盐胁迫下巴西蕉幼苗CaM含量的变化
2017-03-23王文昌周双云乔飞吉福桑李
王文昌++周双云++乔飞++吉福桑++李元元++李新国
摘 要 研究巴西蕉幼苗在60 mmol/L NaCl人工模拟的盐胁迫0、4、8、12、24和48 h不同时间下,通过增加外源CaCl2和钙离子螯合剂EGTA处理后,测定其叶片和根的钙调蛋白(CaM)含量的变化。结果表明,与对照处理相比,在NaCl胁迫过程中,叶片CaM含量在4、8、12 h显著增加而在24和48 h显著减少;根的CaM含量则是在4、12、24 h显著增加,在8 h显著降低。在NaCl胁迫中添加CaCl2处理能显著提高CaM含量,而在NaCl胁迫中添加EGTA处理能显著降低CaM含量。以上说明,通过CaM的含量变化,钙信号系统参与香蕉的耐盐调控。
关键词 巴西蕉幼苗 ;盐胁迫 ;CaM含量
中图分类号 S668.1 文献标识码 A Doi:10.12008/j.issn.1009-2196.2017.02.003
The Change of CaM content in Brazil Banana Seedlings under Salt Stress
WANG Wenchang1,2) ZHOU Shuangyun2)
QIAO Fei3) JI Fusang1) LI Yuanyuan1) LI Xinguo1)
(1 College of Horticulture and Landscape Architecture,HainanUniversity, Haikou, Hainan 570228;
2 Horticultural Research Institute, Guangxi Academy of Agricultural Sciences, Nanning 530007;
3 Tropical Crops Genetic Resource Institute, CATAS, Danzhou, Hainan 571737)
Abstract Brazil banana seedlings were treated with salt under 60 mmol/L NaCl salt stress environment at 0, 4, 8, 12, 24 or 48 h and added with exogenous CaCl2 and calcium chelator EGTA to determine their CaM content in roots and leaves. The results showed that the CaM content of the leaves significantly increased at 4 h, 8 h or 12 h and significantly decreased at 24 h or 48 h under NaCl stress as compared to the control. The CaM content of the roots significantly increased at 4 h, 12 h or 24 h, and significantly decreased at 8 h. The CaM content could significantly increased by adding CaCl2 under the NaCl stress, and significantly decreased by adding EGTA under the NaCl stress. This indicates that calcium signaling system may be involved in the salt regulation of banana through the change of CaM content.
Keywords Brazil banana seedling ; Salt stress ; CaM content
钙调蛋白(CaM)是真核细胞中最为保守的蛋白之一,也是研究最广泛和深入的Ca2+传感蛋白[1],它是一类分布最广、具有重要生物学功能的Ca2+受体蛋白。CaM自身没有激酶活性,必须通过结合Ca2+形成Ca2+-CaM复合体,才能诱导分子构象发生改变从而激活下游各类靶蛋白[2]。相关研究表明[3-5],當受到逆境胁迫时,植物能通过提高细胞质游离Ca2+的浓度,并通过 Ca2+与CaM 结合,启动一系列生理生化过程,从而在作物对逆境胁迫的感受、传递、响应和适应过程中发挥重要作用。钙作为重要的第二信使,参与许多植物对逆境信号的转导。Ca2+-CaM信号途径在植物应对各种胁迫及调控生长发育中起着不可忽视的作用[6]。我们前期研究证明,钙参与香蕉调控盐胁迫生理机制。但是,在钙信号启动过程中,CaM含量的变化还未见报道[7-8]。本研究以巴西蕉幼苗为试材研究盐胁迫下CaM含量的变化,旨在为研究香蕉逆境胁迫下的反应信号传导变化及适应调节提供依据。
1 材料与方法
1.1 材料
材料选自中国热带农业科学院种苗组培中心的巴西蕉(Musa AAA Cavendish var. Brazil)幼苗,苗高约25 cm,粗度基本一致,五叶一心,心叶未展开,无病虫害。将袋装巴西蕉幼苗取出,经去土洗净根系后转入1/2 Hoagland营养液进行水培,每3 d更换一次培养液。培养条件为:光照强度75 μmol/(m2·s),光照时间14 h/d,昼夜温度27℃/21℃,相对湿度85%~90%,培养期间用电动气泵全天持续对营养液通气。预培养6 d后设置对照、60 mmol/L NaCl、60 mmol/L NaCl+5 mmol/L CaCl2和60 mmol/L NaCl+10 mmol/L EGTA 4种处理。分别处理0、4、8、12、24和48 h后取材。分别称取0.10 g的叶片和根,液氮速冻,-80℃下保存备用。
1.2 方法
1.2.1 试验设计
巴西蕉幼苗叶片和根的CaM含量采用 ELISA(Enzyme-linked immunosorbent assay,酶联免疫法)测定,按照上海酶联生物科技有限公司提供的植物CAM ELISA试剂盒方法进行。CaM含量的提取,参考罗充[9]的方法,略作修改,测定时取准备好的冷冻各样品0.10 g,用样品提取液(50 mmol/L Tris-HCl缓冲液pH=7.5;1 mmol/L的EGTA;0.15 mol/L NaCl;0.5 mmol/L PMSF;20 mmol/L的NaHCO3)稀释10倍,在冰浴中研磨并匀浆,然后倒入冷冻离心机的离心管内,在92℃下加热3 min后又立即放入冰浴中冷却,离心管相互平衡后在冷冻离心机中离心(4℃、0.5 h、12 000 r/min),上清液吸入洗净的离心管内,低温-4℃冰箱保存。
1.2.2 数据处理
数据采用DPS 7.05软件进行差异显著性检验,多重比较分析采用DUNCAN新复极差法。
2 结果与分析
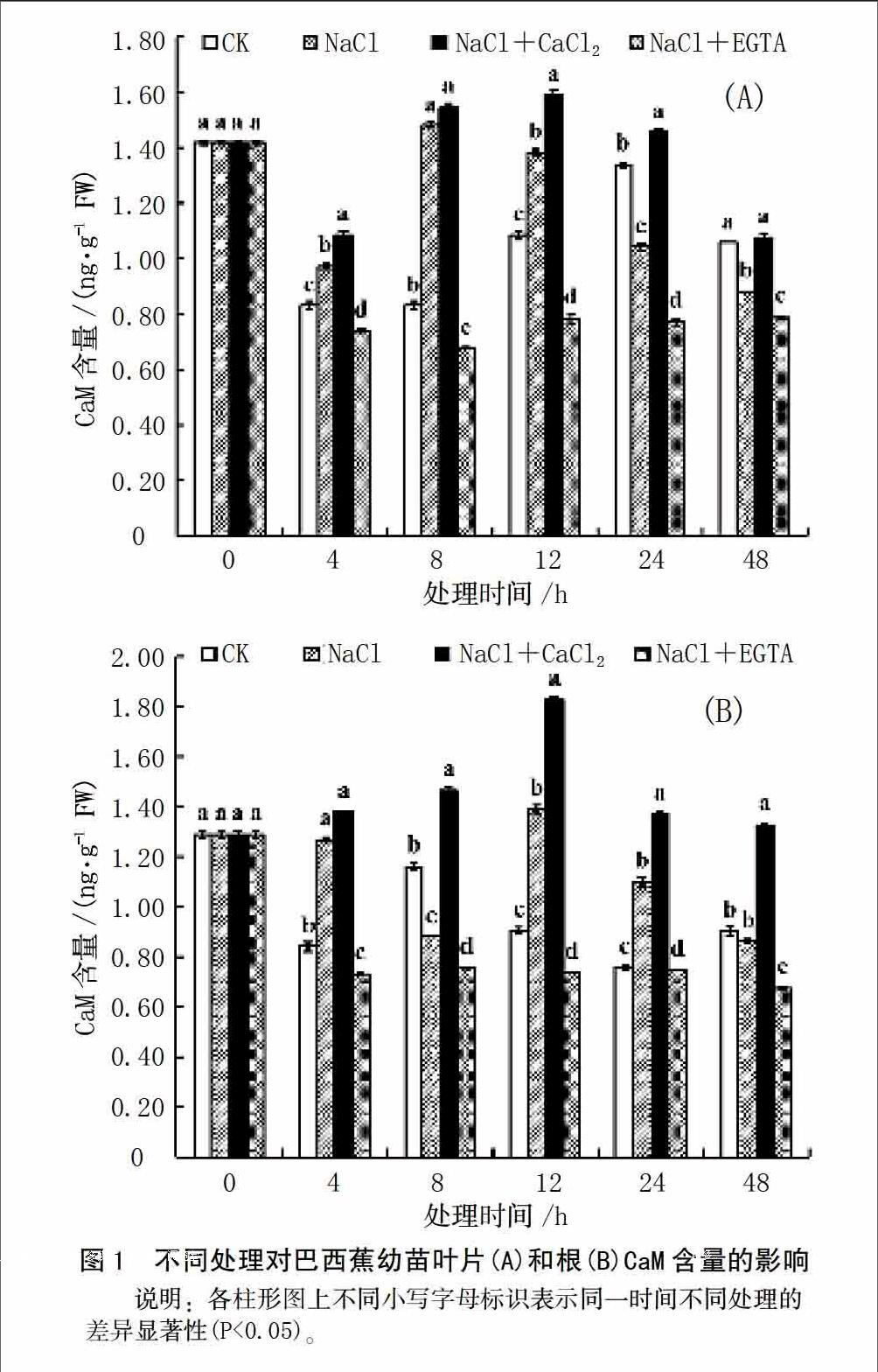
如图1示,与CK处理相比,巴西蕉幼苗叶片在60 mmol/L的NaCl胁迫下,CaM含量在4、8、和12 h处理显著增加(P<0.05),其它时间处理均显著降低;在NaCl胁迫中添加CaCl2处理,除0 和48 h无显著变化外,其它时间处理显著增加(P<0.05);在NaCl胁迫中添加EGTA处理,不同时间处理CaM含量均显著降低(P<0.05)。与NaCl胁迫相比,在NaCl胁迫中添加CaCl2处理,除0和8 h处理无显著变化外,其它时间处理均显著增加(P<0.05);在NaCl胁迫中添加EGTA处理,不同时间处理均显著降低(P<0.05)。CK处理、NaCl处理和NaCl胁迫中添加CaCl2处理的变化趋势一样,均呈先下降再上升最后再下降的趋势,只是下降和上升的幅度和时间表现不同;NaCl胁迫中添加EGTA处理,CaM含量呈下降趋势。CK处理在4 h时,CaM含量显著降低(P<0.05),4和8 h时无显著变化,在12 和24 h显著增加,24 h达到峰值,在48 h又显著降低;NaCl处理4 h时显著降低,8 h又显著增加,在12~48 h则显著降低;NaCl胁迫中添加CaCl2处理在4 h显著降低,在8 h显著增加,在12 h变化不明显,在24和48 h显著降低。
巴西蕉幼苗根的CaM含量,与CK处理相比,在NaCl胁迫下,0 h和48 h未发生显著变化,4、12和24 h显著增加,8 h显著降低;在NaCl胁迫中添加CaCl2处理,不同时间处理均显著增加(P<0.05);在NaCl胁迫中添加EGTA处理,不同时间处理均显著降低(P<0.05)。与NaCl胁迫相比,NaCl胁迫中添加CaCl2处理,除处理0和4 h无显著变化外,其它时间处理均显著增加;NaCl胁迫中添加EGTA处理,不同时间均呈现显著下降(P<0.05)。CK处理呈现双波谷的趋势,NaCl处理呈现先下降再上升最后再下降的趋势,NaCl胁迫中添加EGTA处理呈现先上升再下降的趋势,NaCl胁迫中添加EGTA处理呈现下降的趋势。CK处理在4 h显著降低,在8 h显著增加,12和24 h显著降低,48又显著增加;NaCl处理在4 h无显著变化,在8 h显著降低,在12 h显著增加,在24 和48 h则显著降低;在NaCl胁迫中添加CaCl2处理,在4 和8 h微显著上升,在12 h显著增加(P<0.05),在24 和48 h显著下降;在NaCl胁迫中添加EGTA处理在4 h显著降低,在4~48 h未发生显著变化。
3 讨论
植物在受到非生物刺激诸如盐胁迫时,会普遍产生Ca2+信号,此时,钙调蛋白(CaM)浓度会发生变化,作为靶酶本身不起作用与钙相结合,进而来调节逆境胁迫[10-12]。近年来,在植物逆境胁迫方面,CaM的作用在很多研究领域得到广泛关注[13-14]。当植物遭受盐胁迫时,植物细胞质 Ca2+浓度增加[15]。孙静文[16]指出,苹果补钙可以提高细胞内CaM含量;吴锦程[17]研究发现,钙处理显著提高了低温胁迫后枇杷幼苗叶片细胞CaM含量;林善枝[18]指出,添加CaCl2处理的低温锻炼幼苗,其CaM含量有较大幅度增加;本研究也发现,巴西蕉幼苗叶片的CaM含量,在鹽胁迫除了8 h之外均显著增加,而根的CaM含量在盐胁迫8 h之后才均显著增加。这说明在NaCl胁迫当中,添加CaCl2处理会增加CaM的含量。在显著增加当中,根的CaM含量相对于叶的CaM含量来得多,说明在受到盐胁迫时,巴西蕉幼苗根对逆境的调节比叶片来得高些。
EGTA 处理降低可溶性Ca 和CaM的含量[19],而罗充[20]指出,EGTA处理降低植株Ca2+的含量,但对CaM含量变动规律性不大。加钙螯合剂能明显抑制柑橘原生质体杭冻性的表达,其中,CaM调节原生质体的抗冻性[21]。EGTA处理在干旱胁迫下杨树幼苗叶片的CaM含量明显增加[22]。干旱胁迫在一定程度上提高了CaM含量,而且添加外源钙后更有利于钙信号的传递,但添加各种抑制剂后,CaM含量降低[23];本研究发现,在NaCl胁迫下添加EGTA处理,CaM含量显著降低。
参考文献
[1] 毛国红,宋林霞,孙大业. 植物钙调素结合蛋白研究进展[J]. 植物生理与分子生物学报,2004,30(5):481-488.
[2] Luan S, Kudla J, Rodriguez-Concepcion M, et al. Calmodulins and calcineurin B-like proteins: calcium sensors for specific signal response coupling in plants. Plant Cell, 2002, 14(1): S389-S400.
[3] Mahajan S, Tuteja N. Calcium signaling network in plants[J]. Plant Signaling & Behavior, 2007, 2 (2): 79-85.
[4] 宗 会,李明启. 钙信使在植物适应非生物逆境中的作用[J]. 植物生理学通讯,2001,37(4):330-335.
[5] 杜俊变,段江燕. 逆境胁迫下植物细胞中Ca2+作用的研究进展[J]. 农业与技术,2010,30(2):40-44.
[6] 刘 维. 番茄钙调蛋白和类钙调蛋白的抗病调控功能分析[D]. 浙江大学,2015.
[7] 周双云,蒋 晶,高龙燕,等. 钙对盐胁迫下巴西蕉幼苗膜脂过氧化和抗氧化酶的影响[J]. 应用与环境生物学报,2014,20(30):449-454.
[8] 周双云,蒋 晶,高龙燕,等. 钙对盐胁迫下巴西蕉幼苗膜脂过氧化和抗氧化酶的影响[J]. 西南农业学报,2014,24(6):2 354-2 359.
[9] 罗 充,马湘涛,彭抒昂. 果树生理研究中CaM的ELISA测定. 果树科学,1998(2): 180-185.
[10] 段江燕,张俊红. 植物细胞中钙调素研究进展[J]. 农业与技术,2009,29(6):35-39.
[11] 张君诚,孟玉环,宋育红,等. 植物Ca2+-CaM信号系统及其调控研究进展[J]. 重庆师范大学学报 (自然科学版),2005,22(4):49-52.
[12] Koo S C, Choi M S, Chun H J. The calmodulin-binding transcription factor OsCBT suppresses defense responses to pathogens in rice[J]. Mol Cells, 2009, 27(5): 563-570.
[13] Reddy A S N. Calcium: silver bullet in signaling[J]. Plant Science, 2001, 160(3): 381-404.
[14] Zhou RG, Li B, Liu H, et al. Progress in the participation of Ca2+-calmodulin in heat shock signal transduction[J]. Progress in Natural Science, 2009, 19(10): 1 201-1 208.
[15] Chinnusamy V, Schumaker K, Zhu J K. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signaling in plants. J Exp Bot, 2004, 55(395): 225-36.
[16] 孙静文,周 卫,梁国庆,等. 钙对苹果果实钙调蛋白含量和Ca2+-ATPase活性及其基因表达的影响[J]. 植物营养与肥料学报,2011,17(2):425-432.
[17] 林善枝,张志毅,林元震,等. 钙-钙调素在零下低温诱导毛白杨扦插苗抗冻性中的作用[J]. 植物生理与分子生物学学报,2004,30(1):59-68.
[18] 吴锦程,陈 宇,吴毕莎,等. 钙处理对低温胁迫下枇杷幼苗Ca2+-ATPase活性和膜脂过氧化水平的影响[J]. 西北农林科技大学学报(自然科学版),2016,44(2):121-128.
[19] 易籽林,李志英,何铁光,等. 4种钙素调节剂对乙烯诱导的紫花擎天凤梨花芽分化中钙和钙调素含量的影响[J]. 热带作物学报,2011,32(4):698-701.
[20] 罗 充,彭抒昂,马湘涛. 不同处理对草莓成花过程中钙、钙调素及同化物的含量的影响[J]. 贵州科学,2007,25(S1):470-475.
[21] 李 卫,孙中海,章文才,等. 钙与钙调素对柑橘原生质体抗冻性的影响[J]. 植物生理学报,1997,23(3):262-266
[22] 徐晓辉,刘延吉,王 淼,等. 干旱胁迫对杨树幼苗叶片内ABA和CaM含量的影响[J]. 安徽农业科学,2006(11):2 332-2 335.
[23] 卢 杨. 干旱胁迫下毛竹幼苗钙信号特征及其作用分析[D]. 浙江農林大学,2014.
① 基金项目:国家自然科学基金(No.31260462);海南大学中西部计划学科建设项目(No.ZXBJH-XK008)。
收稿日期:2016-08-19;责任编辑/白 净;编辑部E-mail: rdnk@163.com。
② 王文昌(1989~),男,硕士,主要研究方向为园艺植物栽培与生理,E-mail: 297509593@qq.com。
③ 通讯作者。
