海洋异养硝化-好氧反硝化菌y6同步脱氮除碳特性
2017-04-08王骁静于德爽都松东吴国栋
王骁静,于德爽,李 津,都松东,周 同,吴国栋
(青岛大学环境科学与工程学院,山东 青岛 266071)
海洋异养硝化-好氧反硝化菌y6同步脱氮除碳特性
王骁静,于德爽,李 津*,都松东,周 同,吴国栋
(青岛大学环境科学与工程学院,山东 青岛 266071)
从胶州湾海底沉积物中分离筛选出一株异养硝化-好氧反硝化菌株y6,通过菌株y6的形态以及生理生化特性和16S rRNA基因序列的分析,鉴定该菌株属于克雷伯氏菌属(Klebsiella sp.).在不同的环境条件下,测定菌株y6的生长情况和脱氮能力,研究其同步脱氮除碳特性.实验结果表明,菌株y6的最佳碳源为柠檬酸三钠,最适宜pH值为7.0,最适合的C/N为17.菌株y6在以NH4Cl、KNO3和NaNO2为唯一氮源的反应系统中均有较好的脱氮效果,去除率分别为99.67%、100%、99.20%.菌株y6在脱氮的同时能高效地去除有机物,COD的去除率分别为82.17%、95.75%和97.83%.菌株y6在硝化过程中没有亚硝态氮和硝态氮的积累.在按不同比例混合氮源的反应系统内,首先进行的是硝态氮的好氧反硝化,随后进行的是氨氮、亚硝态氮和COD的去除.在有亚硝态氮存在时氨氮的去除率略低,亚硝态氮会影响y6的异养硝化过程,异养硝化对好氧反硝化过程没有影响.
异养硝化-好氧反硝化;海洋菌株;克雷伯氏菌y6;同步脱氮除碳
随着社会的发展,水污染的防治越来越受到人们的重视.如何解决氮元素过剩所引起的水体富营养化是治理水污染的重要方面.随着科技的进步,生物脱氮以其高效、经济、简便的特性,已经成为去除水中氮元素的主要手段.但是,传统的生物脱氮分为好氧硝化阶段和缺氧反硝化阶段,要求在两个反应器中进行,建设成本高[1],而且自养硝化菌的成长速度缓慢.近年来, 通过发现具有异养硝化—好氧反硝化特性的菌株(Paracoccus pantotrophusATCC 35512),而提出异养硝化—好氧反硝化菌的概念[2].在之后的时间里,越来越多的类似这种性质的菌株被发现.HN-AD菌具有生长速度快、同步脱氮除碳、在一个反应器中完成硝化和反硝化两个过程、占地面积少等特点[3],引起国内外的重视,成为研究的热点.
目前,工业排放的含氮废水盐度都较高,高盐度使得微生物中脱氢酶的活性下降[4],影响微生物的生长代谢,渗透压升高会导致细菌细胞质壁分离甚至破裂死亡.现阶段国内外主要从地表水、土壤和生活工业废水中分离出HN-AD菌,研究较多的是在低盐废水中HN-AD菌的生长以及同步脱氮除碳特性,对高盐环境中的HN-AD菌鲜有报道[5].因此,本实验研究在海洋条件下微生物的异养硝化-好氧反硝化脱氮特性更具有现实意义.
本研究从胶州湾海底沉积物中分离筛选了一株海洋HN-AD菌y6.对其形态、生理生化以及16S rRNA序列进行了研究分析.考察了在海水环境中菌株y6生长的最佳碳源、pH值以及C/N,并分析了在单一和混合氮源环境下的同步脱氮除碳效果,以期丰富高盐条件下的生物脱氮除碳机制,为高盐废水生物处理的实际应用提供了理论依据和支持.
1 材料与方法
1.1 菌株来源、培养基和细菌鉴定
菌株来源、培养基的配制以及细菌鉴定实验方法参照本实验室近年来的研究成果[6],在此基础上进行了改进.HN-AD培养基:KH2PO40.2g,K2SO40.1g,NH4Cl 0.19g,KNO20.36g,柠檬酸三钠 6.944g,微量元素溶液Ⅰ、Ⅱ各1.25mL,海水1000mL,pH 7.0.在相应实验中,对HN-AD培养基中的碳源和氮源做出相应的调整.
1.2 影响菌株y6的生长因素
本实验选择的生长因素为碳源、初始pH值和C/N 3项,碳源分别为乙酸钠、丁二酸钠、柠檬酸三钠、蔗糖和葡萄糖,初始pH值设为5.0、5.5、6、6.5、7、7.5、8、8.5、9,C/N(培养基中COD与总氮的比)为1、5、9、13、17、21、25.设置NH4Cl和KNO3的初始量为50mg/L,总氮为100mg/L.以HN-AD培养基为基础,设置单一变量,碳源为柠檬酸三钠,初始pH值为7,C/N为17.将处于对数生长期的活化菌液按照8%的接种量(吸取8mL)接种到经过高温灭菌的液体培养基中.置于气浴恒温振荡器(转速140r/min,温度30℃)中,培养20h后检测培养基中菌体的生长量(OD600),以及NH4+_N、NO2-_N、NO3-_N和COD的含量.考察不同生长因素对菌株y6脱氮效果的影响,选出适宜菌株y6生长的最佳碳源、初始pH和C/N.
1.3 菌株y6的脱氮特性研究
依照HN-AD培养基,选碳源为柠檬酸三钠,初始pH值为7.0,C/N为17,配菌株y6的脱氮性能培养基.分别设置单一氮源系统和混合氮源系统,考察菌株y6的HN-AD能力.
菌株y6的脱氮性能实验设计如表1:
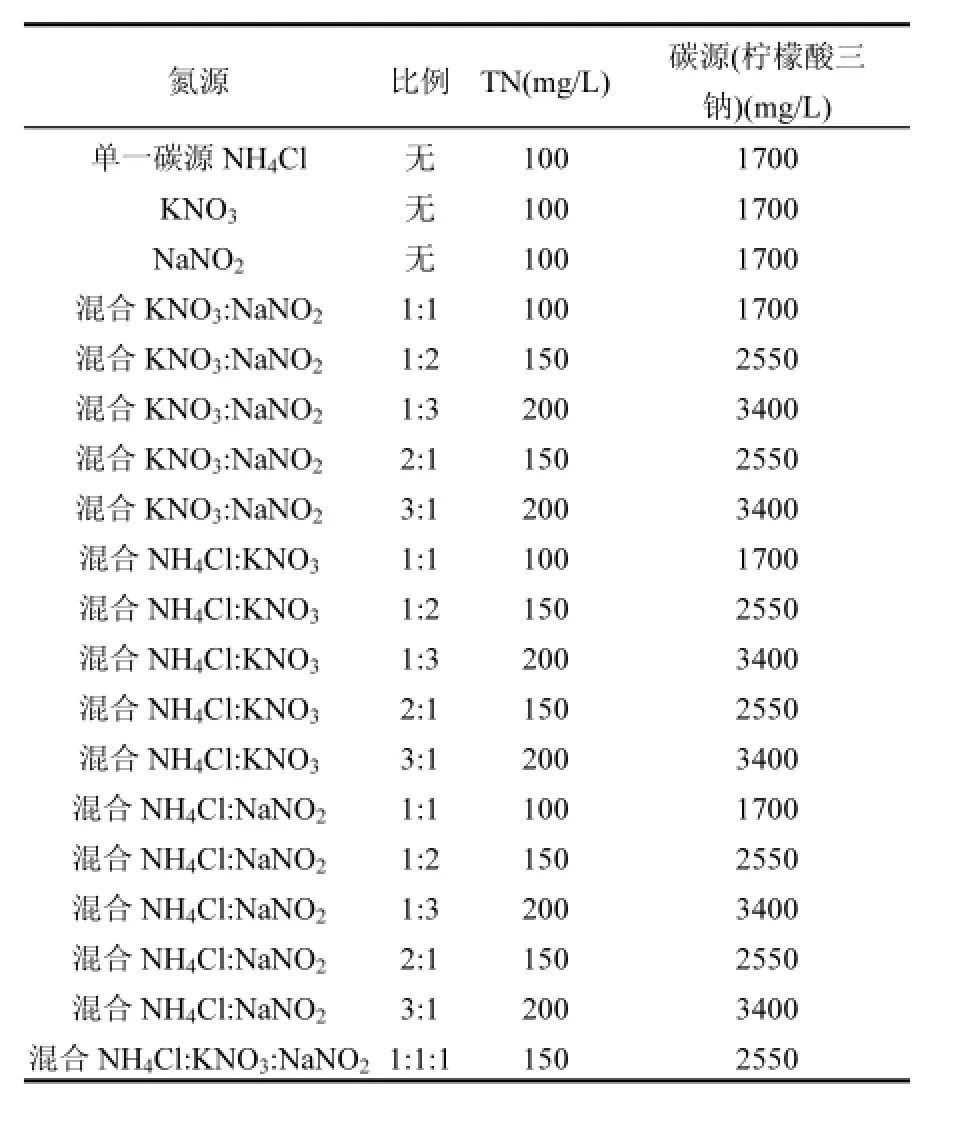
表1 菌株y6的脱氮性能实验设计Table 1 The experimental design on denitrification performance of strain y6
1.4 分析方法
NH4+_N:钠氏试剂分光光度法;NO2-_N: N-(1-萘基)乙二胺分光光度法;NO3-_N:麝香草酚分光光度法;菌体生长量(OD600):测定菌液在600nm波长时的吸光度值;pH值和ORP使用WTW pH/Oxi340i/SET测定;COD:碱性高锰酸钾法.3次重复实验取平均值.
2 结果与讨论
2.1 菌株形态特征与生理生化鉴定
菌株y6在分离和生长固体培养基上培养15h后,形成的菌落呈乳白色,不透明,边缘整齐,表面隆起,易挑取.菌株y6大小为1.1µm×0.7µm,为短杆菌,没有鞭毛和芽孢.菌株y6的生理生化鉴定结果可知,菌株y6氧化酶反应阴性,水解酶反应、甲基红实验以及吲哚实验均呈现阳性.生理生化指标与摩根氏菌有99.93%的相似性.对于鸟氨酸、葡萄糖、丙二盐、色氨酸等大分子有机底物,该菌株y6均能进行水解利用,说明菌株y6有相应的酶系统.
2.2 y6菌株16S rRNA测序及系统发育分析
对菌株y6测序获得长度为1416bp的部分16S rRNA序列.将所得的菌株序列提交至GenBank中通过Blast检索,应用MEGA6软件,以Neighbor-joining 法绘制系统发育树,从菌株y5的系统发育树上可以看出,菌株y6与多株Klebsiellasp. 16S rRNA 的相似性达99%,结合菌株的形态特征和生理生化特性,可初步确定菌株y6属于克雷伯氏菌属(Klebsiella sp)[7].
2.3 菌株脱氮过程中生长因素的研究
2.3.1 碳源对菌株脱氮特性的影响 如表2,当乙酸钠、丁二酸钠、柠檬酸三钠、蔗糖和葡萄糖分别作为HN-AD培养基的唯一碳源时,菌株y6均正常生长并且有一定的脱氮能力.菌株y6在以乙酸钠为碳源时的生长量(OD600)最大,丁二酸钠和柠檬酸三钠次之,蔗糖和葡萄糖的OD600最低.碳源为丁二酸钠时,NH4+_N去除率达到最高,为93.85%,其次是柠檬酸三钠为碳源时的NH4+_N去除率为92.62%.目前研究[8]发现,碳源的结构越简单,分子质量越小,越容易促进菌株生长,保证反硝化的顺利进行.蔗糖和葡萄糖大分子为碳源时,NH4+_N去除率只有30%多.以丁二酸钠和柠檬酸三钠为碳源时,NO3-_N的去除率很高,均达到了90%以上,5种不同碳源的脱氮测试液中COD去除率都达到了80%以上.蔗糖和葡萄糖的NO2-_N积累量很高.丁二酸钠与柠檬酸三钠相比,NO2-_N的积累量略高,所以菌株y6的最佳碳源选择柠檬酸三钠.
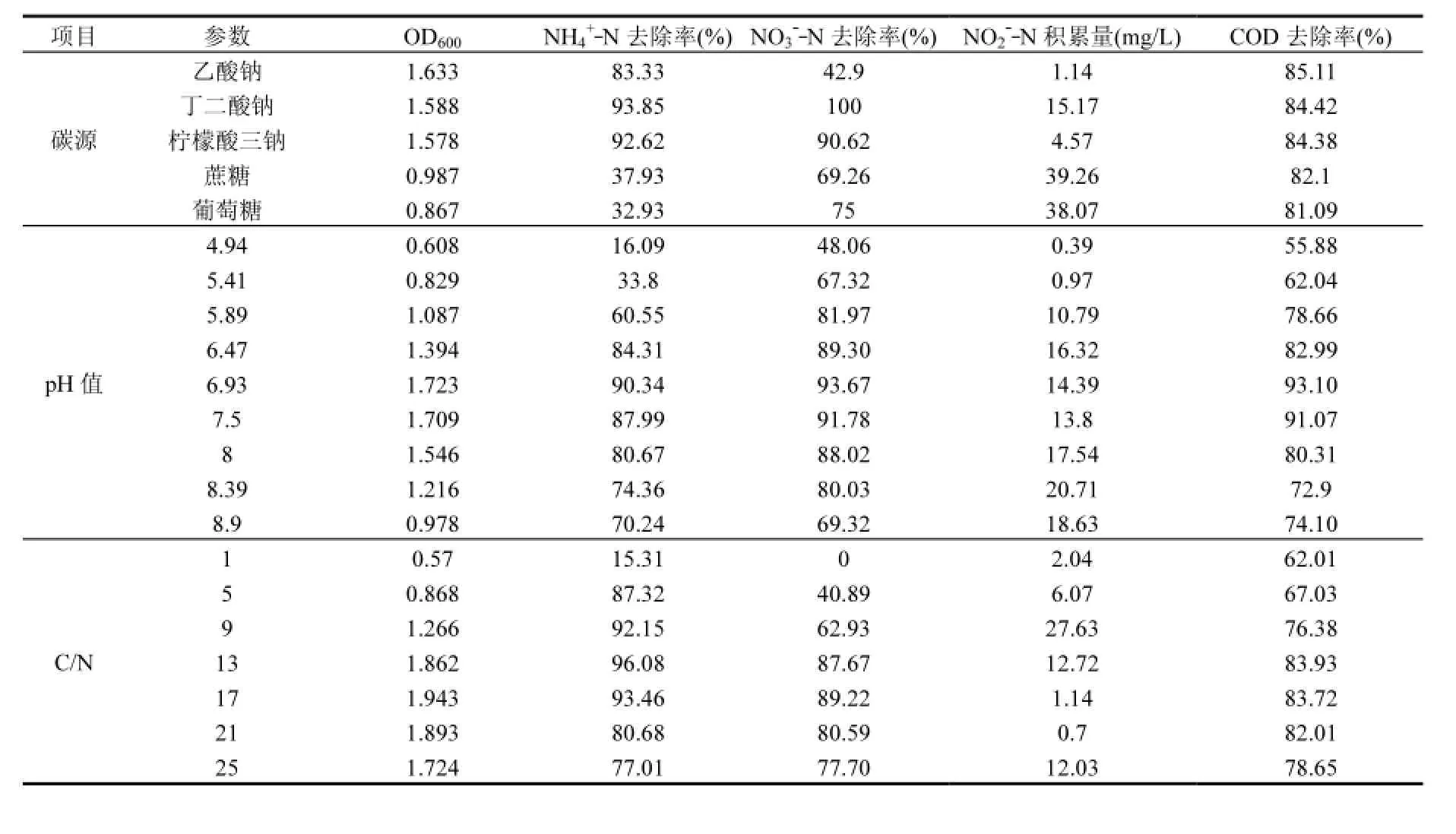
表2 碳源、初始pH和C/N对菌株y6的生长和脱氮反应的影响Table 2 Effects of carbon source, pH and C/N ratio on the growth and nitrogen removal of strain y6
2.3.2 初始pH值对菌株脱氮特性的影响 表2初始pH值在6.5~8之间时,菌株y6的OD600值都达到1.3以上,对NH4+_N和COD的降解率均在80%以上.初始pH值在7~7.5之间时,菌株y6的OD600高,最高达到1.723,NO3-_N和COD的去除率均超过了90%,NO2-_N的积累量较少.HN-AD菌适宜的pH值范围大多是中性或是偏碱性[9].pH值为7时,NH4+_N、NO3-_N和COD的去除率达到了最高,分别为90.34%、93.67%和93.10%.所以菌株y6较为合适的初始pH值在6.5~8范围之间,最适初始pH值为7.0.
2.3.3 C/N对菌株脱氮特性的影响 表2可知, C/N在1~17时,随着C/N的增大,菌株y6的生长量、脱氮效率和COD去除率一直在增大.当C/N超过21,菌株生长量以及脱氮效率均有所下降.C/N在9~17之间时,NH4+_N的降解率达到90%以上;C/N在13~21之间时,NO3-_N和COD的降解率均超过了80%.C/N值为17时,菌株y6的NH4+_N和NO3-_N的降解率均达到最大,分别为93.46%和89.22%,COD的降解率也很高,达到83.72%,NO2-_N几乎没有积累,只有1.14mg/L.所以菌株y6的最适C/N值为17.
2.4 单一氮源中菌株y6的脱氮特性
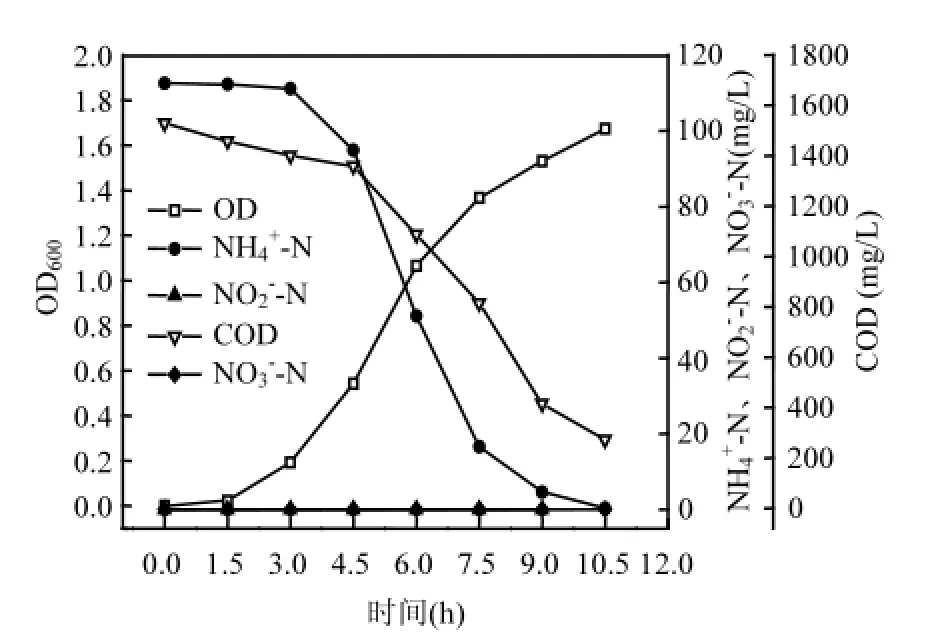
图1 菌株y6的异养硝化过程Fig.1 Process of heterotrophic nitrification of strain y6
2.4.1 菌株y6的异养硝化性能 图1为以NH4Cl为唯一氮源的异养硝化反应过程,从初始状态每隔1.5h取样检测.菌株y6生长的适应期为1.5h,从1.5h后菌株y6开始增长,10.5h后OD600达到最大吸光度1.673.NH4+_N经过3h只有少量的降解,从3h后,NH4+_N迅速降解,到10.5h,NH4+_N浓度降为0.37mg/L.去除率为99.67%.COD在0~4.5h去除的较为缓慢,此段的平均去除速率为38.10mg/(L·h).从4.5h后,COD迅速去除,到10.5h,降解到272.71mg/L,平均降解速率为180.95mg/(L·h).COD的去除率为82.17%.NH4+_N和COD的降解主要发生在菌株y6的对数生长期.菌株y6的异养硝化过程中几乎没有NO3-_N和NO2-_N的积累.
2.4.2 菌株 y6的好氧反硝化性能 图2为以KNO3为唯一氮源的好氧反硝化过程,从初始状态每隔2.5h取样检测.0~5h是y6生长的适应期,5h之后,y6进入对数增长期,15h达到最大OD600值1.37.NO3-_N从初始就被降解,由106.14mg/L降解到73.54mg/L,7.5h后,NO3-_N迅速降解为0,NO3-_N的去除率为100%.7.5hNO3-_N的迅速去除,使得NO2-_N大量积累,浓度达到55.15mg/L,之后又被迅速降解.COD在0~5h去除速率较低,平均去除速率为19.86mg/(L·h);在5~10h之间,进入了y6的对数增长期,COD的降解速率明显提高,平均去除速率为65.38mg/(L·h);10h之后,COD迅速降解,与NO2-_N的降解一致,到15h降解到70.11mg/L,平均去除速率为230.54mg/(L·h),COD的去除率为95.75%.NO3-_N和COD的降解都是在菌株y6的对数增长期,并且COD是随着NO2-_N的降解而迅速去除.
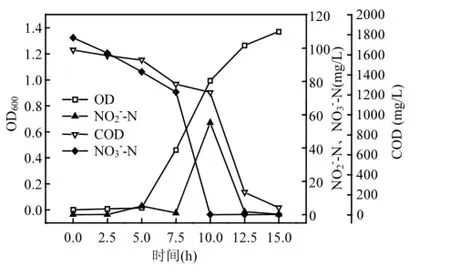
图2 菌株y6的硝酸盐好氧反硝化过程Fig.2 Process of nitrite denitrification of strain y6

图3 菌株y6的亚硝酸盐好氧反硝化过程Fig.3 Process of nitrate denitrification of strain y6
图3为只投加了NaNO2的好氧反硝化过程,0h~7.5hOD600只有微量增长.7.5h后,y6进入了对数增长期,到17.5h后OD600达到了1.339. NO2-_N在0~7.5h之间降解的十分缓慢,7.5h后, NO2-_N迅速去除,到17.5h浓度降为0.77mg/L, NO-_N的去除率为99.20%.COD的降解与NO-_N22的降解统一,从5h后开始去除,到17.5h后浓度为35.7mg/L,平均去除速率为128mg/(L·h), COD的去除率为97.83%.反应过程中几乎没有NO3-_N的积累,只有前期少量出现,而后又被迅速降解.
2.5 混合氮源中菌株y6的脱氮特性
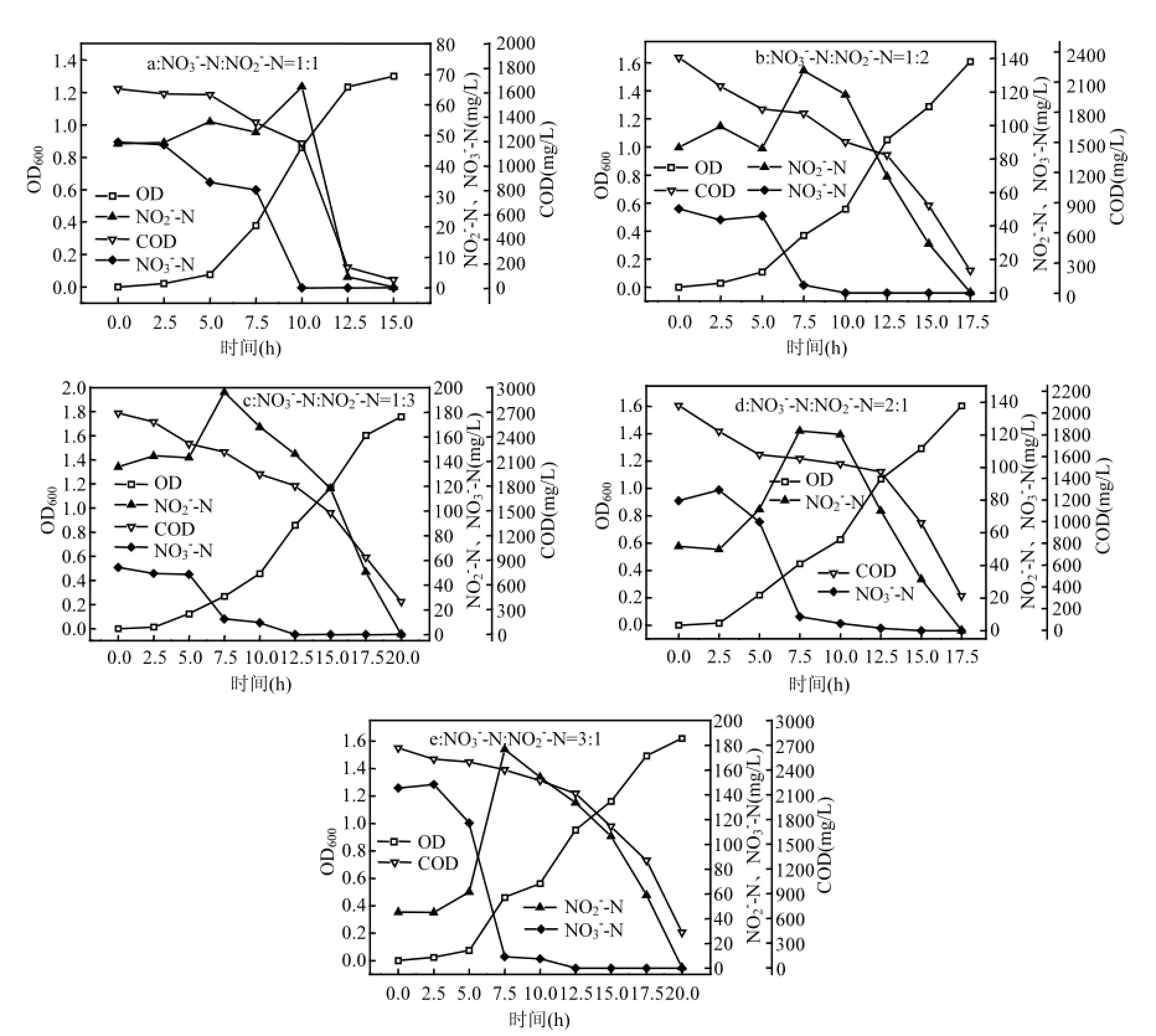
图4 混合氮源系统内菌株y6的好氧反硝化过程Fig.4 Process of mixed nitrogen sources aerobic denitrification of strain y6
2.5.1 菌株y6的好氧反硝化性能 图4所示,在NO3-N和NO2-N以不同比例进行好氧反硝化反应的混合系统中,0~5h为菌株y6的生长适应期,5h之后进入对数生长期,迅速生长.在NO3-N:NO2-N分别为1:1、1:2、1:3、2:1、3:1的系统中,反硝化分别在15h、17.5h、20h、17.5h和20h反应完全,OD600分别为1.301、1.609、1.757、1.605、1.620.无论混合系统的比例如何,首先进行的是NO3-_N的反硝化反应,NO3-_N在y6的生长适应期就开始降解,在对数期迅速降解彻底,并且积累相应浓度的NO2-_N,而后NO2-_N被迅速降解.在y6生长适应期,COD和NO3-_N有少量去除,在对数生长期时,由于NO2-_N短时间的迅速积累,导致COD去除速率降低,而后随着NO2-_N的去除,COD的去除速率迅速提高,很快被降解完全.NO3-N:NO2-N分别为1:1、1:2、1:3、2:1、3:1的系统中,NO3-_N的去除率均为100%,这与添加KNO3的单一氮源系统相比去除率没有变化;NO2-_N的去除率分别为:99.64%、99.71%、99.76%、99.67%、99.12%,不同比例之间相差不大,与添加NaNO2的单一氮源系统比较, NO3-N和NO2-_N混合后NO2-_N的去除率略高;COD的去除率分别为95.71%、90.37%、85.34%、84.79%、83.93%,混合反硝化系统的COD去除率均低于只投加KNO3或NaNO2单一氮源中COD的去除率.

图5 NH4+-N和NO3-N混合氮源系统下菌株y6的脱氮过程Fig.5 Process of nitrogen removal in the NH4+-N and NO3-N systemof strain y6
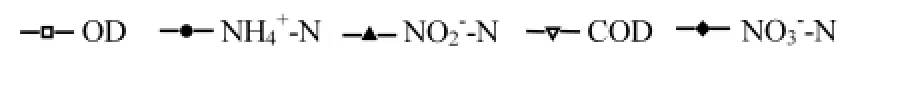
2.5.2 菌株y6的异养硝化-好氧反硝化性能 图5所示,在NH4+-N和NO3-N以不同比例混合的HN-AD系统中,细菌的生长适应期短, NH4+-N:NO3-N为1:2和1:3时,y6的适应期为5h,NH4+-N:NO3-N为1:1、2:1和3:1时,适应期为2.5h,之后菌株y6迅速繁殖,进入对数增长期. NH4+-N:NO3-N分别为1:1、1:2、1:3、2:1、3:1的系统中,最终反应完全的时间分别为17.5、20、22.5、15和17.5h,OD600分别为1.599、1.753、1.852、1.741和2.032.说明NH4+_N的添加对菌株的生长有促进作用.NH4+_N和NO3-N的混合系统中,首先是NO3-_N在y6生长的适应期就开始反应,到7.5hNO3-_N几乎完全降解,NO3-_N的快速降解使得NO2-_N在7.5h达到最高,而后被降解完全,NH4+_N和COD在适应期降解缓慢,在对数生长后期去除,与NO2-_N的降解几乎同步.NH4+-N:NO3-N分别为1:1、1:2、1:3、2:1、3:1的系统中,NO3-_N的去除率均为100%; NH4+_N的去除率分别为:100%、100%、100%、91.97%、91.93%,说明NH4+_N和NO3-_N的混合具有互相促进的作用.NH4+_N的浓度增加具有抑制作用,使得NH4+-N:NO3-N为2:1和3:1的系统中,NH4+_N的去除率降低.COD的去除率分别为90.22%、87.94%、79.18%、74.18%和73.49%,混合反硝化系统的COD去除率均低于只投加KNO3或NaNO2单一氮源中COD的去除率.
图6所示,NH4+-N和NO2-N以不同比例混合的HN-AD系统中,0~2.5h为细菌的生长适应期,之后菌株y6进入对数增长期.NH4+-N: NO2-N分别为1:1、1:2、1:3、2:1、3:1的系统中,最终反应完全的时间分别为15、17.5、20、17.5和17.5h,比NH4+_N和NO3-_N的混合系统的反应时间相对较短,OD600分别为1.634、1.426、1.598、1.572和1.768.NH4+_N、NO2-_N和COD的降解几乎同步,在y6的适应期去除缓慢,进入对数生长期,三者迅速反应,去除速率加快.除了1:3混合系统NO2-_N的去除率为99.47%,其余系统中NO2-_N的去除率均为100%.NH4+_N的去除率分别为:88.00%、84.17%、84.63%、97.93%、94.21%,低于只添加NH4Cl的单一氮源系统的去除率.COD的去除率分别为83.89%、85.77%、88.62%、82.31%和82.07%,混合反硝化系统的COD去除率均低于只投加KNO3或NaNO2单一氮源中COD的去除率.


图6 NH4+-N和NO2-N混合氮源系统下菌株y6的脱氮过程Fig.6 Process of nitrogen removel in the NH4+-N and NO2-N systemof strain y6

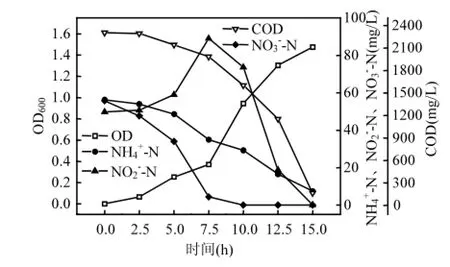
图7 、和混合氮源系统下菌株y6的脱氮过程Fig.7 Process of nitrogen removal in the、andsystemof strain y6
3 讨论
目前已经发现的同时具有HN-AD能力的细菌有:盐单胞菌(Halomonas)、假单胞菌(Pseudomons)、副球菌属(Paracoccus)[10]、硫球形菌(Thiosphaera pantotropha)、芽孢杆菌(Bacillussp.)[11]、产碱杆菌属(Alcaligenes)[12]等.本实验所研究的菌株y6是从胶州湾深海底泥中分离筛选出来,具有HN-AD能力.C/N在很大程度上影响菌株的物质及能量代谢.充足的碳源会提供足够的电子流为菌株生长供应能量.目前研究[13-14]表明,随着C/N增大,反应体系中脱氮效率会增加,达到一定程度后,脱氮效率随着C/N值的增加而有所降低.戴娴等[15]发现C/N过高过低均会制约同步硝化反硝化过程.李鹏章[16]等的研究表明C/N过低时会不断地产生N2O.
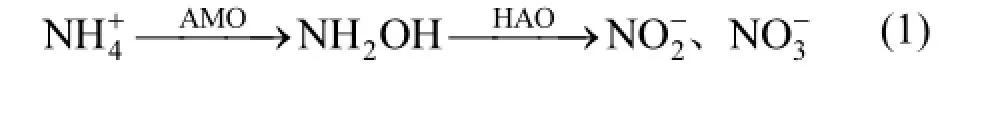
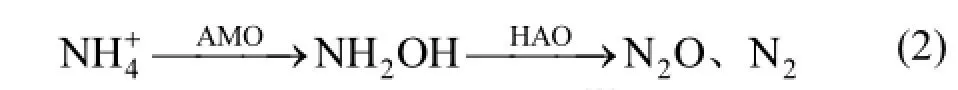
菌株的好氧反硝化途径[3]如式(3)所示.本实验KNO3为唯一氮源的反应过程中,积累了大量的NO2-_N,而后被迅速降解.NaNO2为唯一氮源的反应过程中,几乎无NO3-_N积累.在分别以NH4Cl、KNO3和NaNO2为氮源的系统中,NH4Cl的异养硝化过程菌株y6的生长量高于好氧反硝化过程中的生长量,y6的异养硝化中COD的去除率低于好氧反硝化过程中的.反硝化过程需要消耗碳源,碳源是电子供体,硝态氮或NO2-_N和氧气是电子受体,在好氧环境中完成反应.

NH4+_N、NO3-_N和NO2-_N的混合系统中,首先NO3在硝酸盐还原酶(NAR)作用下迅速转化为NO,NH+的降解略有滞后,随后NO和242NH4+转化为N2溢出.本实验NH4+_N和NO3-_N或是NH4+_N与NO2-_N的混合系统中,NH4+_N的浓度对y6的反硝化过程没有影响.类似于Stewart[21]的观点,NH4+_N不会影响NAR的活性.张培玉等[22]研究认为异养硝化过程不会影响到好氧反硝化过程.NO3-_N的浓度对y6的异养硝化也没有影响,整体去除率都很高;NH4+_N和NO2-_N混合反应时,NH4+_N去除率低于只添加NH4Cl的去除率,说明NO2-_N的浓度对y6的异养硝化过程中的NH4+_N去除率有影响.与Kim等[4]的研究结果类似,亚硝酸盐对异养硝化有一定的抑制作用.NH4+_N与NO3-_N不同比例混合反应时,首先NO3-_N被迅速还原,转化为NO2-_N,NO2-_N在短时间内的大量积累导致在该段时间内菌株y6生长缓慢,并且该混合系统反硝化需要两步完成,NO3-_N转化为NO2-_N再转化为气态氮,使得COD的去除有所滞后,此过程中发现NH4+_N的浓度对COD的降解速率有一定的影响,导致COD的总体去除率不如NH4+_N与NO2-_N的混合系统,并且按不同比例混合的体系所需要的反应时间比NH4+_N与NO2-_N相应比例混合的反应时间要长一些.NO3-_N和NO2-_N按不同比例的混合反应系统中, 由于NO3-_N的氧化还原电位比较高,将其作为电子受体时基质会产生更多的能量[23],所以NO3-_N先进行反硝化,随后进行NO2-_N的反硝化,此系统中NO3-_N、NO2-_N和COD的去除率都很高,其中COD的去除率三个混合系统中最高的.在NH4+_N、NO3-_N和NO2-_N按1:1:1的混合系统中,由于NO2-_N对异养硝化的抑制作用,菌株y6的NH4+_N去除率略低,COD的去除率比NO3-_N和NO2-_N的混合系统低,但是比NH4+_N和NO3-_N或NO2-_N的混合系统均要高.
4 结论
4.1 本实验所用HN-AD菌y6来自于胶州湾海底沉积物,经分离筛选、形态观察、生理生化实验和16S rRNA序列分析,鉴定为克雷伯氏菌属(Klebsiella.sp).
4.2 在分别以乙酸钠、丁二酸钠、柠檬酸三钠、葡萄糖和蔗糖为碳源的反应过程中,菌株y6均能正常生长,其中y6的最佳碳源选柠檬酸三钠,最适pH值为7.0,最佳C/N选为17.
4.3 菌株y6均能去除NH4+_N、NO3-_N和NO2-_N,在脱氮过程中能同步去除COD,主要在菌株的对数生长期进行.
4.4 在混合系统中,首先进行NO3-_N的好氧反硝化,转化为NO-_N,随后是NH+_N和NO-_N242的异养硝化-好氧反硝化同步脱氮除碳过程.NH4+_N的异养硝化不会影响好氧反硝化,NO3-_N对NH4+_N的异养硝化几乎没有影响,NO2-_N在一定程度上抑制了NH4+_N的异养硝化过程.
[1] 马 放,王弘宇,周丹丹.好氧反硝化生物脱氮机理分析及研究进展 [J]. 工业用水与废水, 2005,36(2):11-14.
[2] Robertson L A, Kuenen J G. Thiosphaera pantotropha gen.nov.sp.nov. a facultatively anaerobic,fcultatively autotrophic sulphur hacterium[J]. Microbiology, 1983,129(9):2847-2855.
[3] 苏婉昀,高俊发,赵红梅.异养硝化-好氧反硝化菌的研究进展[J]. 工业水处理, 2013,33(12):01-05.
[4] K imY J, Yoshizawa M, Takenaka S, et al.Isolation and culture conditions of a klebsiella pneumoniae strain that can utilizeammoniumand nitrate ions simultaneously with controlled iron and molybdate ion concentrations [J]. Bioscience Biotechnology & Biochemistry, 2002,66(5):996-1001.
[5] 孙庆花,于德爽,张培玉.1株海洋异养硝化-好氧反硝化菌的分离鉴定及其脱氮特性 [J]. 环境科学, 2016,37(2):647-654.
[6] 孙庆花,于德爽,张培玉.海洋菌株y3的分离鉴定及其异养硝化-好氧反硝化特性 [J]. 环境科学, 2016,37(3):1089-1098.
[7] 聂红云,聂麦茜,白雪蕊,等.克雷伯氏菌NY 1产絮凝剂的阳离子修饰研究 [J]. 中国环境科学, 2016,36(6):1788-1793.
[8] Her J J, Huang J S. Influences of carbon source and C/N ratio on nitrate/nitrite denitrification and carbon breakthrough [J]. Bioresource Technology, 1995,54(1):45-51.
[9] 孙雪梅,李秋芬,张 艳,等.一株海水异养硝化-好氧反硝化菌系统发育及脱氮特性 [J]. 微生物学报, 2012,52(6):687-695.
[10] 司文攻,吕志刚,许 超.耐受高浓度氨氮异养硝化菌的筛选及其脱氮条件优化 [J]. 环境科学, 2011,32(11):3448-3454.
[11] Lin Y, Kong H, He Y, et al. Simultaneous Nitrification and Denitrification in a Membrane Bioreactor and Isolation of Heterotrophic N itrifying Bacteria [J]. Japanese Journal of Water Treatment Biology, 2004,40(3):105-114.
[12] 王弘宇,马 放,杨 开,等.两株异养硝化细菌的氨氮去除特性[J]. 中国环境科学, 2009,29(1):47-52.
[13] 王兆阳,陈国耀,姜 珂,等.1株耐冷兼性嗜碱好氧反硝化菌的分离鉴定及反硝化特性 [J]. 环境科学, 2013,35(6):2341-2348.
[14] 祝贵兵,彭永臻,吴淑云,等.碳氮比对分段进水生物脱氮的影响[J]. 中国环境科学, 2005,25(6):641-645.
[15] 戴 娴,王晓霞,彭永臻,等.进水C/N对富集聚磷菌的SNDPR系统脱氮除磷的影响 [J]. 中国环境科学, 2015,35(9):2636-2643.
[16] 李鹏章,王淑莹,彭永臻,等.COD/N与pH值对短程硝化反硝化过程中N2O产生的影响 [J]. 中国环境科学, 2014,34(8):2003-2009.
[17] Richardson D J, Wehrfritz J-M, Keech A, et al. The diversity of redox proteins involved in bacterial heterotrophic nitrification and aerobic denitrification [J]. Biochemical Society Transactions, 1998,26(3):401-408.
[18] 肖继波,江惠霞,褚淑祎.不同氮源下好氧反硝化菌Defluvibacter lusatiensis str.DN7的脱氮特性 [J]. 生物学报, 2012,32(20): 6463-6470.
[19] 吴建江,王兆阳,许培雅.一株高效异养硝化菌的分离、鉴定及其氨氮去除特性 [J]. 中国环境科学, 2013,33(7):1309-1315.
[20] 邹艳艳,张 宇,李明智,等.一株异养硝化-好氧反硝化细菌的分离鉴定及脱氮活性研究 [J]. 中国环境科学, 2016,36(3):887-893.
[21] Stewart B. Nitrate respiration in relation to facultative metabolismin enterobacteria [J]. Microbiology, 1988,52(2):190-232.
[22] 张培玉,曲 洋,于德爽,等.菌株qy37的异养硝化/好氧反硝化机制比较及氨氮加速降解特性研究 [J]. 环境科学, 2010,31(8):1819-1826.
[23] Taylor Shauna M, He Y, Zhao B, et al. Heterotrophic ammoniumremoval characteristics of an aerobic heterotrophic nitrifyingdenitrifying bacterium, Providencia rettgeri YL [J]. Journal of Environment Science, 2009,21(10):1336-1341.
Separation identification and the characteristics research of simu ltaneous removal of nitrogen and carbon about Marine heterotrophic nitrification and aerobic denitrification strain y6.
WANG Xiao-jing, YU De-shuang, LI Jin*,DU Song-dong, ZHOU Tong, WU Guo-dong
(School of Environmental Science and Engineering, Qingdao University, Qingdao 266071, China). China Environmental Science, 2017,37(2):686~695
A heterotrophic nitrification-aerobic denitrification strain, named y6, was isolated fromsediment of Jiaozhou Bay. It was identified asKlebsiellasp. based on the morphological, physiological and analysis of 16S rRNA gene sequence. Under different environmental conditions, measured strain y6’s growth situation and denitrification ability, so as to analyze its characteristics of simultaneous removal of nitrogen and carbon. The results showed that the optimal carbon source was sodiumcitrate; the optimal pH was 7.0; the optimal C/N was 17. In the reaction systemof NH4Cl, KNO3and NaNO2for the only nitrogen source, Strain y6had better denitrification effect. The removal efficiencies were 99.67%, 100% and 99.20%. Strain y6could efficiently remove organic matter simultaneously in the process of denitrification, with COD removal rate of 82.17%, 95.75% and 97.83%, respectively. Almost no nitrate nitrogen and nitrite nitrogen accumulation in the y6 heterotrophic nitrification process. In the different mixing ratio of reaction system, the first were aerobic denitrification of nitrate nitrogen, Subsequent were ammonia nitrogen, nitrite nitrogen and COD removal. Ammonia nitrogen removal rate was only slightly lower in the presence of the nitrate nitrogen. The nitrite nitrogen affected y6heterotrophic nitrification process. The heterotrophic nitrification of y6had no effect on aerobic denitrification.
heterotrophic nitrification-aerobic denitrification;marine bacterium;K lebsiellasp. y6;Simultaneous removal of nitrogen and carbon
X172
A
1000-6923(2017)02-0686-10
王骁静(1992-),女,山东招远人,青岛大学环境科学与工程学院硕士研究生,主要从事水污染控制方面的研究.
2016-06-23
国家自然科学基金资助项目(51278258;51478229);山东省自然科学基金资助项目(BS2015HZ007);山东省高等学校科技计划项目(J15LC61)
* 责任作者, 教授, ljin0532@126.com
