高寒草甸物种多样性和生产力对养分添加的初期响应
2017-03-23杨元武周华坤李希来周旭辉高健军赵新全
杨元武,周华坤,李希来,周旭辉,高健军,刘 玉,赵新全,叶 鑫
(1.青海大学 农牧学院,西宁 810016;2.复旦大学 生命科学学院,上海 200438;3.中国科学院 西北高原生物研究所,西宁 810001)
高寒草甸物种多样性和生产力对养分添加的初期响应
杨元武1,2,周华坤3,李希来1,周旭辉2,高健军1,刘 玉1,赵新全3,叶 鑫3
(1.青海大学 农牧学院,西宁 810016;2.复旦大学 生命科学学院,上海 200438;3.中国科学院 西北高原生物研究所,西宁 810001)
以高寒草甸为对象,通过N、P、K及其不同养分组合的添加处理,研究养分添加对高寒草甸生态系统物种组成、物种多样性和生产力的影响。结果表明:养分添加改变群落物种组成和群落中的优势种,除P、K单独添加外,其他养分及其组合添加使草地群落物种丰富度显著减小,物种多样性和均匀度变化不大;N、P及其不同养分组合添加极显著增加群落地上生物量;N、P是高寒草地生态系统草地生产力的主要限制性养分,人为输入土壤养分将在短期内影响高寒草地生态系统的物种组成、物种多样性以及生产力。
养分添加;地上生物量;物种多样性;高寒草甸
植被与环境的关系一直是生态学研究的重点问题。在近些年的生态系统功能研究中,自然状态下的草地生态系统越来越受到关注[1]。高寒草地处于中国青藏高原区生态脆弱带,人类频繁干扰和气候变化引起的草地退化给高寒草地生态系统带来深远的影响。自然群落中的物种组成是物种对环境长期适应的结果[2]。土壤作为植物生长的重要物质基础,其物理、化学性质的不同,土壤母质的不同,都可能影响生长于其中的植物[3],从而影响到物种多样性。物种多样性和生产力是生态系统结构和功能的重要指标。N、P、K素是土壤肥料的三要素,也是大多数陆地生态系统初级生产力的主要限制因子[4-5],但过多的物质输入将使生态系统达到饱和状态,以致影响许多生态系统过程,如生态系统生产力、物种组成以及多样性等[6-7]。目前,N素添加对陆地生态系统生产力和生物多样性影响的研究已有很多报道[8-9]。欧洲和北美一些发达国家和地区对于草地生态系统对全球性氮输入增加的响应进行了大量的研究[10-11],Mitchell[12]在阿拉斯加苔原的长期施肥试验表明,施肥使植物年平均生物量增加1倍,但造成了深层土壤碳氮的损失。Clemmensen 等[13]研究灌丛草地丛枝菌根真菌丰富度对施肥和长期变暖的响应。Grellmann[14]在挪威北部的石楠灌丛的不同土地利用方式下进行施肥(N/P/K)试验,结果表明不同施肥方式对石楠灌丛的生长产生不同的影响。Mitchelle[12]在阿拉斯加研究土壤原生动物对N/P添加的响应,进而影响土壤有机质的转化和营养的可利用性。
国内有关磷素和钾素添加对草地生态系统影响的研究相对较少[15-16]。沈振西等[17]研究青藏高原高寒矮嵩草草甸植物类群对模拟夏季增减雨量、冬春增雪及增施氮肥的响应,结果表明矮嵩草草甸高强度施氮对不同植物类群有显著影响。牛克昌等[18]以青藏高原东部高寒草甸植物群落的24种主要组分为材料,研究表明施肥对多年生草本植物繁殖分配有显著影响。然而,养分对植物群落物种多样性的影响机制以及生物多样性维持草地生产力的机制目前尚不清楚。为此,本研究以高寒草甸为例,通过人为土壤养分添加输入,研究探讨N、P、K素及其组合养分的添加对高寒草地生态系统物种多样性和生产力的影响,旨在揭示高寒草地物种多样性和生产力对土壤养分添加的响应,为高寒草地的适应性管理提供科学依据。
1 材料与方法
1.1 研究区概况
研究地点位于中国科学院海北高寒草甸生态系统开放试验站,站区位于青藏高原东北隅祁连山东段北支冷龙岭南(37°29′~37°45′N, 101°12′~101°33′E)。该站区地形以低山、丘陵、滩地和河流阶地为主,海拔3 200~3 500 m,最高峰海拔5 600 m。由于地理位置及地形条件的影响而具有明显的高原大陆性气候特点,太阳辐射强,全年太阳辐射为6.93×108J/cm,年均日照时数为2 672.6 h,日照率达60%,年温差较小而日温差较大,年均温为-1.7 ℃,1月均温为-14.8 ℃, 绝对最低气温达-35.2 ℃,最热月7月均温为9.9 ℃,绝对最高气温为 23.7 ℃,≥5 ℃积温为1 176.0 ℃。年均降水量为 614.8 mm, 多集中于 6-8月,历年平均最大降水量在7月,为126.3 mm,12月平均降水量最小,仅有4.0 mm,无绝对无霜期。由于独特的地理气候和长期过度放牧,该区形成特殊的矮嵩草草甸。矮嵩草草甸是该区分布最普遍的草地类型之一,形成的群落结构复杂,种类组成丰富,群落总盖度约85%,土壤含水量约30%。主要优势种为矮嵩草(Kobresiahumilis),伴生种有异针茅(Stipaaliena)、垂穗披碱草(Elymusnutans)、早熟禾(Poa.spp.)、羊茅(Festucaspp.)、芝剪股颖(AgrostistriniiTurcz.)、高山嵩草(Kobresiapygmaea)、美丽凤毛菊(Saussureasupera)、麻花艽(Gentianastraminea)、粗喙苔草(Carexsczbrirostris)、苔草(Carexspp.)、矮火绒草(Leontopodiumnanum)、雪白委陵菜(Potentillanivea)、鹅绒委陵菜(Potentillaanserina)、高山唐松草(Thalictrumalpinum)、甘肃棘豆(Oxytropiskansuensis)、异叶米口袋(Gueldenstaedtiadiversifolia)、甘肃马先蒿(PedicularisKansuensis)、洽草(Koeleriacristata)、小花刺参(Morinachineasis)、花苜蓿(TrigonellaruthenicaL.)、藏异燕麦[Helictotrichontibeticum(Roshev.)]、高山紫菀(AsteralpinusLinn.)、双叉细柄草(PtilagrostisdichotomaKeng ex Tzel.)及多种毛茛 (Ranuncuulusspp.)等[19]。
1.2 试验设计与方法
选择植被生长均匀,物种组成较为一致的平坦地段,采用完全随机区组设计试验,以养分的不同组合N、P、K、NP、NK、PK、NPK和对照(CK)共8个处理,每个处理重复6次,共计48个小区,每个小区面积为36 m2(6 m×6 m)。裂区之间设置2 m的缓冲带,小区之间设置1 m的缓冲带。施肥时间为5月15日-5月17日,所施肥料为氮肥、磷肥、钾肥,施肥量按照国际养分网络(Nutrient Network)上认可的标准进行配比[20-21]。每年的施肥量经折算后分别以尿素24.4 g/m2、重过磷酸钙40.7 g/m2、K2SO422.3 g/m2的形式手工均匀地撒在小区试验地地表,2012年5月和2013年5月各施1次。
1.3 取样及指标测定
1.3.1 取样方法 于2014-08-23取样,植物地上生物量达到高峰期时进行群落学调查。样方面积为1 m2,调查项目包括植物种、密度、盖度、频度及群落地上生物量。地上生物量鲜质量指齐地面剪取样方内植物地上部所称量的,之后在80 ℃下烘10 h后称干质量。
1.3.2 植物多样性测度 本研究中,物种丰富度指数用1 m2样方内出现的物种数表示。
相对重要值=(相对密度+相对盖度+相对频度)/3
式中:相对密度为某一物种的密度占全部物种密度之和的百分比;相对盖度为某一物种的盖度占所有盖度之和的百分比;相对频度为某一物种的频度占全部物种频度之和的百分比[22]。
物种多样性选用 Shannon-Wiener多样性指数(H)进行多样性的测度[23]:
H=-ΣPilnPi
式中:Pi为样方中种i的相对重要值,Pi=Ni/N;Ni为种i的绝对重要值;N为种i所在样方的各个种的重要值之和。
同时计算群落的Pielou均匀度指数(J)[23]:
J=H/lnS
式中:S为群落样方中所有物种的个体数之和。
1.4 数据处理
采用 Microsoft excel 2007和SAS 9.0软件进行数据处理分析。利用一元方差分析方法(one-way ANOVA )对试验结果进行分析,先用 Levene进行方差齐性检验,如数据满足齐性检验,则利用 LSD 多重比较法进行显著性检验,否则利用 Tamhane分析。
2 结果与分析
2.1 养分添加对高寒草地群落物种组成和群落结构的影响
由表1可以看出,养分添加对群落物种组成有明显的影响。在CK中,垂穗披碱草、甘肃棘豆和小花刺参为优势种,其重要值分别为28.6、24.2和10.4,其优势度总和占群落总优势度的63.2%;在N、NK和NPK添加中,垂穗披碱草、芒剪股颖和早熟禾的重要值位居前3位,其优势度总和分别占群落总优势度的 62.8%、76.8%和73.8%;在P添加中,垂穗披碱草、甘肃棘豆和美丽凤毛菊成为群落优势种,在K添加中,垂穗披碱草、美丽凤毛菊和麻花艽成为群落优势种,在NP添加中,垂穗披碱草、藏异燕麦和芒剪股颖成为群落优势种;在PK添加中,甘肃棘豆、美丽凤毛菊和麻花艽成为群落优势种。除PK外,其他养分添加和CK中垂穗披碱草的优势度最高,但与CK相比,N、NP、NK和NPK处理中其优势度极显著增大,而P、K处理与CK相比,其优势度无显著变化,PK的添加使垂穗披碱草的优势度极显著降低;除PK添加外,其他所有养分及其组合添加极显著降低了甘肃棘豆的优势度。从以上结果可以看出:不同的养分添加中,群落中优势种的优势程度发生变化,亚优势物种有所变化;不同养分的增加,群落中相对重要值占前 3位优势种的物种优势度总和占群落总优势度的比例显著高于CK; N、NK、NPK组合养分利于大多数禾本科类植物的生长,而P、K、PK养分利于阔叶杂类草的生长。总之,养分添加导致了群落中物种优势度的显著变化。
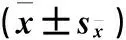
表1 不同养分添加下群落物种组成及其相对重要值Table 1 Species composition and relative importance value under different nutrition additions
注:同一行不同大写字母代表不同处理间差异极显著(P<0.01)。
Note:Different capital letters in same row represent significant difference between treatments (P<0.01).
2.2 养分添加对高寒草地物种多样性的影响
2.2.1 群落物种丰富度的变化 群落物种丰富度、多样性和均匀度是群落的物种重要特征。由图1可以看出,除P、K添加处理对高寒草地物种丰富度影响不显著外,其他养分添加处理群落物种丰富度显著低于CK,尤其在NK和NPK添加处理中,物种数从CK的12.66种减少到8种和7种,分别减少37%和45%。说明在N及其N素组合养分添加时,群落中部分高大禾草的强烈生长抑制了矮小落叶植物的生长,群落的物种数减少。表明养分添加对草地植物群落物种丰富度的影响较大。
2.2.2 群落物种多样性和均匀度的变化 群落物种多样性和均匀度是指群落中的物种数目以及各物种个体数目分配的均匀程度,也是物种多样性的重要指标。
由图2和图3可以看出,在NP养分组合添加时,群落物种H和J显著高于CK,二者值分别从CK的1.90和0.77增加到2.00和0.82;其余所有养分及其组合添加对高寒草地物种H和J影响不显著。说明除NP组合养分添加外,其余养分及其组合添加对群落各物种数目的分配无显著影响。表明养分添加初期对草地植物群落物种多样性及均匀度的影响不大。
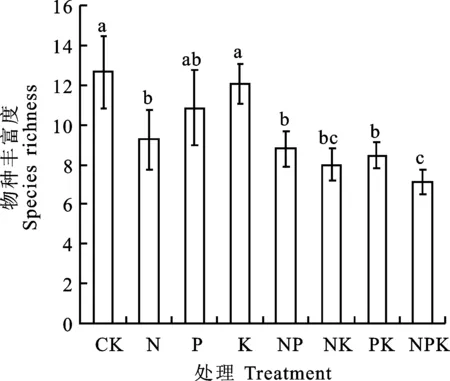
不同小写字母代表不同处理间差异显著(P<0.05),下同。
Different lowercase letters represent significant difference between treatments at 0.05 level, the same below.
图1 不同养分添加群落的物种丰富度
Fig.1 Species richness under different nutrition additions
图2 不同养分添加群落的物种多样性Fig.2 Species diversity under different nutrition additions

图3 不同养分添加群落的物种均匀度Fig.3 Species evenness under different nutrition additions
2.3 养分添加对高寒草甸植物个体地上生物量的影响
试验分析了群落中6种主要植物个体生物量对养分添加响应,结果表明(图4),禾本科植物垂穗披碱草(Elymusnutans)、早熟禾(Poa.spp.)和芒剪股颖(AgrostistriniiTurc.)的地上生物量对N添加的响应最大,添加N时,这3种植物个体地上生物量显著高于CK,添加NP、NK和NPK对其影响次之,但仍显著大于CK,垂穗披碱草(Elymusnutans)的地上生物量对单独P和K的添加无显著响应。莎草科植物矮嵩草(Kobresiahumilis)个体地上生物量则在添加P、K后表现出显著增加,对单独N的添加无显著响应。美丽凤毛菊(Saussureasupera)个体生物量在添加NP后显著增加,而其他养分添加后其生物量均显著降低。雪白委陵菜(Potentillanivea)的个体生物量在添加N后显著增加,添加K后则显著降低,对P的添加则无显著响应变化。
2.4 养分添加对高寒草甸群落地上生物量的影响
为了检验养分添加在群落水平上对生物量的影响,分析了不同养分添加后群落生物量的变化,方差分析结果表明,与对照相比,除添加K、PK养分对草地群落地上生物量的影响不显著外,其他养分及其组合添加都极显著增加了群落地上生物量(图5),养分添加对生物量鲜质量和干质量的影响情况一致。其中当N和NPK组合添加时,群落地上生物量达最大,干质量分别为904.5 g/m2和779.7 g/m2,分别较对照提高103%和75%。说明养分组合添加对草地植物群落生产力有很大的影响。
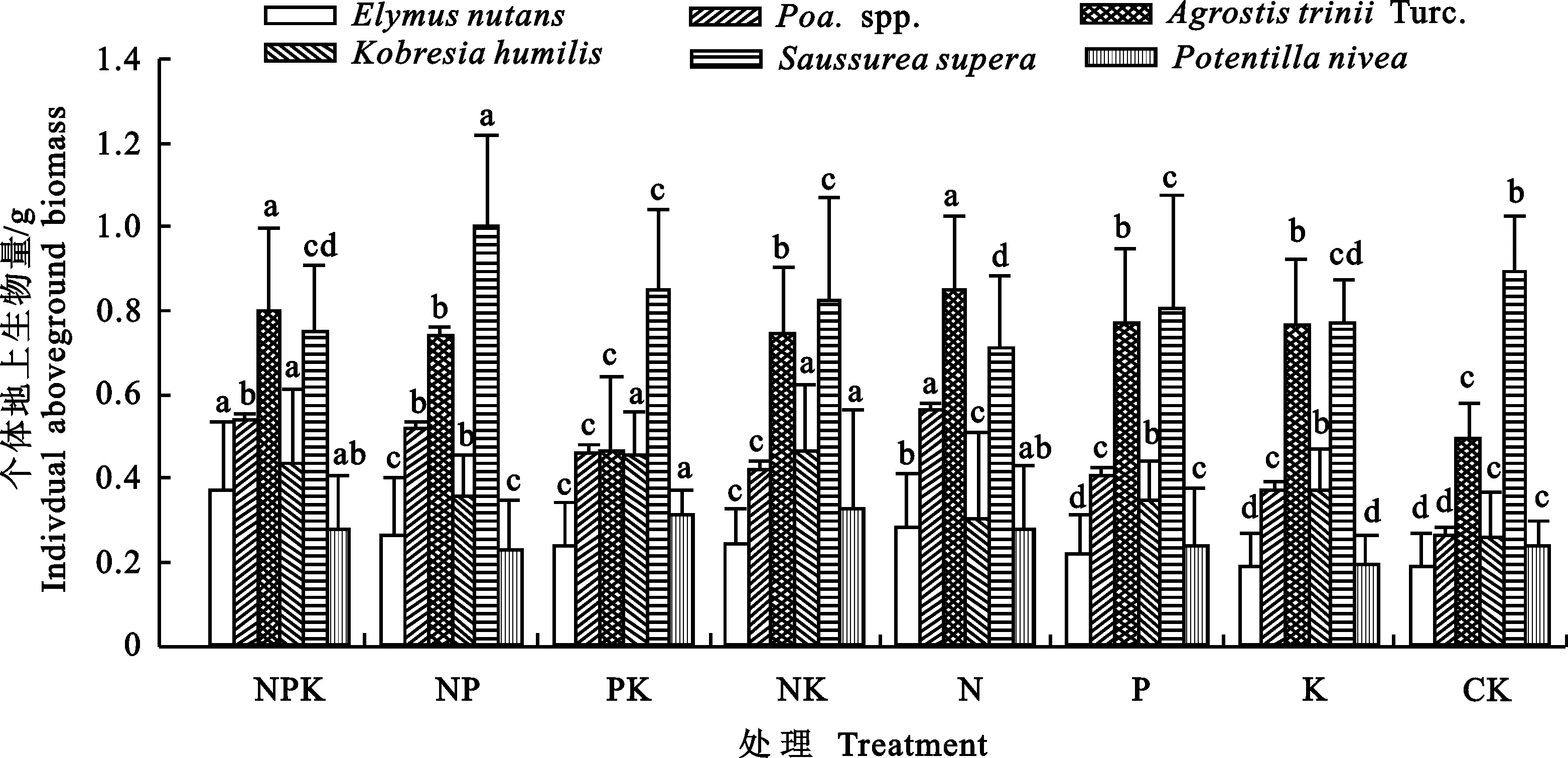
图4 不同养分添加对主要植物个体地上生物量的影响Fig.4 Effects of nutrition addition on individual aboveground biomass of main plants
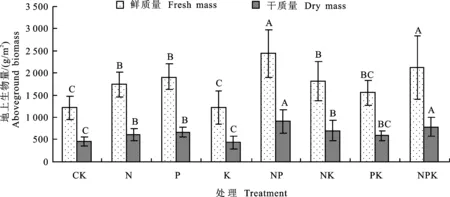
不同大写字母代表不同处理间差异极显著(P<0.01)。
Different capital letters represent significant difference between treatments(P<0.01).
图5 不同养分添加下高寒草甸群落的地上生物量
Fig.5 Biomass of aboveground plant in alpine grassland under different nutrition additions
3 讨论与结论
关于养分添加对群落物种组成及其物种多样性影响的研究报道很多[9,24]。Tilman等[24]认为,与优势种生物量的增加相比,物种组成的变化或许是群落对养分添加更为重要的一种响应。然而,目前有关养分添加导致物种组成和群落结构改变的观点并不一致。一些研究认为,物种间养分竞争力的差异是导致优势种变化的主要因素[25];而另一些研究表明,物种对光资源的竞争是导致物种组成发生变化的主要因素,土壤养分使物种对营养的竞争转向对光资源的竞争[24,26]。生态系统中植物限制性资源的转化,改变了种间的竞争格局和过程,使一些物种逐渐被另一些物种所取代,进而使生态系统的组成和结构发生改变。物种对养分添加的响应不仅与物种本身的功能特性有着密切的关系[1],还受物种所在群落环境的影响,是物种适应性和群落环境变化相互作用的结果[27-28]。
本研究中,养分添加可能解除了群落中某些物种的养分限制,比如禾草状草类的生长显著加快,从而通过竞争改变群落的物种组成(表1)。群落物种多样性对养分添加的响应机理比较复杂,多数研究认为养分添加会导致物种多样性下降[29-31],本研究也证实这个观点。Foster等[32]在美国密西根州的研究结果表明,2个生长季的氮素添加降低了物种的丰富度。本研究结果表明,养分添加对高寒草甸植物群落物种丰富度的影响较大,而对物种多样性及均匀度的影响不大。养分添加使物种丰富度显著降低,可能由于养分添加引发的种间竞争力的不平衡导致群落物种多样性减小[29]。N和P是高寒草地生态系统初级生产力的主要限制因子(图4),大量N素和P素的添加增加了植物可利用养分的水平,从而消除了生态系统对养分的限制,养分利用效率高的植物在地下资源的竞争中占有优势,得以迅速生长,同时该类植物的生长减小了群落下层的光照强度,遮蔽了其他一些矮小物种或不耐阴物种,使之在光的竞争中处于劣势,在群落中的数量逐渐减少或者消失,导致群落物种多样性下降。
土壤营养限制着陆地生态系统的净初级生产力[4]。因此,在研究生态系统生产力的限制因子时,人们常常采用N素添加的方法。当N素添加增加了净初级生产力时,该生态系统被认为受N素限制[5]。Ditommaso等[30]认为在多数研究中,养分添加促进植物生长,提高植物生产力,Lebauer等[5]通过整合分析126个N素添加试验,评估陆地生态系统净初级生产力的N素限制状况,结果表明,大多数陆地生态系统的生产力受N素的限制,且不同类型的生态系统受养分种类和数量的限制不同。本研究中,施肥2 a后N素和P素添加显著提高了群落地上生物量(图4),说明二者是高寒草地生态系统植物生长的主要限制因子,而K素的添加对植物群落生产力的影响不大,说明高寒草地土壤K素为非限制性因子,这与其他一些研究结果一致[33]。然而,养分添加对植物地下生物量的影响不容忽视,因为营养的可利用性会影响到植物地上与地下生物量的分配。今后的相关试验应当对地下生物量同时进行分析讨论。
总之,养分添加改变了高寒草甸群落物种组成和群落中的优势种,降低了物种丰富度,不同物种的地上生物量对养分添加的响应不同,但养分添加显著促进了群落地上生物量的增加。由此认为,人为添加养分将影响高寒草地生态系统中物种组成、物种多样性以及生产力,高寒草甸物种多样性和生产力对不同类型养分的短期响应发生了明显分异,但是养分添加对该物种的长期影响有待进一步监测和研究。
Reference:
[1] 万宏伟,杨 阳,白世勤,等.羊草草原群落6 种植物叶片功能特性对氮素添加的响应[J].植物生态学报,2008,32(3):611-621.
WAN H W,YANG Y,BAI SH Q,etal.Variations in leaf functional traits of six species along a nitrogen addition gradient inLeymuschinensissteppe is Inner Mongolia [J].ActaPhytoecologicaSinica,2008,32(3):611-621(in Chinese with English abstract).
[2] 宋理明,娄海萍.青海湖地区天然草地土壤湿度研究[J].中国农业气象,2006,27(2):151-155.
SONG L M,LOU H P.Studies on soil moisture dynamics of natural grassland in Qinghai lake region [J].ChineseJournalofAgrometeorology,2006,27(2):151-155(in Chinese with English abstract).
[3] TILMAN D,WEDIN D,KNOPS J.Productivity and sustain ability influenced by biodiversity in grassland ecosystems [J].Nature,1996,379(22):718-720.
[4] VITOUSEK P M,HOWARTH R W.Nitrogen limitation on land and in the sea:how can it occur? [J].Biogeochemistry,1991,13(2):87-115.
[5] LEBAUER D S,TRESEDER K K.Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed [J].Ecology,2008,89(2):371-379.
[6] ABER J D,MCDOWELL W H,NADELHOFFER K J.Nitrogen saturation in temperate forest ecosystems[J].Bioscience,1998,48(1):921-934.
[7] VITOUSEK P M,ABER J D,HOWARTH R W,etal.Human alteration of the global nitrogen cycle:sources and consequences [J].EcologicalApplications,1997,7(3):737- 750.
[8] BOBBINK R,HORNUNG M ,ROELOFS J G.The effects of air-borne nitrogen pollutants on species diversity in natural and semi-natural European vegetation [J].JournalofEcology,1998,86(5):717-738.
[9] XIA J Y,WAN S Q.Global response patterns of terrestrial plant species to nitrogen addition [J].NewPhytologist,2008,179(2):428-439.
[10] WEDIN D A ,TILMAN D.Influence of nitrogen loading and species composition on the carbon balance of grasslands [J].Science,1996,274(5293):1720-1723.
[11] STEVENS C J,DISE N B,MOUNTFORD J O,etal.Impact of nitrogen deposition on the species richness of grasslands [J].Science,2004,303(5665):1876-1879.
[12] MITCHELL E A D.Response of testate amoebae (protozoa) to N and P fertilization in an arctic wet sedge tundra [J].ArcticAntarcticandAlpineResearch,2004,36(1):78-83.
[13] CLEMMENSEN K E,MICHELSEN A,etal.Increased ectomycorrhizal fungal abundance after long-term fertilization and warming of two arctic tundra ecosystems [J].NewPhytologist,2006,171(2):391-404.
[14] GRELLMANN D.Plant responses to fertilization and exclusion of grazers on an arctic tundra heath [J].Oikos,2002,98(2):190-204
[15] XU W H,WAN SH Q.Water- and plant-mediated responses of soil respiration to topography,fire,and nitrogen fertilization in a semiarid grassland in northern China [J].SoilBiologyandBiochemistry,2008,40(3):679-687.
[16] NIU S,WU M,HAN Y I,etal.Nitrogen effects on net ecosystem carbon exchange in a temperate steppe [J].GlobalChangeBiology,2010,16(1):144-155.
[17] 沈振西,周兴民,陈佐忠,等.高寒矮嵩草草甸植物类群对模拟降水和施氮的响应[J].植物生态学报,2002,26(3):288-294.
SHEN ZH X,ZHOU X M,CHEN Z ZH,etal.Response of plant groups to simulated rainfall and nitrogen supply in alpineKobresiahumilis[J].ActaPhytoecologicaSinica,2002,26(3):288-294(in Chinese with English abstract).
[18] 牛克昌,赵志刚,罗燕江,等.施肥对高寒草甸植物群落组分种繁殖分配的影响[J].植物生态学报,2006,30(5):817-826.
NIU K CH,ZHAO ZH G,LUO Y J,etal.Fertilization effects on species reproductive allocation in an alpine meadow plant community [J].ActaPhytoecologicaSinica,2006,30(5):817-826(in Chinese with English abstract).
[19] 杨元武,李希来,李积兰,等.高寒草甸矮嵩草对放牧扰动的生长反应[J].西北农业学报,2011,20(9):18-24.
YANG Y W,LI X L,LI J L,etal.Growth response ofKobresiahumilisto grazing disturbance on alpine meadow [J].ActaAgriculturaeBoreali-occidentalisSinica,2011,20(9):18-24(in Chinese with English abstract).
[20] ELSER J J,BRACKEN M E,CLELAND E E,etal.Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater,marine and terrestrial ecosystem [J].EcologyLetters,2007,10(12):1135-1142.
[21] CLARK C M,CLELAND E E,COLLINS J E,etal.Environmental and plant community determinants of species loss following nitrogen enrichment [J].EcologyLetters,2007,10(7):596-607.
[22] 何兴东,高玉葆,刘惠芬.重要值的改进及其在羊草群落分类中的应用[J].植物研究,2004,24(4):466-472.
HE X D,GAO Y B,LIU H F.Amending of importance value and its application on classification ofLeymuschinensiscommunities [J].BulletinofBotanicalResearch,2004,24(4):466-472(in Chinese with English abstract).
[23] 宋永昌.植被生态学[M].上海:华东师大出版社,2001.
SONG Y CH.Vegetation Ecology [M].Shanghai:East China Normal University Press,2001(in Chinese).
[24] TILMAN D.Secondary succession and the pattern of plant dominance along experimental nitrogen gradients [J].EcologicalMonographs,1987,57(3):189-214.
[25] AERTS R.Interspecific competition in natural plant communities:mechanisms trade-offs and plant-soil feedbacks [J].JournalofExperimentalBotany,1999,50(330):29-37.
[26] TILMAN D.Plant Strategies and the Dynamics and Structure of Plant Communities [M].New Jersey:Princeton University Press,1988.
[27] KNOPS J M,REINHART K.Specific leaf area along a nitrogen fertilization gradient [J].AmericanMidlandNaturalist,2000,144(2):265-272.
[28] PENNINGS S C,CLARK C M,CLELAND E F,etal.Do individual plant species show predictable responses to nitrogen addition across multiple experiments? [J].Oikos,2005,110(3):547-555.
[29] RAJANIEMI T K.Why does fertilization reduce plant species diversity testing three competition-based hypotheses [J].JournalofEcology,2002,90(2):316-324.
[30] DITOMMASO A,AARSSEN L W.Resource manipulations in natural vegetation:a review [J].PlantEcology,1989,84(1):9-29.
[31] GOUGH L,OSENBERG C W,GROSS K L,etal.Fertilization effects on species density and primary productivity in herbaceous plant communities [J].Oikos,2000,89(3):428-439.
[32] FOSTER B L,GROSS K L.Species richness in successional grassland:effects of nitrogen enrichment and plant litter [J].Ecology,1998,79(8):2593-2602.
[33] 于占源,曾德慧,姜凤岐,等.半干旱区沙质草地生态系统循环关键过程对水肥添加的响应[J].北京林业大学学报,2006,28(4):45-50.
YU ZH Y,ZENG D H,JIANG F Q,etal.Responses of key carbon cycling processes to the addition of water and fertilizers to sandy grassland in semi-arid region [J].JournalofBeijingForestryUniversity,2006,28(4):45-50(in Chinese with English abstract).
(责任编辑:史亚歌 Responsible editor:SHI Yage)
Initial Response of Species Diversity and Productivity to Nutrients Addition on Alpine Meadow
YANG Yuanwu1, ZHOU Huakun3, LI Xilai1, ZHOU Xuhui2, GAO Jianjun1,LIU Yu1, ZHAO Xinquan3and YE Xin3
(1.Agricultural and Animal Husbandry College, Qinghai University, Xining 810016, China;2.School of Life Sciences,Fudan University, Shanghai 200438, China; 3.Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810001, China)
Species diversity and productivity are the important indices of the structure and functioning of ecosystems.With an alpine grassland as tested object, this paper studied its species composition, species diversity, and productivity under effects of different nutrients (N/P/K) and their composition addition.Nutrients addition altered the species composition and the dominant species in the community.The species richness decreased significantly in different nutrients and their composition addition except P/K adding lonely.The diversity and evenness had little change.Aboveground biomass of community increased significantly under N/P and their different composition.N and P are main limited nutrients of productivity in alpine grassland eco-system.It indicates that artificial soil nutrients inputting would affect the species composition, species diversity, and productivity of alpine grassland ecosystems.
Nutrients addition; Aboveground biomass; Species diversity; Alpine meadow
YANG Yuanwu,male,Ph.D candidate.Reseach area:grassland ecology and environmental protection.E-mail:yyuanwu@163.com
日期:2016-12-29
网络出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20161229.1005.002.html
2016-01-22
2016-03-27
青海省科技厅应用基础研究计划(2014-ZJ-779);国家自然科学基金(31560151);教育部长江学者和创新团队发展计划(IRT13074);科技部国际科技合作计划 (2015DFG31870,2011DFG93160);国家“973”计划(2009CB421102-04)。
杨元武,男,副教授,博士研究生,研究方向为草地生态与环境保护。E-mail: yyuanwu@163.com
Q948.113
A
1004-1389(2017)02-0159-08
Received 2016-01-22 Returned 2016-03-27
Foundation item Applied Basic Research of Qinghai Provincial Department of Science and Technology(No.2014-ZJ-779);National Natural Science Foundation of China(No.31560151);Changjiang Scholars and Innovation Team Development Plan of Ministry of Education of P.R.China(No.IRT13074);International Scientific and Technological Cooperation Program of Ministry of Science and Technology(No.2015DFG31870,No.2011DFG93160);National 973 Program(No.2009CB421102-04).
