拔节期杂交中稻对淹水胁迫的响应及指示性指标探讨
2017-03-22晏军吴启侠朱建强徐笑笑张露萍
晏军 吴启侠 朱建强* 徐笑笑 张露萍
(1湖北省主要粮食作物产业化协同创新中心,湖北荆州434025;2长江大学农学院,湖北荆州434025;第一作者:yj111708@sina.com;*通讯作者:zyjb@sina.com)
拔节期杂交中稻对淹水胁迫的响应及指示性指标探讨
晏军1,2吴启侠1,2朱建强1,2*徐笑笑1张露萍1
(1湖北省主要粮食作物产业化协同创新中心,湖北荆州434025;2长江大学农学院,湖北荆州434025;第一作者:yj111708@sina.com;*通讯作者:zyjb@sina.com)
长江中下游地区夏季强降水频发,杂交中稻在拔节期易受涝害影响。为定量揭示淹水胁迫对水稻的影响和科学评估灾损,提供形态学、生理特性和产量结构的数据基础,同时筛选拔节期水稻淹水胁迫后受涝及减产程度的指示性指标,在丰两优香1号拔节期设计了不同淹深(1/4PH、1/2PH、3/4PH和全淹)及不同淹水持续时间(3 d、6 d和9 d)交互试验。结果表明,拔节期杂交中稻受淹后,株高、节间显著伸长,且第2节间伸长速度快于第1节,植株黄叶数显著增多,最长叶片长度显著增长,且株高增长量(YPH)、节间长度(YIL)、黄叶数(YYL)、最长叶片长度(YLL)均与淹涝天数(D)和淹深(H)呈显著的二元一次关系;同一淹水持续时间处理,随着淹水深度的增加,水稻叶片中叶绿素a、叶绿素b含量和可溶性蛋白含量逐渐降低,而可溶性糖含量呈先增高后降低的趋势;剑叶中MDA含量均随淹水历时和淹水深度的增加而升高,在根系中,当淹水持续时间为3 d和6 d时,其MDA含量随淹水深度的增加而升高,但在淹水持续时间为9 d时,表现为先升高后降低,其拐点在1/2 PH淹水深度;同时,在同一淹水持续时间下,随着淹水深度的增加,叶片和根系中SOD活性先升高后降低,而剑叶中POD活性持续升高,且水稻受淹越深,其增加幅度愈大,但在根系中的变化趋势是先升高后降低,拐点出现在淹水第6 d的1/2 PH淹深;拔节期水稻受到不同程度涝害后,结实率和千粒重显著下降,当淹水深度达到株高的1/4且持续6 d,其结实率降低20%以上,产量减少19%以上。灰色关联度和聚类分析表明,叶绿素b、可溶性蛋白和叶绿素a可作为拔节期水稻受涝后的关键性生理指标,用于指示、监测水稻受涝及减产程度。
杂交中稻;淹水胁迫;形态结构;生理特性;指示性指标
洪涝灾害是我国主要的农业灾害之一,降水变化影响着我国粮食产量的变化[1]。近年来,全球异常天气增多,局部地区暴雨频发,致使低洼的农田、林地积水并发生涝害,对该地区的农林生产、生态效益及环境质量等方面都造成了潜在的威胁。长江中下游是我国主要的粮食生产区,每年6-8月份为暴雨洪涝的多发期,此时正值早稻生长后期、中稻生长中期、晚稻生长前期,对水稻生产有着重要影响。有关淹水胁迫对水稻生产的影响国内外进行了大量研究,宁金花等[2-6]研究发现,淹水胁迫会引起水稻发生一系列形态和生理生化特征变化,如叶片绿叶数减少、叶鞘伸长、细胞膜脂过氧化作用加剧、丙二醛含量升高、体内保护酶受损、光合速率降低、蒸腾作用减弱、籽粒结实率降低、充实度变差、产量下降等。国外在深水稻适应淹水胁迫机理方面及耐淹基因上研究较多。Raskin等[7-9]分析了深水稻耐淹涝胁迫的生理基础及适应机制,Fukao等[10]研究了水稻的耐淹基因。但这些研究主要集中在早稻、深水稻和常规水稻品种上,淹水的时期多在水稻生长的敏感时期(孕穗期、抽穗扬花期等)和生育后期,且注重淹水胁迫对水稻的影响及适应机制方面,对于拔节期杂交中稻受涝后的系统分析及受涝和减产程度的指示性指标的筛选鲜见报道。
本试验从水稻形态因子、生理特性和产量因子入手,探讨了拔节期杂交中稻受到不同程度淹水胁迫后株高、节间长度、黄叶数、最长叶片长度、生理指标的变化特点以及产量构成因素的变化,并在此基础上以相对产量为参数,筛选拔节期淹水胁迫后受涝及减产程度的指示性指标。旨在明确我国长江中下游地区杂交中稻在适应不同淹水胁迫环境下形态和生理特性变化特点,以及不同淹水环境下水稻的减产机制,为定量揭示淹水胁迫对水稻的影响、洪涝灾害致灾能力及强度的鉴定提供形态学、生理特性和产量结构的数据基础,同时筛选拔节期淹水胁迫后受涝及减产程度的指示性指标。
1 材料与方法
1.1 试验材料
供试品种为丰两优香1号,属中熟中籼两系杂交稻,在湖北省作一季中稻栽培,一般4月下旬至5月上旬播种,5月下旬至6月上旬移栽,8月上旬始穗,9月上中旬成熟,全生育期133 d左右[11],株高约115 cm。该品种拔节期正值江汉平原降水集中期,易遭受淹水胁迫[12]。2014年5月3号播种,6月4日移栽,试验采用盆栽,盆高28~30 cm,内径25.5 cm,每盆装风干土13 kg,每盆2丛,每丛1株。土壤pH值7.6,土壤碱解氮69.4mg/kg、速效磷28.7 mg/kg、速效钾118.7 mg/kg。每盆施肥量为N 2.50 g,K2O 1.95 g,P2O51.13 g。其中,氮肥70%作为基肥、30%作为分蘖肥,钾肥、磷肥全部基施。
1.2 试验设计
在水稻拔节期(第2节间刚长出时、7月13日)进行淹水试验。试验设淹水深度(H)和淹水时间(D)2个因素。淹水深度设4个水平:1/4淹(淹水深度为当时水稻株高的25%,1/4 PH)、1/2淹(1/2 PH)、3/4淹(3/ 4PH)、没顶淹(4/4 PH)。淹水时间设3个水平:3 d、6 d和9 d,每个处理6次重复。以整个生育期正常水分管理为对照(CK),设15个重复。淹水试验在1.5m深的水泥池中进行,池中水为静止、洁净的井水,9 d内不换水。试验期间根据水稻长势随时调整淹水深度,保证试验准确性。每个淹水时间到后取出6盆,其中3盆用于形态指标和生理指标的测定,另外3盆置于接近盆高的水池中继续生长,水池中灌满水,尽量消除温度等环境因素对盆栽水稻的影响。除淹水外,水稻其他管理同大田。
1.3 测定指标
1.3.1 形态指标
分别于试验开始时和结束时测定水稻株高、第1、2节间长度、黄叶数与每丛中最长叶片长度(最长叶片用笔做好标记)。每盆2丛水稻平均值作为1个重复值,每个处理测定3盆,为每个处理的3个重复值。在淹水前和取出后对每盆中所有茎上的黄叶数进行统计,按照农业气象相关规范,能进行光合作用的叶片均为绿叶,部分黄化的叶片则按黄色部分占整个叶片的百分比进行计算,如黄色部分面积占整个叶片面积的50%,其余部分是绿色,则记为0.5片黄叶,为目测估算值,结果保留1位小数。在淹水前和淹水结束后对每丛做记号的叶片进行测量。同时,对照盆中的株高、第1、2节间长度、黄叶数与每丛中最长叶片长度进行平行观测、统计。
1.3.2 生理指标
于试验结束时测定对照和处理水稻剑叶和根系的叶绿素、可溶性糖、可溶性蛋白、丙二醛(MDA)含量及抗氧化酶(SOD、POD)活性。采用浸提法(95%乙醇)测定剑叶的叶绿素a、b的含量;蒽酮比色法测定可溶性糖含量;考马斯亮蓝G-250法测定可溶性蛋白含量;硫代巴比妥酸(TBA)比色测定丙二醛(MDA)含量;氮蓝四唑(NBT)法测定超氧化物歧化酶(SOD)活性,以抑制NBT光化还原50%为1个酶活性单位(U)表示;愈创木酚法测定过氧化物酶(POD)活性,POD活性以470 nm处吸光度每分钟变化0.01为1个酶活性单位(U)表示。每个处理重复测3次,取平均值[13]。
1.3.3 产量
水稻成熟后,单独统计每盆水稻有效穗数(大于等于5粒实粒的穗子),取平均值作为该盆的有效穗数,然后将2丛一起收获,随机取10穗,分别统计各穗的实粒数、秕粒数和空粒数,并计算整穗的结实率,然后统计每个处理的千粒重(g),每个处理水稻脱粒后晒干除杂称重,作为每个处理的实际产量(g)。
1.4 统计分析
通过盆栽试验,依据拔节期杂交中稻对不同程度淹水胁迫的响应,求得淹水持续天数(D)、淹水深度(H)与水稻株高增长量(YPH)、节间伸长量(YIL)、黄叶数(YYL)及最长叶片长度(YLL)之间的关系:

式1中,Y指代YPH、YIL、YYL、YLL;式(2)中,PH2为淹水结束时各处理株高(cm),PH1为淹水开始时各处理株高(cm)。
根据灰色系统理论,首先对各指标值进行无量纲化处理。相对产量与15个形态、生理性状看作1个灰色系统,各指标看作是该系统的1个灰因素,其中相对产量设为参考数列,15个形态、生理性状设为比较数列。应用DPS15.10软件进行方差、回归分析、灰色关联度分析和聚类分析,利用Microsoft Excel 2003作图,采用LSD法进行处理间多重比较。
2 结果与分析
2.1 淹水胁迫对水稻形态结构的影响
2.1.1 淹水胁迫对水稻株高和节间增长量的影响
图1表明,淹水天数分别为3 d、6 d和9 d时,淹水深度每增加1/4 PH时,水稻株高分别增长了1.3 cm、3.5 cm和4.7 cm;在同一淹涝深度下,淹涝天数每增加3 d,第1节间伸长值为0.9~1.5 cm(对照0.8 cm),第2节间伸长值为3.0~6.6 cm(对照1.2 cm),第2节间伸长速度快于第1节间。表明同一淹水持续天数下,随着淹水深度的增加,水稻株高的增长速度与淹水深度呈正比;在同一淹水深度下,节间伸长度与淹水持续天数呈正比。LSD检验表明,淹水3 d、6 d和9 d时,淹深分别达到1/2PH、1/4PH和1/4PH处理的株高显著高于对照(P<0.05)。
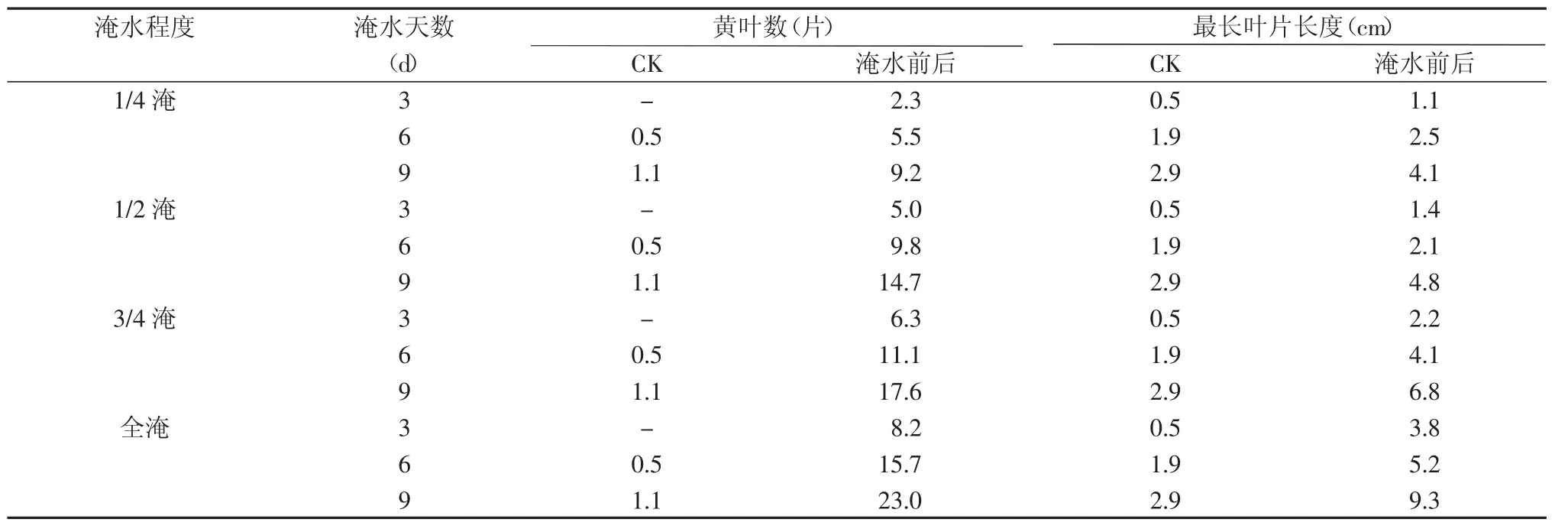
表1 不同淹水处理后黄叶数与最长叶片长度变化

图1 不同淹水处理后株高和节间长变化
2.1.2 淹水胁迫对杂交中稻黄叶数与最长叶片长度的影响
从表1可见,与对照相比,淹水处理下的水稻植株黄叶数变化均非常明显。在同一淹水深度下,拔节期植株黄叶数和最长叶片长度的变化趋势与淹水天数呈正比,淹水天数每增加3 d,水稻的黄叶数与对照相比增加了2.3~21.9片,最长叶片长度增加了0.6~6.4 cm。即同一淹水深度,淹水时间越长,绿叶衰老死亡越快。黄叶数及叶片长度的变化除了与淹涝环境有关外,还与水稻植株所处的生育期有很大的关系。淹涝胁迫是外因,水稻本身的生理因素是内因。不同淹水深度的淹涝胁迫引起黄叶数和叶片长度的变化是内外因综合的表现。
2.1.3 淹水胁迫下水稻形态结构与D、H的量化关系
对图1和表1试验结果作回归分析,结果见表2。从表2可见,拔节期淹水胁迫处理结束时水稻株高增长量、节间伸长量、黄叶数及最长叶片长度与淹水天数和淹水深度之间呈极显著的二元一次关系,且回归关系式的F值达极显著水平(sig.<0.01)。因此,可以用这些回归关系式定量化诠释淹水天数和淹水深度对水稻株高增长量、节间伸长量、黄叶数及最长叶片长度的综合影响。从标准化回归系数的绝对值可以看出,淹水深度对株高伸长量、第2节间长度和黄叶数的影响要大于淹水持续天数,淹水持续天数对第1节间长度和最长叶片长度的影响要大于淹涝深度。
2.3 淹水胁迫对杂交中稻叶片中叶绿素含量的影响
从外观上看,水稻在拔节期受淹水胁迫后叶片最显著的响应是失绿黄化,其原因是叶绿素水解加快、叶黄素合成增加。试验所测叶绿素各组分含量见图2。受淹水胁迫处理叶片中的Chl a、Chl b含量均比受涝前低,且受涝时间越长其含量越低。LSD检验表明,在淹水时间为3 d、6 d和9 d时,淹水深度达1/4 PH以上时,Chl a、Chl b含量均显著低于对照(P<0.05),表明拔节期水稻在受到轻微淹水胁迫后叶绿素出现快速水解,即叶绿素对淹水胁迫极为敏感。

表2 淹水处理后杂交中稻YPH、YIL、YYL、YLL分别与D、H的量化关系

图2 不同淹水处理后剑叶中叶绿素含量变化

图3 不同淹水处理后剑叶和根系中MDA含量变化
2.4 淹水胁迫对杂交中稻膜脂过氧化作用的影响
MDA作为脂质过氧化程度的指标,是脂质过氧化的一种典型产物,它能与蛋白质的氨基或核酸反应生成shiff碱,MDA的积累会对膜和细胞造成进一步的伤害,进而引起一系列生理生化变化[14]。图3给出了淹水胁迫对水稻剑叶和根系MDA含量的影响。剑叶中的MDA含量均随淹水历时和淹水深度的增加而升高;但在根系中,当淹水持续时间为3 d和6 d时,其MDA含量随淹水深度的增加而升高,但在淹水持续时间为9 d时,表现为先升高后降低,其拐点在1/2 PH淹水深度。LSD检验表明,除淹水持续时间为3 d、淹水深度为1/4 PH和淹水时间为9 d、淹水深度为3/4 PH和4/4 PH,其余处理剑叶和根系中MDA含量均显著高于CK(P<0.05),表明杂交中稻受涝时间达到3 d、淹水深度1/4 PH左右时,其剑叶和根系细胞膜脂就遭受严重过氧化。根系中淹水第9 d、1/2 PH淹深的处理MDA含量最高,继续淹水MDA含量呈下降趋势,这可能与根系器官严重受损有关。
2.5 淹水胁迫对杂交中稻可溶性蛋白与可溶性糖含量的影响
可溶性蛋白作为重要的渗透调节物质,可以起到调节细胞渗透压,保护膜结构稳定的功能,糖类是新陈代谢的基础,植物体内糖含量越高,则新陈代谢越旺盛,其生命力就越强[15]。图4表明,在同一淹涝持续天数下,随着淹水深度的增加,受淹水稻叶片中的可溶性蛋白含量逐渐降低,而可溶性糖含量呈先增高后降低的趋势。LSD检验表明,当淹水持续时间分别达到3 d、6 d和9 d,在淹水深度分别为1/2 PH、1/4 PH和1/4 PH以上时,水稻叶片中的可溶性蛋白含量显著低于CK(P<0.05),而可溶性糖含量在淹水深度分别为1/2 PH、1/4 PH和1/2 PH、1/4 PH时,显著高于CK(P<0.05),但是在第6 d和第9 d全淹时显著低于CK(P<0.05)。
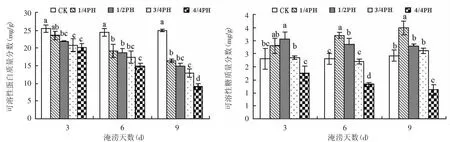
图4 不同淹水处理后剑叶中可溶性蛋白与可溶性糖含量变化

图5 不同淹水处理后剑叶和根系中SOD、POD含量变化
2.6 淹水胁迫对杂交中稻抗氧化酶活性的影响
淹水胁迫会影响植物体内活性氧代谢系统的平衡,一方面超氧化物阴离子自由基、羟基自由基、单线态氧和过氧化物等大量产生,抑制保护性酶的活性;另一方面SOD、POD等酶组成了防御生物活性氧毒害的酶保护系统,在植物体内行使清除活性氧的功能。图5表明,水稻在受到不同程度淹水胁迫后,叶片和根系中SOD活性的变化趋势是先升高后降低;POD活性在叶片中的变化趋势是持续升高,且淹水时间越长,其增加幅度愈大,而在根系中的变化趋势是先升高后降低,拐点出现在淹水第6 d和第9 d的1/2 PH淹深。LSD检验表明,处理间差异显著。表明杂交中稻在淹水3 d后其体内的POD、SOD活性就发生显著改变,并且活性会随着淹水深度和淹水时间的增加而显著增加,起到有效清除活性氧自由基的功能,减轻活性氧自由基对细胞膜的伤害。但是,当淹水胁迫达到一定程度,POD、SOD的活性就会呈下降趋势,这可能是由于无氧呼吸产生的有害物质逐渐增多,从而影响了保护酶的产生,最终使得作物生长受到伤害。
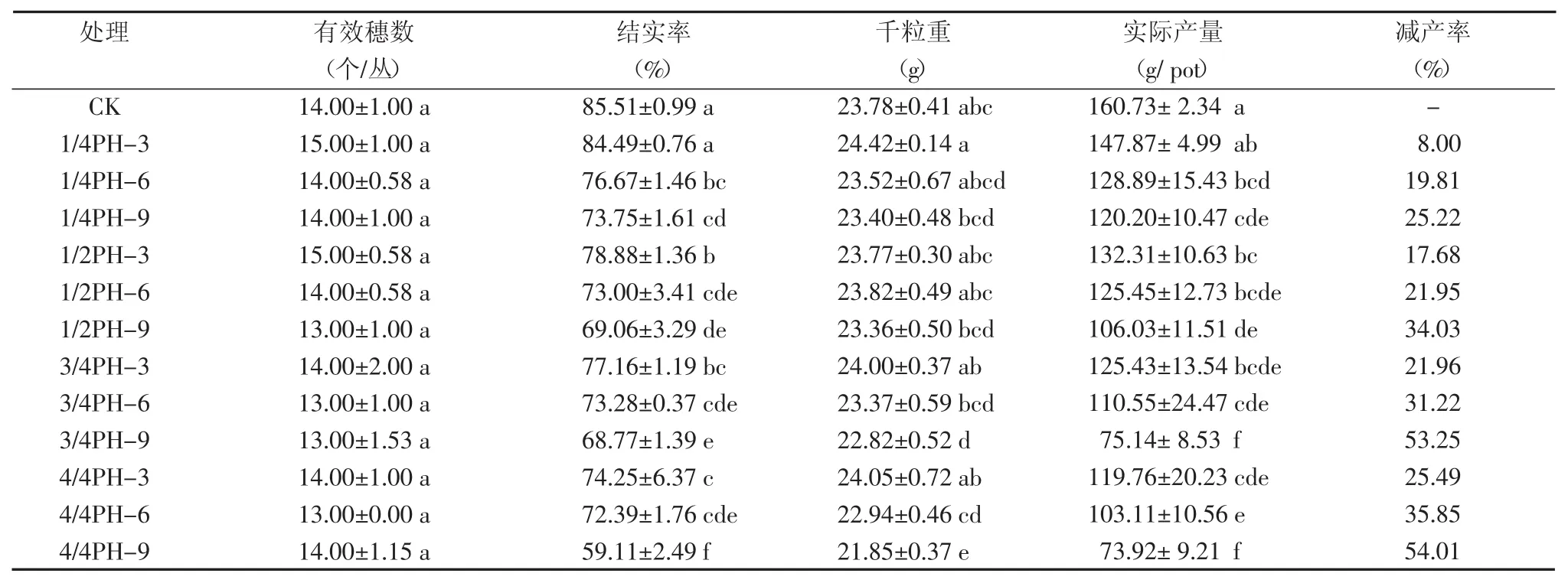
表3 拔节期不同淹水处理后杂交中稻产量结构

表4 淹水持续时间、深度与实际产量及产量构成因素的相关性
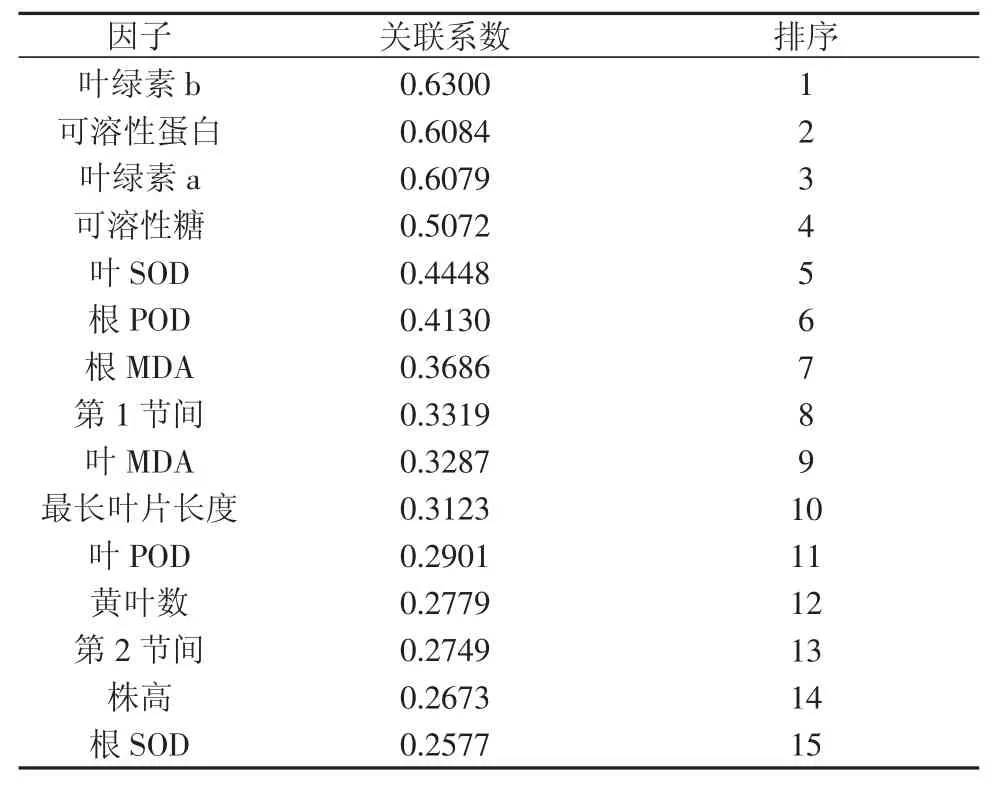
表5 相对产量与各形态、生理指标的灰色关联度分析
2.7 淹水胁迫对水稻产量要素的影响
如表3所示,与对照相比,4个淹水处理水稻结实率、千粒重、实际产量降低,而且淹水深度越深、淹水持续时间越长,结实率、千粒重、实际产量越低。LSD检验表明,水稻拔节期遭受不同程度淹水胁迫后,有效穗数与对照相比差异不显著(P>0.05),结实率除1/4 PH淹深持续3 d的处理外,其余处理结实率与对照差异显著(P<0.05);千粒重除3/4 PH淹深持续9 d、4/4 PH淹深持续9 d两个处理外,其余处理千粒重与对照差异不显著;而实际产量除1/4 PH淹深持续3 d的处理外,其余处理实际产量与对照差异显著(P<0.05)。表4表明,4个淹水深度处理下,有效穗数、结实率、千粒重、实际产量与淹水时间呈显著或极显著负相关;在3个淹水持续时间下,结实率和实际产量与淹水深度呈显著负相关,有效穗数和千粒重与淹水深度相关性不显著。总结得出,水稻拔节期受到不同程度淹水后,主要导致结实率和实际产量降低,当淹水深度达到株高的1/4并持续淹水6 d,其结实率显著降低20%以上,实际产量减少19%以上,可能是由于生育期推迟,花粉发育受阻,甚至花药坏死,生长期延长和籽粒灌浆受阻等多种原因所致。
2.8 水稻受淹及减产程度的指示指标探讨
根据灰色系统理论,可将相对产量和相关形态、生理生化性状看作一个灰色系统,各性状作为该系统的一个灰因素,其中相对产量设为参考数列,15个逆境胁迫生理指标设为比较数列,通过灰色关联度分析法确定叶绿素a、b含量等13个指标与拔节期水稻受淹后相对产量的关联度。关联度值越大,两数列的变化态势越接近,相互关系越密切。由表5可以看出,在同一淹水程度下,不同指标与相对产量的关联度不同。根据关联度系数大小进行排序可见,叶绿素b、可溶性蛋白、叶绿色a等指标在水稻受淹水胁迫后关联度排在前3位,说明其与植株相对产量的相关性最高,可作为拔节期水稻受淹后的指示指标,初步确定其受淹及减产程度。
依据各抗逆相关形态、生理指标关联度系数进行聚类分析,结果见图6。由聚类分析结果可以看出,各性状可分为三大类。第一类包括生理指标叶绿素b、可溶性蛋白和叶绿素a,它们与相对产量的关联度最大,稳定性高;第二类包括生理指标可溶性糖、叶SOD和根POD,它们与相对产量的关联程度较为密切,稳定性较低;第三类主要为根MDA、第一节间长度和叶MDA等生理、形态指标,这些指标关联度最小,稳定性最差。因此第一类指标(叶绿素b、可溶性蛋白和叶绿素a)可作为关键性生理指标用于指示、监测水稻受淹及减产程度。

图6 关联系数的聚类分析
3 讨论与结论
3.1 水稻形态结构与淹水胁迫
水淹植物对光的需求与其自身的形态紧密相关[16],随着光强的减弱,会产生响应,如叶片尺寸[17]和株高[18]发生改变。前人对淹水胁迫后的水稻节间伸长[19]的现象进行了研究,主要针对浮稻,当被水淹没时,耐淹品种往往是以茎和/或叶柄的伸长使植物从水中露出,从而恢复与大气的接触,空气由叶片气孔或茎部皮孔进入,沿着通气组织输送到低氧根部,缓解了低氧危害。淹水胁迫可以诱导植物叶柄和茎的伸长生长[18,20],研究对象主要是深水稻,对于深水中的不耐涝的水稻,在水下的快速生长被认为可以使许多植物提高生存机会[21],在长期淹水条件下的茎秆伸长可以使嫩枝达到水面以上,有助于保证充足的氧气和二氧化碳,使超过水位的叶子能够进行光合作用和确保获取有效碳[22]。淹水使节间和株高发生变化的原因比较复杂,生理学研究表明,植物乙烯、脱落酸(ABA)和赤霉素(GA)参与调节淹水胁迫下水稻伸长生长反应[8],GA是与淹水胁迫下水稻的伸长生长反应直接相关的激素,它能促进叶和节间伸长生长[7]。本研究表明,对杂交中稻进行淹水处理后,水稻株高、第1、2节间、最长叶片长度表现出增加的趋势,且淹水时间越长、淹水深度越深,增加幅度越大,株高增长量、节间长度、最长叶片长度均与淹水天数和淹水深度呈显著的二元一次正相关关系,相比较而言,淹水深度对株高伸长量、第2节间长度和黄叶数的影响要大于淹水持续天数,淹水持续天数对第1节间长度和最长叶片长度的影响要大于淹涝深度。
3.2 水稻生理特性与淹水胁迫
淹水对植物的胁迫作用最主要的是导致缺氧,从而诱发一系列的生理方面的变化,这种变化是植物对逆境的反应或适应。相关研究[23-26]表明,水稻在淹水条件下绿叶数、叶绿素含量、光合速率降低,除泥沙附在叶片上堵塞气孔外,与叶片中叶绿素的分解增加、MDA积累以及在淹水条件下水稻植株体内高乙烯与低CO2的相互作用有关。从本试验结果来看,随着淹水时间和淹水深度的增加,Chl a、Chl b逐渐降低,MDA在叶片和根系中的含量均逐渐升高。Richard等[27]发现,水稻淹水60 min内,葡萄糖-6-磷酸脱氢酶(G-6-PDH)基因增加了转录,并在以后的141 h内活性一直增高,这反映了磷酸戊糖途径活性的增强。本研究发现,叶片中可溶性糖含量随着淹水持续时间和深度的增加,呈现先增高后降低的趋势,也验证了该结论。李萍萍等[28-29]研究发现,高温、干旱等胁迫均导致水稻可溶性蛋白含量不同程度降低。本试验结果表明,拔节期淹水胁迫导致水稻叶片中可溶性蛋白含量显著降低。Chang等[30]指出,低氧适应反应不是某种蛋白合成增加或减少的结果,也不只是低氧诱导蛋白的作用,而是多种基因产物共同作用的结果。
淹水胁迫导致植物生长环境发生改变,超氧化物歧化酶(SOD)和过氧化物酶(POD)是作物内在的保护酶系统,它们既有分工又有合作,在作物抗逆代谢中起着重要的保护作用。SOD活性的增加对植株有效抵御厌氧胁迫非常关键,它是过氧化物O2-的主要清除剂,能将O2-转化成H2O2和O2,而POD能催化H2O2释出新生氧以氧化某些酚类物质和胺类物质[31]。国内相关研究[2-6]主要集中在水稻的叶片或者根系,且没有将2个器官的变化进行比较。本试验表明,SOD的活性随着淹水时间和淹水深度的增加在叶片、根系中表现出先增高后降低的趋势,POD的活性随着淹水时间和淹水深度的增加在叶片中表现出逐渐增高的趋势,在根系中表现出先增高后降低的趋势,说明水稻受到淹水胁迫达到一定程度,叶片和根系的保护酶系统就会受到破坏,只是叶片相对于根系来说可能会相对滞后一点,原因是根系对于周围环境变化相对更加敏感。
3.3 淹水胁迫对水稻产量性状的影响及指示性指标探讨
不同生育期水稻受到淹水时各产量构成要素的反应不一样。宁金花等[3]研究发现,拔节期早稻受淹涝对产量的影响均表现为空秕粒率增加、千粒重和实粒数降低;孙系巍等[32]研究发现,乳熟期淹涝胁迫对水稻产量的影响主要表现为空壳率、秕粒率升高,千粒重下降;张艳贵等[33]认为,分蘖期水稻受淹水胁迫后产量结构表现为穗小、实粒数少、千粒重低;而宣守丽等[34]认为,水稻单丛穗数减少、单穗产量下降是分蘖期淹水后产量下降的主要原因。本试验结果表明,拔节期杂交中稻受到不同程度淹水后,主要导致结实率和实际产量降低,当淹水深度达到株高的1/4并持续6 d,其结实率显著降低20%以上,实际产量减少19%以上。
评估作物受淹后恢复程度的指标很多,各指标在综合评价时所占权重又各不相同。关联度分析法不仅可以给出质的定性解释,也可以给出量的确切描述,关联度越大,说明两数列的变化态势越接近,相互关系越密切[35]。国内外学者已筛选出许多与作物抗水分胁迫有关的形态及生理生化指标,如叶片水势、气孔导度、蒸腾速率、可溶性糖、脯氨酸、丙二醛、保护酶等,这些指标可作为指示棉花渍水胁迫后胁迫程度或恢复水平的候选生理指标[36-38];在研究小麦耐渍性时发现,净光合速率、脯氨酸、丙二醛、根系活力等综合指标可作为鉴定小麦耐渍性的生理指标[39];在作物抗旱性方面,光合作用特性指标、活性氧代谢指标可用作甘蔗抗旱性生理生化鉴定指标[40]。本研究认为,叶绿素b、可溶性蛋白和叶绿素a可作为拔节期水稻受涝后的关键性生理指标,用于指示、监测水稻受涝及减产程度。但是本试验是采用盆栽试验,试验条件可控,且只进行了1年,研究的影响指标较少,是否还有其他关联性更高的指标(如净光合速率、气孔导度、蒸腾速率等),有待于进一步确定和验证。
[1]潘澜,薛立.植物淹水胁迫的生理学机制研究进展[J].生态学杂志,2012,31(10):2662-267.
[2]宁金花,霍治国,陆魁东,等.不同生育期淹涝胁迫对杂交稻形态特征和产量的影响[J].中国农业气象,2013,34(6):678-684.
[3]宁金花,陆魁东,霍治国,等.拔节期淹涝胁迫对水稻形态和产量构成因素的影响[J].生态学杂志,2014,33(7):1 818-1 825.
[4]蔺万煌,孙福增,彭克勤,等.洪涝胁迫对水稻产量及产量构成因素的影响[J].湖南农业大学学报,1997(1):53-57.
[5]彭克勤,夏石头,李阳生.涝害对早中稻生理特性及产量的影响[J].湖南农业大学学报,2001,27(3):173-176.
[6]李阳生,李绍清.淹涝胁迫对水稻生育后期的生理特性和产量性状的影响[J].武汉植物学研究,2000,18(2):117-122.
[7]Raskin I,Kende H.Role of gibberellin in the growth response of submerged deepwater rice[J].PlantPhys,1984,76(4):947-950.
[8]Kende H,Van D K E,Cho H T.Deep water rice:A model plant to studystem elongation[J].PlantPhys,1998,118(4):1 105-1 110.
[9]Van DSD,Zhou Z,Prinsen E.A comparativemolecular-physiologicalstudy ofsubmergence response in lowland and deep water rice[J]. PlantPhys,2001,125:955-968.
[10]Fukao T,Harris T,Bailey-serres J.Evolutionary analysisof the Sub1 gene cluster thatconferssubmergence tolerance to domesticated rice [J].Ann Bot,2009,103:143-150.
[11]彭斯敏,耿延琢,程勤学,等.丰两优香1号在湖北省种植表现及其高产栽培技术[J].杂交水稻,2009,24(5):52-53.
[12]朱建强.易涝易渍农田排水应用基础研究[M].北京:科学出版社,2007:43-44.
[13]张治安,陈展宇.植物生理学实验指导[M].长春:吉林大学出版社,2008.
[14]Chaoui A,Mazhoudi S,Ghorbal M H.Cadmium and Zinc induction of lipid peroxidation and effectson antioxidanten-zymeactivities in bean[J].PlantSci,1997,127:139-147.
[15]李佐同,靳学慧,张亚玲,等.水稻幼苗可溶性糖及可溶性蛋白含量与抗瘟性的关系[J].北方水稻,2009(4):6-9.
[16]Middelobe A L,Markager S.Depth limits and minimum light requirementsof freshwatermacrophytes[J].Freshwater Biol,1997,137: 553-568.
[17]罗文泊,谢永宏,宋凤斌.洪水条件下湿地植物的生存策略[J].生态学杂志,2007,09:1478-1485.
[18]Jackson M B.Ethylene-promoted elongation:an adaptation to submergencestress[J].Ann Bot,2008,101(2):229-248.
[19]Hirano T,Koshimura N.Growth and distribution of photo assimilates in floating riceunder submergence[J].Jpn JTropical Agri,1995,39(3):177-183.
[20]Naoyoshi Kawano,Osamu Ito,Jun-Ichi Sakagami.Relationship between shootelongation and drymatterweight during submergence in Oryza sativa L.and O.glaberrima Steud.rice cultivars[J].PlantProd Sci,2008,11:316-323.
[21]Jackson M B.Ethylene and responses of plants to soilwaterlogging and submergence[J].Annu Rev PlantPhysiol,1985,36:145-174.
[22]Ito O,Ella E,Kawano N.Physiological basis of submergence tolerance in rainfed lowland rice ecosystem[J].Field Crop Res,1999,64: 75-90.
[23]李乐农.淹水处理对不同水稻品种碳、氮代谢的影响[J].湖南农学院学报,1995,21(3):235-238.
[24]李绍清,李阳生,李达模,等.乳熟期淹水对两系杂交中稻源库特性的影响[J].杂交水稻,2000,15(2):38-40.
[25]Reddy M D.Effect of complete plant submergence on vegetative growth,grain yield and some biochemical change in rice plant[J]. PlantSoil,985,87(3):365-374.
[26]PalachaM C,Vergara BS.Environmentaleffectson the resistanceof rice seedling to complete submergence[J].Crop Sci,1972,12:209-212.
[27]Richard B,Rivoal J,Spiteri A,et al.Anaerobic stress induces the transcription and translation of sucrose synthase in rice[J].Plant Physiol,1991,95:669-674.
[28]李萍萍,程高峰,张佳华,等.高温对水稻抽穗扬花期生理特性的影响[J].江苏大学学报,2010,31(2):1 671-1 675.
[29]王贺正.水稻抗旱性研究及其鉴定指标的筛选[D].成都:四川农业大学,2007.
[30]ChangWW P,Huang L,Shen M,etal.Patternsof protein synthesis and toleranceofanoxia in root tipsofmaize seedlingsacclimated toa low-oxygen environmentand identification of proteinsbymass spectrometry[J].PlantPhysiol,2000,122(2):295-317.
[31]谭淑端,朱明勇,张克荣,等.植物对水淹胁迫的响应与适应[J].生态学杂志,2009(9):1 871-1 877.
[32]孙系巍,宁金花,张艳桂,等.乳熟期淹涝胁迫对水稻形态特性及产量的影响[J].湖南农业科学,2015(6):27-30.
[33]张艳贵,宁金花,谢娜,等.分蘖期淹涝胁迫对水稻形态及产量的影响[J].湖南农业科学,2014(7):14-17.
[34]宣守丽,石春林,张建华,等.分蘖期淹水胁迫对水稻地上部物质分配及产量构成的影响[J].江苏农业学报,2013(6):1 199-1 204.
[35]胡江龙,郭林涛,王友华,等.棉花渍害恢复的生理指示指标探讨[J].中国农业科学,2013,46(21):4 446-4 453.
[36]冯方剑,宋敏,陈全家,等.棉花苗期抗旱相关指标的主成分分析及综合评价[J].新疆农业大学学报,2011,34(3):211-217.
[37]Ennahli S,Earl H J.Physiological limitations to photosynthetic carbon assimilation in cotton underwater stress[J].Crop Sci,2005,45(4):2 374-2 380.
[38]王贺正,马均,李旭毅,等.水稻开花期抗性鉴定指标的筛选[J].作物学报,2005,31(11):1 485-1 489.
[39]周广生,梅方竹,周竹青,等.小麦不同品种耐湿性生理指标综合评价及其预测[J].中国农业科学,2003,36(11):1 378-1 382.
[40]高三基,罗俊,张华,等.甘蔗抗旱性生理生化鉴定指标[J].应用生态学报,2006,17(6):1 051-1 054.
Abctract:In themiddle and lower reaches of the Yangtze River,there ismore rainfall in summer,especially in jointing stage of hybridmid-season rice,so that rice is liable to suffer from waterlogging damage.In order to quantitively reveal the influence of flooding on hybrid mid-season rice growth,scientifically assess the yield loss caused by it,and to provide data base of the morphological, physiological characteristics and yield structure,those indexes presented higher correlation degree with relative yield could be used as keymorphological and physiological indexes to indicate the yield reduction level at the jointing stage of hybrid mid-season rice after flooding,a cross test of flooding depth(a quarter,a half,three-quarters and full submerged)and waterlogging duration(3,6 and 9 days)was done in the jointing stages of hybrid mid-season rice.The results showed that after submergence treatment,the rice height and the internode length all present elongation(the 1rd and 2th internode elongatemainly in the booting stage,the 2rd internode elongating faster),the yellow leaves number and the longest leaf length of plant increased significantly,which is significant linear positive correlation with the increase of flooding depth and waterlogging duration,A significant correlation of two variableswith the first power exists among YPH(plant height growth),YIL(internode length),YYL(yellow leaves number),YLL(longest leaf length)and D(submergence days),H(submergence depth);as far as physiological characteristics,under the identical flooding duration,chlorophyll(Chla and Chlb)and soluble protein content of sword leaf was decreased with the increase of waterlogging duration,which has significant negative correlation with flooding depth,but the contentof soluble saccharide was increased first and then decreased with the increase of flooding depth,while the MDA content in the sword leafwas significantly positively related with the increase of flooding depth,and in the rootwas increased first and then decreased when the flooding duration was 9 days,which the turning pointwas a half submergence depth.At the same time,the activity of SOD in the sword leaf and rootwas increased first and then decreased with the increase of flooding depth,the activity of POD in sword leaf presents the rising trend with the increase of flooding depth,moreover the severe the waterlogged depth was,the higher the amplification of POD activity could be,and that in root was increased first and then decreased with the increase of flooding depth,which the turning pointwas a half of submergence for 6 days,under the identical flooding duration.In lightof the yield component,the seed setting rate reduced bymore than 20%and yield decreased by 19%than thatof the controlwhen the rice run into quarter of submergence for 6 days at the jointing stage.Through gray correlation and cluster analysis in this research,it was concluded that Chlb,soluble protein and Chla content could be used as key physiological indexes in evaluating waterlogged and reduction of yield degree after flooding.
Response of Hybrid M id-season Rice to Flooding and Discussion of Indicative Index at the Jointing Stage
YAN Jun1,2,WU Qixia1,2,ZHU Jianqiang1,2*,XU Xiaoxiao1,ZHANG Luping2
(1Hubei Collaborative Innovation Center for Grain Industry,Jingzhou 434025,China;2College of Agriculture,Yangtze University,Jingzhou 434025, China;1st author:yj111708@sina.com;*Corresponding author:zyjb@sina.com)
hybridmid-season rice;flooding stress;morphological structure;physiological property;indicative index
S963;S511
:A
:1006-8082(2017)01-0017-09
2016-09-29
农业部公益性行业(农业)科研专项(201203032)
