的结果,轻微的疤痕疙瘩组织可通过塑形得以纠正,但过多的疤痕疙瘩组织难以通过简单的塑形予以解决[14-15]。胆道增生性疤痕的形成意味着较高的BBS发病率。
成纤维细胞是组织损伤修复的主要效应细胞。组织损伤后,成纤维细胞迅速增殖并分化为肌成纤维细胞,分泌大量的细胞外基质(extracellular matrix,ECM)收缩及填补创面,同时又分泌TGF-β1等多种细胞因子调控组织修复过程[16-17]。正常情况下,当创面被完全表皮化,组织修复过程趋于停止,成纤维细胞及肌成纤维细胞就开始加速凋亡,细胞外基质的分泌就明显减少,同时开始加速降解;病理情况下,一些细胞因子表达异常,成纤维细胞等修复细胞过度增殖、细胞外基质过度积聚而导致增生性疤痕形成。成纤维细胞是胆管疤痕修复及BBS形成的主要效应细胞[3-4]。
TGF-β1是由成纤维细胞/肌成纤维细胞等细胞分泌的细胞因子,在调控细胞的分化、增殖、肿瘤形成及进展、疤痕形成等生理及病理过程中发挥了重要作用[18]。胆管疤痕组织及疤痕成纤维细胞中TGF-β1持续过表达,促使成纤维细胞过度增殖并转化为肌成纤维细胞,从而分泌大量细胞外基质,同时调控其它细胞因子的分泌,引起瘢痕形成,TGF-β1是目前已知的与胆管疤痕形成最密切的细胞因子[2-4,10]。TGF-β1主要依赖于它与TβRⅢ、TβRII及TβRI受体、Smad3/Smad2、Smad4、Smad7/Smad6等蛋白形成的TGF-β1/Smads通路发挥其生物学作用,该通路中,Smad3及Smad2是R-Smad(受体Smad),Smad4是通用Smad(co-Smad),Smad7及Smad6是抑制Smad(I-Smad),该通路可自行通过正负反馈进行调节[18]。TGF-β1/Smads通路的主要成分TβRII、TβRI、Smad4及TGF-β1等在胆管疤痕组织中均明显高表达,提示TGF-β1/Smads通路在BBS形成过程中可能发挥了重要作用[2]。目前治疗疤痕的重要措施之一就是调控疤痕组织及其成纤维细胞中TGF-β1/Smads通路的表达[19-20]。
IL-13有IL-13 Rαl和IL-13Rα2两种结构和功能截然不同的受体,IL-13Rαl通过与IL-4 Rα结合形成IL-13Rαl-IL-4Rα复合物参与IL-13的促纤维化作用[21]。IL-13在刺激成纤维细胞的迁移、增殖及活化并最终引起器官纤维化过程中发挥了重要作用[22]。IL-13通过调控气道成纤维细胞中TGF-β1及基质金属蛋白酶2(matrix metalloproteinase 2)的表达诱导I型胶原的产生[6]。并且IL-13在诱导气道成纤维细胞产生I型胶原的过程中可能受到了JAK/STAT6/PDGF/ERK1/2 MAPK通路的调控[7]。研究还发现,鼠疤痕组织中的IL-13的表达明显增高,提示IL-13在疤痕形成过程中可能发挥了重要作用[23]。IL-13在调控人的皮肤及疤痕成纤维细胞的胶原平衡过程中发挥了重要作用[24]。IL-13通过STAT6途径促进瘢痕成纤维细胞株胶原合成[25]。干扰素γ(IFN-γ)能抑制IL-13诱导的成纤维细胞I型胶原蛋白mRNA和蛋白表达水平的上调,从而抑制IL-13对成纤维细胞的纤维化作用[12]。本实验结果提示IL-13能上调胆管成纤维细胞TGF-β1/Smads通路的TGF-β1、Smad3及Smad4的mRNA及蛋白的表达(均P<0.05),从而刺激胆管成纤维细胞的增殖(P<0.05),提示IL-13可能通过活化胆管成纤维细胞TGF-β1/Smads通路在胆管疤痕修复及BBS形成过程中发挥了重要作用。
报道[8-9]显示Dex有抑制疤痕成纤维细胞增殖、抑制疤痕形成等作用,其机理可能与调控TGF-β1/Smads信号转导通路等因素有关。笔者[13]的前期研究结果也提示Dex能抑制BBS模型胆管成纤维细胞的增殖,调控BBS模型胆管成纤维细胞中TGF-β1/Smads信号转导通路的表达。本实验结果提示Dex(0.05~0.25 mg/mL)能下调IL-13诱导的胆管成纤维细胞TGF-β1/Smads通路的TGF-β1、Smad3及Smad4的mRNA及蛋白的表达(均P<0.05),从而抑制IL-13诱导的胆管成纤维细胞的增殖(P<0.05),提示Dex抑制BBS形成的机理之一可能与它抑制IL-13对胆管成纤维细胞TGF-β1/Smads通路的活化有关。
综上所述,IL-13可能通过活化胆管成纤维细胞TGF-β1/Smads通路在胆管疤痕修复及BBS形成过程中发挥了重要作用,Dex抑制BBS形成的机理之一可能与它抑制IL-13对胆管成纤维细胞TGF-β1/Smads通路的活化有关。
参考文献
[1] Shimada H, Endo I, Shimada K, et al. The current diagnosis and treatment of benign biliary stricture[J]. Surg Today, 2012,42(12):1143–1153. doi: 10.1007/s00595–012–0333–3.
[2] Geng ZM, Zheng JB, Zhang XX, et al. Role of transforming growth factor-beta signaling pathway in pathogenesis of benign biliary stricture[J]. World J Gastroenterol, 2008, 14(31):4949–4954.
[3] Geng ZM, Yao YM, Liu QG, et al. Mechanism of benign biliary stricture: a morphological and immunohistochemical study[J].World J Gastroenterol, 2005, 11(2):293–295.
[4] Xu J, Geng ZM, Ma QY. Microstructural and ultrastructural changes in the healing process of bile duct trauma[J]. Hepatobiliary Pancreat Dis Int, 2003, 2(2):295–299.
[5] Huang C, Ogawa R. Pharmacological treatment for keloids[J].Expert Opin Pharmacother, 2013, 14(15):2087–2100. doi:10.1517/14656566.2013.826651.
[6] Firszt R, Francisco D, Church TD, et al. Interleukin-13 induces collagen type-1 expression through matrix metalloproteinase-2 and transforming growth factor-β1 in airway fibroblasts in asthma[J].Eur Respir J, 2014, 43(2):464–473. doi: 10.1183/09031936.00068712.
[7] Lu J, Zhu Y, Feng W, et al. Platelet-derived growth factor mediates interleukin-13-induced collagen I production in mouse airway fibroblasts[J]. J Biosci, 2014, 39(4):693–700.
[8] Syed F, Singh S, Bayat A. Superior effect of combination vs. single steroid therapy in keloid disease: a comparative in vitro analysis of glucocorticoids[J]. Wound Repair Regen, 2013, 21(1):88–102. doi:10.1111/j.1524–475X.2012.00862.x.
[9] Wrzeszcz A, Dittrich B, Haamann D, et al. Dexamethasone released from cochlear implant coatings combined with a protein repellent hydrogel layer inhibits fibroblast proliferation[J]. J Biomed Mater Res A, 2014, 102(2):442–454. doi: 10.1002/jbm.a.34719.
[10] Li KY, Shi CX, Huang JZ, et al. Tetramethylpyrazine Effects on the Expression of Scar-Related Genes in Rabbit Benign Biliary Stricture Fibroblasts[J]. J Coll Physicians Surg Pak, 2016, 26(10):813–817.
[11] 李克跃, 石承先, 汤可立, 等. TNF-α对兔胆管成纤维细胞P311/TGF-β1/α-SMA通路的影响及川芎嗪的干预作用[J]. 中国普通外科杂志, 2017, 26(2):172–178. doi: 10.3978/j.issn.1005–6947.2017.02.007.Li KY, Shi CX, Tang KL, et al. Influence of TNF-α on activity of P311/TGF-β1/α-SMA signaling pathway in rabbit bile duct fibroblasts and the interventional effect of tetramethylpyrazine[J].Chinese Journal of General Surgery, 2017, 26(2):172–178. doi:10.3978/j.issn.1005–6947.2017.02.007.
[12] 熊丽霞, 李文林, 蔡震宇, 等. 干扰素γ抑制IL-13对成纤维细胞的纤维化作用[J]. 中国病理生理杂志, 2013, 29(3):504–508.doi:10.3969/j.issn.1000–4718.2013.03.021.Xiong LX, Li WL, Cai ZY, et al. Suppressive effect of interferon γ on IL-13-induced fibrosis in fibroblasts[J]. Chinese Journal of Pathophysiology, 2013, 29(3):504–508. doi:10.3969/j.issn.1000–4718.2013.03.021.
[13] Li K-, Shi C-, Huang J-, et al. Dexamethasone Effect on the Expression of Transforming Growth Factor-β1/Smads Signaling Pathway in Benign Biliary Stricture Fibroblasts in Rodent[J]. J Coll Physicians Surg Pak, 2017, 27(3):131–134. doi: 2571.
[14] Gauglitz GG, Korting HC, Pavicic T, et al. Hypertrophic scarring and keloids: pathomechanisms and current and emerging treatment strategies[J]. Mol Med, 2011, 17(1/2):113–125. doi: 10.2119/molmed.2009.00153.
[15] Berman B, Maderal A, Raphael B. Keloids and Hypertrophic Scars:Pathophysiology, Classification, and Treatment[J]. Dermatol Surg,2017, 43(Suppl 1):S3–18. doi: 10.1097/DSS.0000000000000819.
[16] Hu B, Phan SH. Myofibroblasts[J]. Curr Opin Rheumatol, 2013,25(1):71–77. doi: 10.1097/BOR.0b013e32835b1352.
[17] Hinz B. Myofibroblasts[J]. Exp Eye Res, 2016, 142:56–70. doi:10.1016/j.exer.2015.07.009.
[18] No authors listed. Correction: Role of Transforming Growth Factor(beta) in Human Disease[J]. N Engl J Med, 2000, 343(3):228.
[19] Zhou P, Shi L, Li Q, et al. Overexpression of RACK1 inhibits collagen synthesis in keloid fibroblasts via inhibition of transforming growth factor-beta1/Smad signaling pathway[J]. Int J Clin Exp Med, 2015, 8(9):15262–15268.
[20] Zhang YF, Zhou SZ, Cheng XY, et al. Baicalein attenuates hypertrophic scar formation via inhibition of the transforming growth factor-beta/Smad2/3 signalling pathway[J]. Br J Dermatol,2016, 174(1):120–130. doi: 10.1111/bjd.14108.
[21] Zheng T, Zhu Z, Liu W, et al. Cytokine regulation of IL-13Ralpha2 and IL-13Ralpha1 in vivo and in vitro[J]. J Allergy Clin Immunol,2003, 111(4):720–728.
[22] Wynn TA. Integrating mechanisms of pulmonary fibrosis[J]. J Exp Med, 2011, 208(7):1339–1350. doi: 10.1084/jem.20110551.
[23] Li SH, Yang HL, Xiao H, et al. Inflammation and cutaneous nervous system involvement in hypertrophic scarring[J]. Neural Regen Res,2015, 10(10):1678–1682. doi: 10.4103/1673–5374.167769.
[24] Oriente A, Fedarko NS, Pacocha SE, et al. Interleukin-13 modulates collagen homeostasis in human skin and keloid fibroblasts[J]. J Pharmacol Exp Ther, 2000, 292(3):988–994.
[25] 熊丽霞, 李文林, 蔡震宇, 等. 白细胞介素13通过STAT6途径促进瘢痕成纤维细胞株胶原合成[J]. 中国细胞生物学学报, 2010,32(6):868–872.Xiong LX, Li WL, Cai ZY, et al. Upregulation of Collagen Generation by IL-13 in Human Keloid Fibroblasts Strain via the STAT6 Signaling Pathways[J]. Chinese Journal of Cell Biology,2010, 32(6):868–872.
