长江上游宜昌鳅 早期资源特征及影响因子分析
2017-04-06田辉伍高天珩董微微陈大庆段辛斌
田辉伍,王 涵,2,高天珩,2,董微微,2,吕 浩,2,陈大庆,段辛斌
(1.中国水产科学院长江水产研究所,武汉 430223;2.西南大学动物科技学院/生命科学学院,重庆 400715)
长江上游宜昌鳅 早期资源特征及影响因子分析
田辉伍1,王 涵1,2,高天珩1,2,董微微1,2,吕 浩1,2,陈大庆1,段辛斌1
(1.中国水产科学院长江水产研究所,武汉 430223;2.西南大学动物科技学院/生命科学学院,重庆 400715)

长江上游地形复杂,生境多样,河床海拔落差大,可开发的水能资源占全流域的87%[1],根据《金沙江干流综合规划报告》中推荐的开发方案,金沙江水电开发由19个梯级组成[2]。金沙江一期工程中的向家坝、溪洛渡两个梯级电站均为高坝巨型深水库,已于2012年10月、2013年5月相继蓄水,不仅直接阻断了鱼类的洄游路径,造成生境破碎化,还改变了保护区内的流量、流速、水温、溶氧等水动力学和水质特征,进而对长江上游鱼类特别是产漂流性卵鱼类的产卵繁殖造成直接的影响[3-4]。长江上游不仅拥有丰富的水能资源,而且具有众多全球和国家保护意义的重要生境和生态敏感区[5],其中仅长江上游珍稀特有鱼类保护区水域内就有鱼类199种,并包括胭脂鱼等3种珍稀鱼类和圆口铜鱼等66种特有鱼类,占长江特有鱼类的44%[6]。
1 材料与方法
1.1 材料与数据来源
2011—2015年,每年5月5日—7月10日在长江上游重庆市江津几江江段(N29°18′, E106°15′)进行鱼类早期资源调查。断面设置在三峡库区库尾上游35 km,向家坝下游375 km处(图1)。调查江段位于长江上游珍稀特有鱼类国家级自然保护区内,江岸平直,断面宽约500 m。每天上午和下午各进行一次采样。采样断面采样点设置以便于操作和尽可能的采集各水层样本,在断面的左岸、中间和右岸设置3个采集点,左岸、右岸两点距离岸边80 m左右,水深为3~5 m,每个采集点在表层、中层和底层采集鱼卵。
采样网具为圆锥网和弶网。圆锥网网口面积0.196 m2,网长2.5 m,网目0.5 mm,网后接圆柱形集苗器,用于定量分析,确定通过江津几江断面的鱼卵数量。弶网网口呈半圆形,网口直径1 m,网长2 m,网目0.5 mm,网口面积0.393 m2,用于定性分析。圆锥网网口处装有LS45A型流速仪,测量采样时过滤流量。同时记录每次采样时间、透明度、水温、pH、电导率等数据。水位和流量数据参考朱沱水文站,数据来源自水利部全国水雨情信息网站(http://xxfb.hydroinfo.gov.cn/ssIndex. html),朱沱水文站距离采样点75 km,水文站与采样断面之间没有主要支流汇入。

图1 长江上游鱼类早期资源采样断面图Fig.1 Locations of the sampling cross-section in the upper Yangtze River
1.2 材料处理
将所采集的鱼卵带回室内,使用奥林巴斯解剖镜SZX61观察,记录鱼卵特征及发育时期,并测定其卵径、膜径。然后继续对鱼卵进行培养,观察其早期发育情况。依据王昌燮[14]、曹文宣等[13]的方法鉴定鱼卵、仔鱼的种类。对于难以直接鉴定的鱼卵及仔鱼则用无水乙醇保存,带回实验室提取鱼卵DNA,经PCR扩增细胞色素B进行测序,使用DNA STAR软件包中的Seqman对返回序列进行检查,然后在NCBI网站(http://blast. ncbi. nlm. nih. gov / Blast. cgi)中进行比对,以序列相似度最高为鉴定标准。
1.3 数据分析
di=m/vat
式中:di表示第i个采样点采集的宜昌鳅鱼卵密度(ind./m3);m表示i次采样采到的宜昌鳅鱼卵数量;v表示网口流速(m/s);a表示网口面积(m2);t表示采集时间(s)。
档案管理及保护部门的工作人员专业素养较差,档案管理部门在进行档案管理及保护时,没有选取有专业能力的工作人员,招聘的工作人员并没有相关的工作经验也没有学习过专门的档案管理及保护知识。这样的现象不仅是对档案保护的不负责,也是对档案信息安全的不负责。
Nm=∑M+∑M’
一次采集的断面鱼卵径流量(M),计算公式如下:

相对多度(relativeabundance, RA)是表示物种优势度的参数[16]:
RA=Ni/N×100%
式中,Ni为采样期间第i种的产卵量,N为所有鱼类的产卵总量。
2 结果与分析
表1 2011—2015年长江上游江津几江江段宜昌鳅卵的出现天数、出现率、采集数量及所占比例
Tab.1 Occurrence days, occurrence rates, numbers of G.filifer eggs and proportion of totaleggs in upstream of Jiangjin section from 2011 to 2015

表1 2011—2015年长江上游江津几江江段宜昌鳅卵的出现天数、出现率、采集数量及所占比例
年份出现天数/d出现率/%鱼卵数量/粒占鱼卵总数比例/%2011182687329108320122131343641414201323343312313032014294328182125020154465674941674
2.2 早期资源量及相对多度
表2 2011—2015年长江上游宜昌鳅鱼卵日均密度与日径流量均值比较
Tab.2 Daily average density and runoffs of G.filifer eggs average comparison in theupper Yangtze River from 2011 to 2015

表2 2011—2015年长江上游宜昌鳅鱼卵日均密度与日径流量均值比较
比较因子20112012201320142015日均密度/(ind/1000m3)193±052ab167±037ab078±017a317±071bc442±069c日径流量/(104ind)10713±3014ab8275±1913ab3910±882a14703±3439bc21058±3287c
注: 同一行中不同字母上标表示存在极显著差异(P<0.01),相同字母表示差异不显著(P>0.05)。
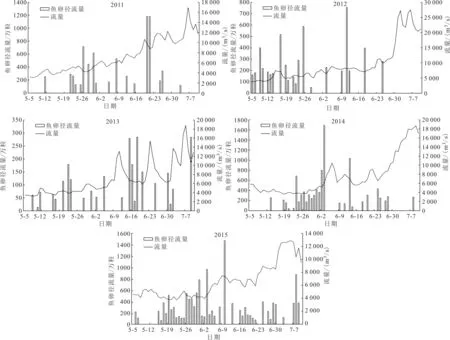
图2 5—7月流量和宜昌鳅鱼卵日径流量变化(2011—2015年)Fig.2 Daily runoff of eggs of G.filifer from May to July (2011—2015)

图3 2011—2015年长江上游江津断面宜昌鳅早期资源及相对多度Fig.3 Relative abundance and quantity of G.filifer eggs at Jiangjin in the upper Yangtze River from 2011 to 2015
通过Pearson Correlation,对2011—2015年宜昌鳅鱼卵日均密度与水温、透明度、pH、电导率、流量和两日平均流量差等环境因子进行相关性分析。发现宜昌鳅鱼卵日均密度与透明度存在极显著的正相关关系(P<0.01),与水位、流量、电导率、流速则存在极显著的负相关关系(P< 0.01),

图4 2011—2015年江津断面宜昌鳅鱼卵月平均密度Fig.4 Monthly average of G.filifer eggs density at Jiangjin section in the upper Yangtze River from 2011 to 2015
与pH、水温、两日平均流量差则无显著的相关关系(表3)。不同年份环境因子与宜昌鳅的相关系数、显著性水平存在一些差异,但是各年间鱼卵密度与透明度均为正相关,各年间鱼卵密度与水位、流量、电导率、流速均为负相关。而pH、水温、两日平均流量差等与鱼卵密度相关性不显著的环境因子的相关系数,其相关性的正负则存在不一致的情况。
各水文环境因子之间的Peason相关性见表4。可以看出流量与水位、流速、温度之间存在极显著的正相关关系(P<0.01),流量、水位与透明度间则存在极显著的负相关关系(P<0.01)。说明涨水过程是一个流量、水位、流速同步增加的过程,而流量增加的同时也常常伴随着透明度的下降。
表3 2011—2015年长江上游宜昌鳅鱼卵密度与环境因子的相关系数
Tab.3 Correlation coefficient between eggs density of G.filifer and environmental variables atJiangjin section in the upper Yangtze River from 2011 to 2015
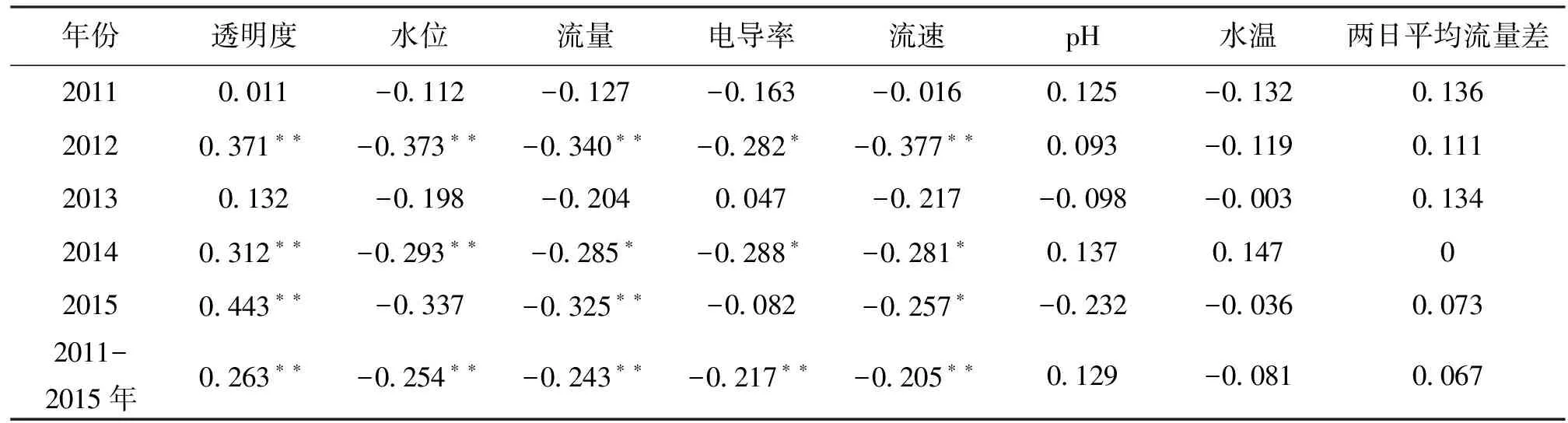
表3 2011—2015年长江上游宜昌鳅鱼卵密度与环境因子的相关系数
年份透明度水位流量电导率流速pH水温两日平均流量差20110011-0112-0127-0163-00160125-0132013620120371∗∗-0373∗∗-0340∗∗-0282∗-0377∗∗0093-0119011120130132-0198-02040047-0217-0098-0003013420140312∗∗-0293∗∗-0285∗-0288∗-0281∗01370147020150443∗∗-0337-0325∗∗-0082-0257∗-0232-003600732011-2015年0263∗∗-0254∗∗-0243∗∗-0217∗∗-0205∗∗0129-00810067
注:*表示0.05水平(双侧)上显著相关;**表示0.01水平(双侧)上显著相关

表4 长江上游江津断面各环境因子的相关性分析
注:*表示0.05水平(双侧)上显著相关;**表示0.01水平(双侧)上显著相关
3 讨论
漂流性卵的比重略大于水,需要一定的流速漂流一定的距离,完成早期发育过程,直到自由生活阶段[17]。如果流速过低,受精卵将沉入水中,窒息死亡。产漂流性卵鱼类的受精卵从产卵场漂流至育肥场生长育肥,是鱼类为扩大生存范围、加强基因交流、获得充足的饵料长期进化而成的一种生存机制,是鱼类生活史中不可或缺的一部分[18-19]。鱼类的产卵繁殖是由内源性的繁殖周期等生理因素和外源性的环境条件共同决定的,对于产漂流性卵鱼类来说,流量和水温通常被认为是影响繁殖最重要的两个环境因素[20-21]。
在适宜温度范围内,温水性鱼类受精卵的发育速度往往和水温呈正相关关系[17]。同时温度还能直接影响鱼类的产卵和排卵,正在产卵的温水鱼类如遇水温突然降低,会发生停产现象,若水温回升,又会重新产卵[26]。研究显示,溪洛渡工程的水温垂向分层现象明显,4月的下泄水温较建库前的自然水温显著偏低,四库运营之后对长江上游珍稀特有鱼类保护区水温的影响将更为明显[27]。河流水温的变化不仅会影响亲鱼的产卵繁殖,还会缩短幼鱼的生长期并降低幼鱼的越冬成活率[28]。本次研究未发现宜昌鳅产卵与水温存在相关性,可能是因为早期资源调查开始于5月,而宜昌鳅繁殖的温度下限较低,水温已经达到了宜昌鳅的产卵繁殖区间,所以暂时未观测到水温对宜昌鳅繁殖的影响。
透明度的变化不仅会对仔鱼的摄食活动产生影响,而且还会改变鱼类对光周期的反映,在非生物因子中光周期的变化对鱼类自身内部节律的调节有着重要影响[12]。宜昌鳅产卵与透明度存在极显著的正相关关系,可能是因为水位、流量较低的时候透明度一般也相对较高。另一个可能的原因是较高的透明度能为仔鱼提供良好的视线,增加摄食效率,以便仔鱼在由内源性营养转为外源性营养的关键时期,能够及时摄食开口饵料,但也因此增加了其被敌害生物捕食的概率。
水利工程的建设会改变原有的水文环境条件,并将对鱼类群落结构造成长期的影响,特别是对产漂流性卵鱼类的影响更大[20]。研究表明,产漂流性卵鱼类以鲤科为主,其中洄游性鱼类如四大家鱼、圆口铜鱼等产卵对水文条件要求最高,且产卵次数天数均较少;而小型鱼类产卵对水文条件的要求则较低[29]。向家坝工程蓄水之后,保护区河段的高流量频率、高流量历时偏差、栖息地类型多样性、垂向透水性等4项指标均出现了较大变化[30],这将对四大家鱼等洄游性鱼类的繁殖造成较大的影响,而对宜昌鳅等小型鱼类的影响则相对较小。
[1] 李红清.长江流域自然保护区建设现状与生态保护[J].长江流域资源与环境, 2011, 20(2): 150-155.
[2] 罗小勇, 陈 蕾, 李 斐.金沙江干流梯级开发环境影响分析[J].水利水电快报, 2004, 25(14): 7-10.
[3] 沈 忱, 吕平毓, 冯顺新,等.向家坝水库蓄水对下游江段溶解氧饱和度影响研究[J].淡水渔业, 2014, 44(6): 31-36.
[4] 尹正杰, 许继军, 田辉伍,等.长江上游珍稀特有鱼类保护区水文情势时空变化特征初步分析[J].淡水渔业, 2014, 44(6): 37-43.
[5] 李德旺, 李红清, 雷晓琴,等.基于GIS技术及层次分析法的长江上游生态敏感性研究[J].长江流域资源与环境, 2013, 22(5): 633-639.
[6] 危起伟.长江上游珍稀特有鱼类国家级自然保护区科学考察报告[M].北京: 科学出版社, 2012.
[7] 丁瑞华.四川鱼类志[M].成都: 四川科学技术出版社, 1994.
[8] 杨少荣, 高 欣, 马宝珊,等.三峡库区木洞江段鱼类群落结构的季节变化[J].应用与环境生物学报, 2010, 16(4): 555-560.
[9] 熊 飞, 刘红艳, 段辛斌,等.长江上游宜宾江段渔业资源现状研究[J].西南大学学报:自然科学版, 2015, 37(11): 43-50.
[10] 高志发, 赵 燕, 邓中粦.宜昌鳅仔稚鱼的形态特征[J].水生生物学报, 1988, 12(2): 186-188.
[11] 王 伟, 何舜平, 陈宜瑜.线粒体DNA d—loop序列变异与鳅亚科鱼类系统发育[J].自然科学进展, 2002, 12(1): 33-36.
[12] Pavlov D S.The downstream migration of young fishes in rivers: mechanisms and distribution[J].Folia Zoologica, 1994, 43(3): 193-208.
[13] 曹文宣, 常剑波, 乔 晔, 等.长江鱼类早期资源[M].第一版, 北京: 中国水利水电出版社, 2007.
[14] 王昌燮.长江中游“野鱼苗”的种类鉴定[J].水生生物学集刊, 1959,(3): 324-341.
[15] 易伯鲁, 余志堂, 梁秩燊,等.长江干流草、青、鲢、鳙四大家鱼产卵场的分布, 规模和自然条件[M].//易伯鲁, 余志堂, 梁秩燊,等.葛洲坝水利枢纽与长江四大家鱼.武汉: 湖北科学技术出版社, 1988: 1-46.
[16] 李振基, 陈小麟, 郑海霞, 等.生态学[M].北京:科学出版社, 2001: 200-201.
[17] 殷名称.鱼类生态学[M].北京: 中国农业出版社, 1995.
[19] Zitek A, Schmutz S, Ploner A.Fish drift in a Danube sidearm-system: II.Seasonal and diurnal patterns[J].J Fish Biol, 2004, 65(5): 1339-1357.
[20] Duan X, Liu S, Huang M et al.Changes in abundance of larvae of the four domestic Chinese carps in the middle reach of the Yangtze River, China, before and after closing of the Three Gorges Dam[J].Dev Environ Biol Fish, 2009, 28(1): 13-22.
[21] Jiang W, Liu H, Duan Z et al.Seasonal variation in drifting eggs and larvae in the upper Yangtze, China[J].Zool Sci, 2010, 27(5): 402-409.
[22] 母红霞.长江鱼类生活史对策及其早期生活史阶段对环境的适应[D].武汉: 中国科学院水生生物研究所, 2014.
[23] 王尚玉, 廖文根, 陈大庆,等.长江中游四大家鱼产卵场的生态水文特性分析[J].长江流域资源与环境, 2008, 17(6): 892-897.
[24] 易伯鲁, 梁秩燊, 余志堂,等.长江草、青、鲢、鳙四大家鱼早期发育的研究[M].//易伯鲁, 余志堂, 梁秩燊,等.葛洲坝水利枢纽与长江四大家鱼.武汉: 湖北科技出版社, 1988: 69-112.
[25] 余志堂, 梁秩燊, 易伯鲁.铜鱼和圆口铜鱼的早期发育[J].水生生物学报, 1984,(4): 371-388.
[26] 王 胜.环境温度在水产养殖中作用的研究[C].中国气象学会2006年年会“气候系统模式发展与应用”分会场论文集, 2006.
[27] 骆辉煌, 李 倩, 李 翀.金沙江下游梯级开发对长江上游保护区鱼类繁殖的水温影响[J].中国水利水电科学院学报, 2012, 10(4): 255-259.
[28] 吴清江, 程 飞.金沙江下游梯级电站运行对产漂浮性卵鱼类的影响及保护策略[C].三峡工程与长江水资源开发利用及保护国际研讨会.2008.
[29] 黎明政, 姜 伟, 高 欣,等.长江武穴江段鱼类早期资源现状[J].水生生物学报, 2010, 34(6): 1211-1217.
[30] 赵伟华, 曹慧群, 黄 茁,等.向家坝蓄水前后长江上游珍稀特有鱼类国家级自然保护区物理完整性评价[J].长江科学院院报, 2015,(6): 76-80.
[31] 姚维志, 苏胜齐, 熊 波,等.长江上游鱼类自然保护区江津段渔民生产生活现状调查[J].渔业信息与战略, 2013, 28(3): 192-198.
[32] 熊 飞, 刘红艳, 段辛斌,等.长江上游江津江段鱼类群落结构及资源利用[J].安徽大学学报:自然科学版, 2014,(3): 94-102.
[33] 李修峰, 黄道明, 谢文星,等.汉江中游产漂流性卵鱼类产卵场的现状[J].大连海洋大学学报, 2006, 21(2): 105-111.
[34] 吴 强, 段辛斌, 徐树英,等.长江三峡库区蓄水后鱼类资源现状[J].淡水渔业, 2007, 37(2): 70-75.
(责任编辑:邓 薇)
Early resources ofGobiobotiafiliferand its relationship withenvironmental factors in the upper Yangtze River
TIAN Hui-wu1, WANG Han1,2, GAO Tian-heng1,2, DONG Wei-wei1,2, LV Hao1,2,CHEN Da-qing1, DUAN Xin-bin1
( 1.YangtzeRiverFisheriesResearchInstitute,ChineseAcademyofFisheryScience,Wuhan430223,China;2.CollegeofAnimalandTechnology/CollegeofLifeScience,SouthwestUniversity,Chongqing400715,China)
In order to explore the changes of fish early resources before and after the impoundment of the first phase of the Jinsha River hydroelectric project, the investigations of fish early resources were carried out at Jiangjin section from May 5 to July 10, 2011-2015.Gobiobotiafiliferwas selected as the main research object in this experiment in the upper Yangtze River.The results show: from 2011 to 2015, during the 335 days’ survey, 1 492 eggs ofG.filiferwere collected in 135 days.The recruitments of eggs at Jiangjin section were estimated as 7.18×107, 5.54×107, 2.62×107, 9.85×107and 14.11×107ind.respectively.It was one of largest populations of fishes which spawn drifting eggs in the upper Yangtze River.The relations between drifting density of eggs and main environment factors was analyzed based on Pearson Correlation.The results indicated that the drifting average eggs density was positively correlated with water transparency (P<0.01), and negatively correlated with water level, water discharge, flow velocity and conductivity (P<0.01).From 2011 to 2015, the early resources ofG.filifershowed a first declining and then an upward trend before and after the impoundment.While its relative abundance of early resources was mounting up steadily, increasing from 4.64% to 11.05%.
Gobiobotiafilifer; the upper Yangtze River; early fish resources; environmental factors
2016-05-11;
2016-09-02
国家自然科学基金(51509262);中国长江三峡集团公司资助项目(0799570, 0799574)
田辉伍(1984-),男,助理研究员,研究方向为鱼类生态学。E-mail: taosha8413@163.com 通讯作者:段辛斌。E-mail: duan@yfi.ac.cn
S932.4
A
1000-6907-(2017)02-0071-08
