秦岭火地塘林区不同海拔森林土壤NO通量1)
2017-03-13李玉田庞军柱彭长辉张硕新侯琳陈书军王效科张红星
李玉田 庞军柱 彭长辉 张硕新 侯琳 陈书军 王效科 张红星
(陕西秦岭生态系统国家野外科学观测研究站)(中国科学院生态环境研究中心)(西北农林科技大学,杨凌,712100)
秦岭火地塘林区不同海拔森林土壤NO通量1)
李玉田 庞军柱 彭长辉 张硕新 侯琳 陈书军 王效科 张红星
(陕西秦岭生态系统国家野外科学观测研究站)(中国科学院生态环境研究中心)(西北农林科技大学,杨凌,712100)
选取秦岭火地塘林区不同海拔(1 560~2 160 m)的有代表性的5个森林样地,从2014年10月到2015年10月对其土壤NO通量采用静态箱—氮氧化物分析仪法进行1 a的监测。结果表明,土壤NO排放主要集中在植物生长季(2015年5月—2015年9月),但整个观测期NO排放保持在较低水平。在非生长季(2014年10月—2015年4月),大部分样地的NO排放先减少后增加,而且监测有NO吸收。不同海拔NO年排放总量分别为2 160 m红桦林0.06 kg·ha-2·a-1,2 040 m青杄林0.08 kg·ha-2·a-1,1 963 m 华山松林0.02 kg·ha-2·a-1,1 585 m油松林0.11 kg·ha-2·a-1,1 560 m 锐齿栎林0.19 kg·ha-2·a-1。除华山松外,NO年排放总量随海拔的升高而减少。不同海拔5个样地土壤NO通量均与地温显著相关(P<0.05)。华山松林土壤NO排放与土壤孔隙充水率呈负相关(P<0.05)。油松林土壤NO排放与土壤铵态氮质量分数负相关(P<0.05)。除油松林外,各样地的土壤孔隙充水率都低于60%,可以推断,硝化反应是本地区NO的重要生成源,但受到降雨和土壤有机质等理化性质的影响又伴随有反硝化过程。
秦岭;森林;土壤;一氧化氮通量

热带和温带森林土壤被认为是重要的NO排放源[8-12]。然而,当前对土壤NO通量的研究大多集中在欧洲或美洲[11,13-18],有关亚洲的报道很少[12,19]。Luo等[20]指出,为了更好地理解温带森林土壤NO排放过程和季节变化,需要更多监测周期较长(≥1 a)和采样频率较高的研究。Pilegaard[6]指出,为了加深对景观尺度NO通量的理解,很有必要考虑地形条件对土壤环境因子的调控以及进一步对NO排放和吸收的影响。
地形因素可以通过影响土壤的微气候、土壤的理化性质、植被的生长以及地下碳输入等途径影响土壤微生物的生长及生物量[21-22],即使很小的地形变化也会造成短距离内土壤条件的剧烈变化[23]。环境随着海拔变化而变化,海拔是影响光、热、水、气的因子之一,其直接作用于生境的气候生态学特征,并通过对气候环境的改变使土壤发生物理、化学和生物方面的变化,进一步影响植物群落结构和类型的演化。一定的海拔高度就有一定的均温、活动积温和降水量,地形的垂直变化,会显著地影响土壤水热条件[24]。另外,不同林分下地表植被和地表枯落物不同,导致土壤养分的归还量也不同,影响硝化作用和反硝化作用,从而影响土壤NO排放。
陕西秦岭森林生态系统国家野外科学观测研究站所在的秦岭南坡中段植被垂直分布带明显、完整,类型多样。由低海拔向高海拔依次为落叶阔叶林带、针阔叶混交林带、桦木林带和云冷杉林带等植被带,给我们提供了一个研究海拔梯度对NO通量调控的理想场所。本文的目的是确定不同海拔森林土壤NO通量在季节上的变化以及与土壤环境因子的关系,研究结果可为构建我国不同陆地生态系统NO通量数据库提供秦岭地区更加准确的参数。
1 研究区概况
秦岭火地塘林区位于秦岭南坡中段(33°18′~33°28′N,108°21′~108°39′E),地处陕西省宁陕县境内,其北部与秦岭主梁之一的平河梁相接,南部与宁陕县县城相接,并有210国道和长安河贯穿其中部。林区海拔800~2 500 m,年平均气温9 ℃,年降水量900~1 200 mm,年蒸发量800~950 mm,年日照时间1 100~1 300 h,无霜期170 d,处于我国北亚热带和暖温带的过渡地带。土壤主要有山地棕壤、暗棕壤和山地草甸土。森林覆盖率91.8%,森林植被属温带针阔混交林和寒温性针叶林,森林垂直分布比较明显,树种繁多,达450余种。主要成林树种有:锐齿栎(Quercusalienavar.acuteserrata)、山杨(Pobulusdavidiana)、红桦(Betulaalbo-sinensis)、油松(Pinustabulaeformis)、华山松(Pinusarmandii)、青杄(Piceawilsonii)、云杉(Piceaasperate)、冷杉(Abieschensiensis)等。
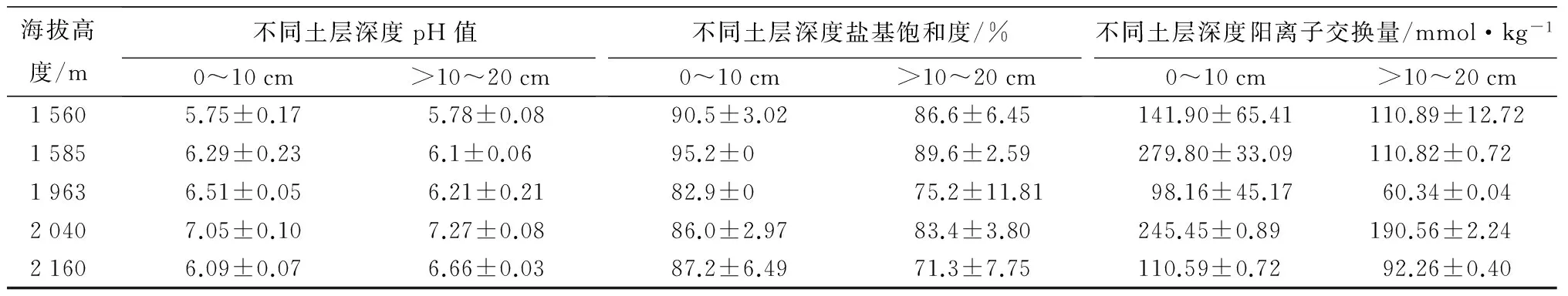
样地特征:在陕西秦岭森林生态系统国家野外科学观测研究站1 560 m的锐齿栎林,1 585 m的油松林,1 963 m的华山松林,2 040 m的青杄林,2 160 m的红桦林五个长期观测采样地开展不同海拔高度森林土壤NO通量监测,在每个样地分别设置4个固定采样点,埋入不锈钢底座。不同海拔森林土壤的基础理化性质见表1。
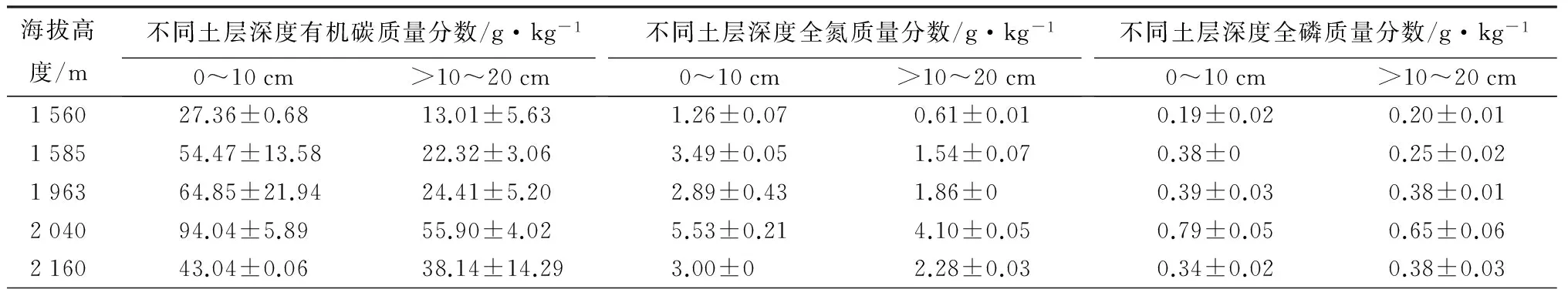
表1 不同海拔样地土壤的主要理化性质
海拔高度/m不同土层深度pH值0~10cm>10~20cm不同土层深度盐基饱和度/%0~10cm>10~20cm不同土层深度阳离子交换量/mmol·kg-10~10cm>10~20cm15605.75±0.175.78±0.0890.5±3.0286.6±6.45141.90±65.41110.89±12.7215856.29±0.236.1±0.0695.2±089.6±2.59279.80±33.09110.82±0.7219636.51±0.056.21±0.2182.9±075.2±11.8198.16±45.1760.34±0.0420407.05±0.107.27±0.0886.0±2.9783.4±3.80245.45±0.89190.56±2.2421606.09±0.076.66±0.0387.2±6.4971.3±7.75110.59±0.7292.26±0.40
注:表中数据为平均值±标准误差。海拔1 560 m时,土壤类型为壤土;海拔在1 585 m时土壤类型为粉砂壤土;海拔在1 963 m时土壤类型为粉砂壤土;海拔在2 040 m时土壤类型为壤土(0~10 cm),粉砂壤土(>10~20 cm);海拔在2 160 m时土壤类型为粉砂壤土。
2 研究方法
从2014年10月至2015年10月对秦岭火地塘林区5个不同海拔森林土壤NO排放进行了一整年的监测。NO气体采集利用静态箱法,在各个不同海拔样地分别设置4个底座,采样频率大概为生长季(2015年5—9月)每月2次,非生长季(2014年10月—2015年4月)每月1次。采样时间多选择在上午08:00—11:00,夏季为07:00—10:00。采样箱用2 mm厚不锈钢板压制成,外覆绝热材料,底面积50 cm×50 cm,箱体高35 cm。观测时将采样箱放到已插入地下5 cm深处的不锈钢底座外缘四周的凹槽中,并用水密封。在采样箱箱口高度的树枝上挂有温度计,分别读取采样开始前和采样完成后的气温。采样开始前,在采样箱箱口高度采集气样作为第一个样品作为箱内初始浓度,采样箱箱盖装有用于采气用的胶皮管和采气三通阀以及气压平衡管(内径为4 mm,长度为4 cm)[25],采样时将平衡管打开,其余罩箱时间关闭平衡管,分别抽取盖箱后30 min时的气体样品,每一次采样都用手持式采气泵抽取采样箱内气体置于5 L密封气袋中,采样持续时间1.5 min,然后运回实验室在2 h内完成分析。准确记录每次罩箱时间和每次从箱内取样的时间。在实验室用化学发光氮氧化物分析仪(ML 9841 B Nitrogen Oxides Analyzer)测定气袋内样品NO浓度。
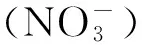
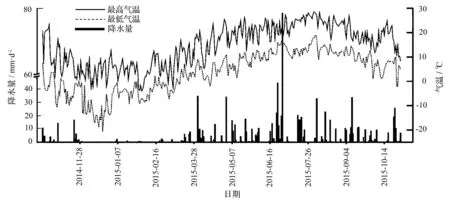
图1 秦岭火地塘林区林场降雨和日气温
NO的通量用公式(1)计算:
(1)
式中,F为单位时间、单位面积的NO通量(104μg·m-2·h-1),dc/dt为单位时间内采样箱内NO浓度的变化(10-3μL·L-1·h-1),H为采样箱高度(m),M为NO的摩尔质量(g·mol-1),P为大气压(MPa),R为气体常数(cm3·MPa·K-1·mol-1),T为通量箱内气温(K)。
NO的年通量是将所有每两个相邻采样日期之间的NO通量加起来,每两个相邻采样日期之间的NO通量Xn用公式(2)计算:
(2)
式中,fn表示特定时间段的最后一天的NO通量,fn-1表示特定时间段的初始一天的NO通量,Dn是特定时间段的最后一天,Dn-1是特定时间段的初始一天[27]。
利用Excel进行数据处理,以每个海拔梯度4个点各指标的平均值表示该海拔梯度的土壤理化性质和NO通量。采用Origin Pro 2015进行作图,并使用SPSS 21.0对观测数据进行相关分析和逐步线性回归分析,显著水平设定为α=0.05。
3 结果与分析
3.1 不同海拔环境和土壤因素的变化
不同海拔5个样地0~10 cm地温的季节变化规律明显(图2),且季节变化规律基本一致,表现为雨热同季特征。随着海拔的升高,地温基本呈逐渐下降的趋势,锐齿栎(1 560 m)、油松(1 585 m)、华山松(1 963 m)、青杄(2 040 m)、红桦(2 160 m)5个样地的年平均地温分别为9.5、9.3、7.9、5.4、6.3 ℃。红桦林、华山松林和锐齿栎林的最高地温出现在2015年7月25日,分别为13.8、15.1、17.0 ℃,青杄林和油松林的最高地温出现在2015年6月27日,分别为12.9、16.6 ℃。5个样地的最低地温均出现在2015年2月8日,由低海拔到高海拔依次为1.8、0.7、0.2、-1.5、-1.5 ℃。
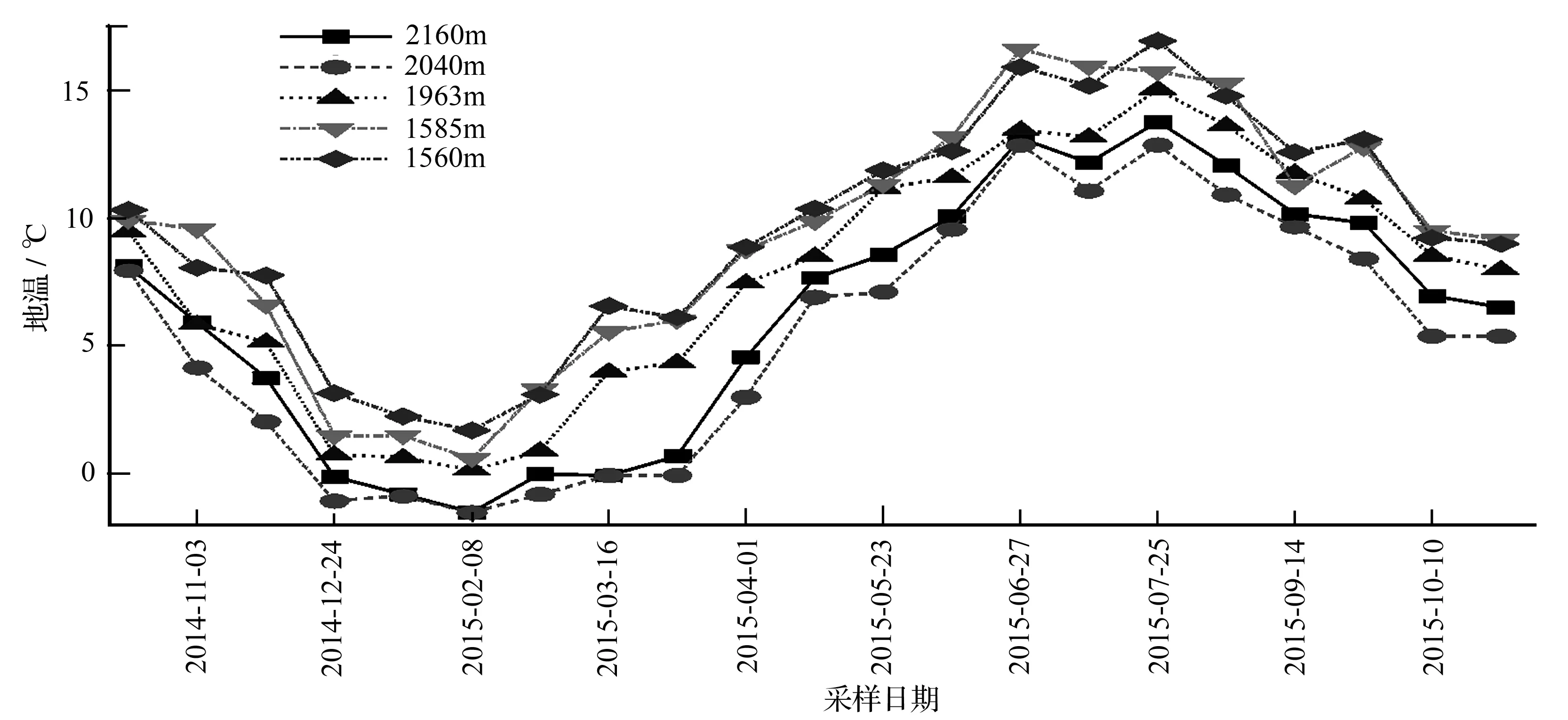
图2 地温(10 cm)的变化
不同海拔表层土壤(0~20 cm)土壤孔隙充水率变化规律基本一致,在非生长季波动较大,在生长季波动较小(图3a)。红桦、青杄和华山松林的最大土壤孔隙充水率均出现在2014年12月24日,分别为55.54%、115.59%、101.10%,油松林的最大土壤孔隙充水率为94.99%,锐齿栎林的最大土壤孔隙充水率为58.10%。最低的土壤孔隙充水率出现在2015年2月8日红桦林土壤中,土壤孔隙充水率值不到20%。5个样地由低海拔到高海拔,土壤孔隙充水率年平均值分别为锐齿栎林46.94%、油松林74.07%、华山松林55.63%、青杄林46.94%、红桦林35.60%。
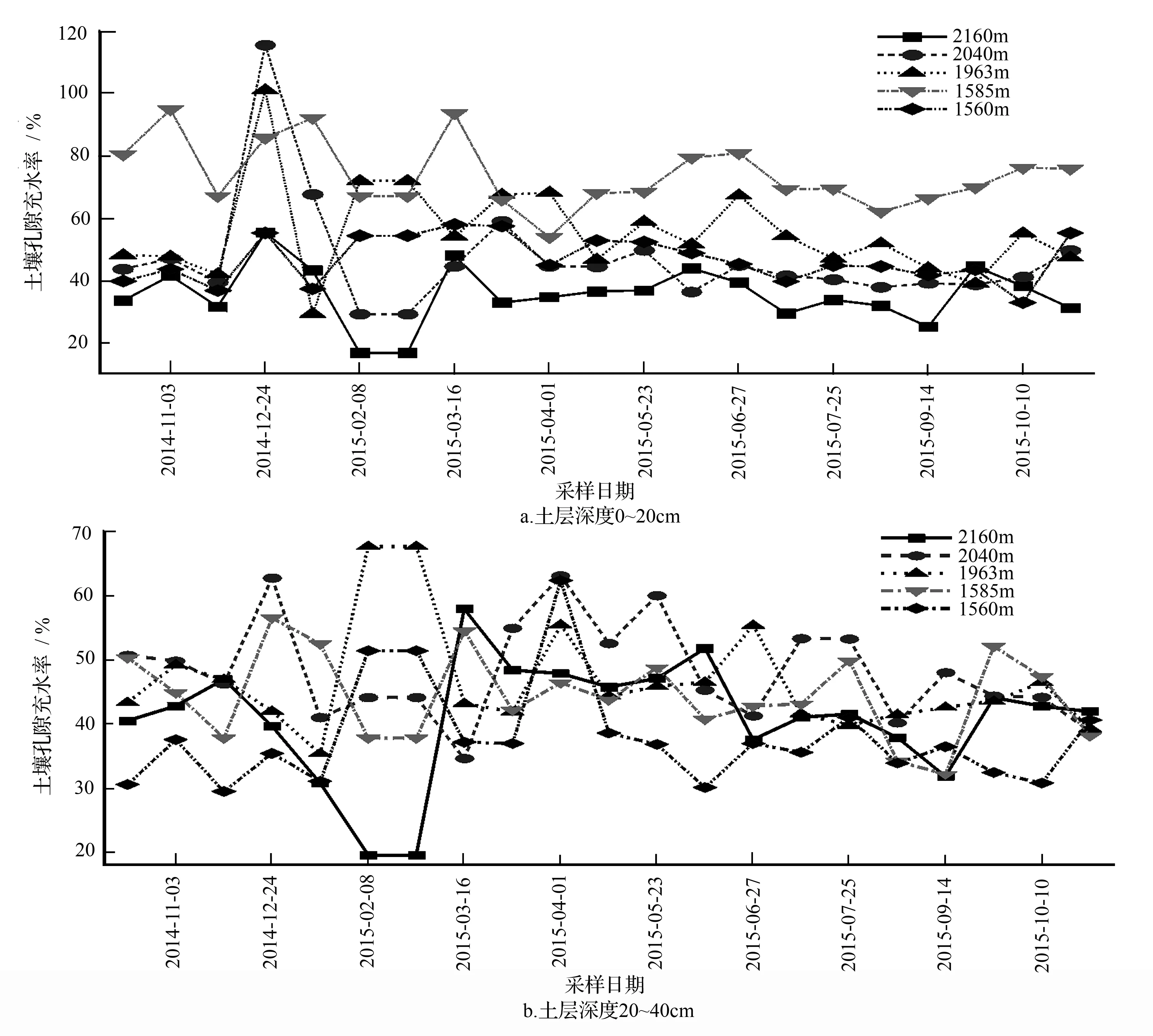
图3 土壤孔隙充水率变化
不同海拔20~40 cm土壤层土壤孔隙充水率季节变化不明显,表现为非生长季波动较大,生长季波动较小(图3b)。青杄和锐齿栎林的最大土壤孔隙充水率均出现在2015年4月1日,分别为63.20%、62.47%。油松林最大土壤孔隙充水率出现在2014年12月24日,达到56.64%。红桦林最大土壤孔隙充水率出现在2015年3月16日,为58.03%。最低土壤孔隙充水率也出现在2015年2月8日红桦林土壤中,土壤孔隙充水率值不到20%。5个样地由低海拔到高海拔,土壤孔隙充水率年平均值分别为锐齿栎林38.03%、油松林44.55%、华山松林46.74%、青杄林48.32%、红桦林40.92%。
不同海拔森林土壤0~20 cm硝态氮和铵态氮的质量分数随季节的变化有所不同(图4)。5个样地中硝态氮质量分数的年平均值由高海拔到低海拔分别为红桦20.12 μg·g-1、青杄24.18 μg·g-1、华山松12.08 μg·g-1、油松18.06 μg·g-1、锐齿栎4.85 μg·g-1。同时,由高海拔到低海拔铵态氮质量分数依次为14.19、14.0、17.32、16.52、13.74 μg·g-1。从图4中可以看出,在植物生长季土壤中硝态氮的质量分数会增加且数值波动较大,而铵态氮质量分数比较低,基本处于平稳状态。在非生长季铵态氮的质量分数比较大且波动明显,而硝态氮质量分数则比较低。

图4 表层土壤(0~20 cm)硝态氮、铵态氮质量分数
3.2 不同海拔NO通量动态
不同海拔森林土壤NO排放在时间和空间上的变化都很大(图5)。整个观测期NO排放均保持在较低水平,在植物生长季,土壤NO排放速率基本呈“W”形变化,受到降雨的影响,排放峰多出现在降雨后,红桦、青杄和锐齿栎林均在2015年8月26日达到最大值,分别为6.23、7.46、13.95 μg·m-2·h-1。而华山松和油松林分别于2015年5月23日和2015年3月10日达到生长季最大值,分别为5.49、9.04 μg·m-2·h-1。在非生长季,大部分样地的NO排放先减少后增加,而且监测有NO吸收。各样地土壤NO吸收最大值和出现日期分别为红桦林2015年1月22日-4.80 μg·m-2·h-1、青杄林2014年11月3日-6.71 μg·m-2·h-1、华山松林2015年2月8日-5.61 μg·m-2·h-1、油松林2014年10月20日-4.28 μg·m-2·h-1、锐齿栎林2015年3月10日-6.81 μg·m-2·h-1。不同海拔土壤NO排放通量有所不同,5个样地的年平均值分别为红桦林1.27 μg·m-2·h-1、青杄林0.97 μg·m-2·h-1、华山松林0.87 μg·m-2·h-1、油松林1.21 μg·m-2·h-1、锐齿栎林2.30 μg·m-2·h-1。
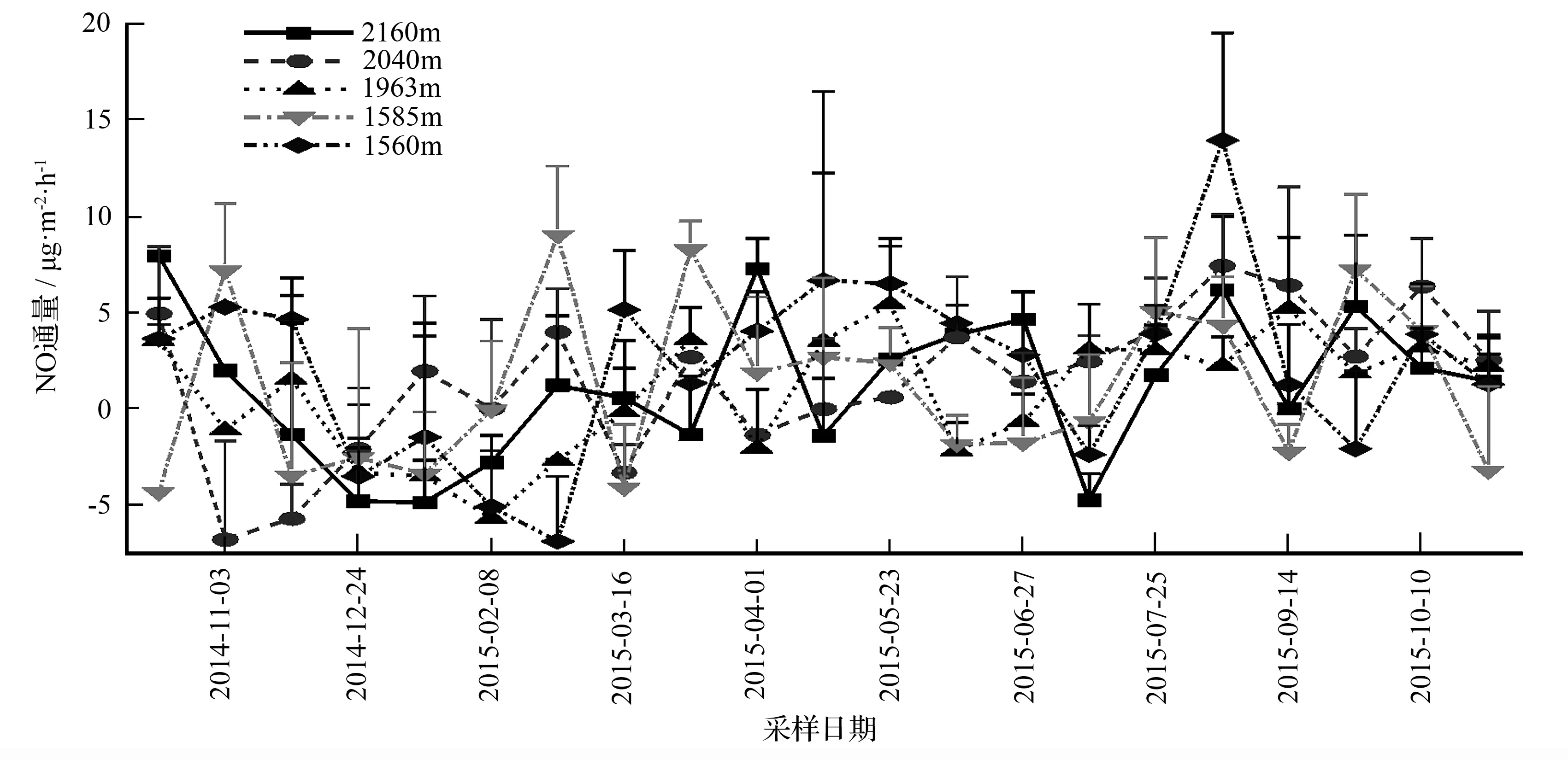
图5 不同海拔森林土壤NO通量的季节排放动态
3.3 不同海拔NO通量的影响因素
不同海拔5个样地土壤NO通量均与地温显著相关(P<0.05),华山松林土壤NO排放与土壤孔隙充水率负相关(P<0.05),油松林土壤NO排放与土壤铵态氮含量负相关(P<0.05,表2)。逐步线性回归结果(表3)同样可以表明,温度显著影响着不同海拔森林土壤NO排放通量。从整个海拔梯度来看,土壤NO排放随着地温呈指数增长(图6),相对较大的NO排放出现在40%~45%土壤孔隙充水率(图7)。
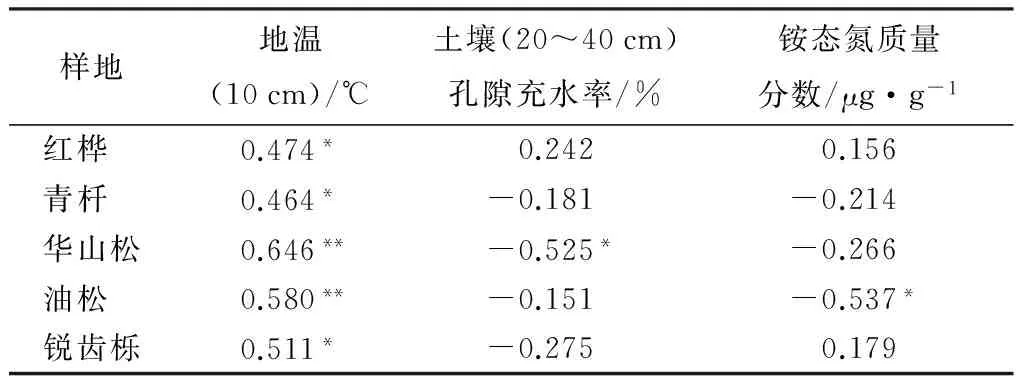
表2 不同海拔样地NO通量与环境和土壤因子的相关关系
注:*代表相关性达到0.05的显著水平,** 代表相关性达到0.01的显著水平。
表3 不同海拔样地NO通量与环境和土壤因子逐步线性回归分析
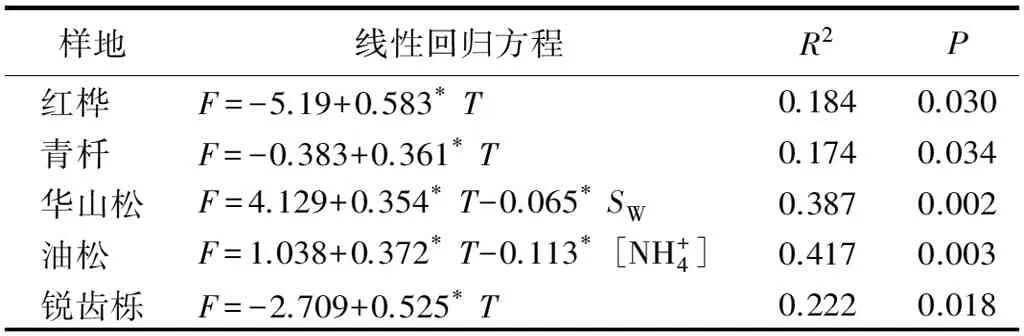
样地线性回归方程R2P红桦F=-5.19+0.583*T0.1840.030青杄F=-0.383+0.361*T0.1740.034华山松F=4.129+0.354*T-0.065*SW0.3870.002油松F=1.038+0.372*T-0.113*[NH+4]0.4170.003锐齿栎F=-2.709+0.525*T0.2220.018
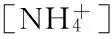
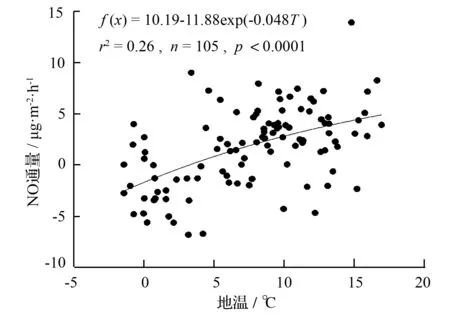
图6 土壤NO排放与地温之间的相互关系
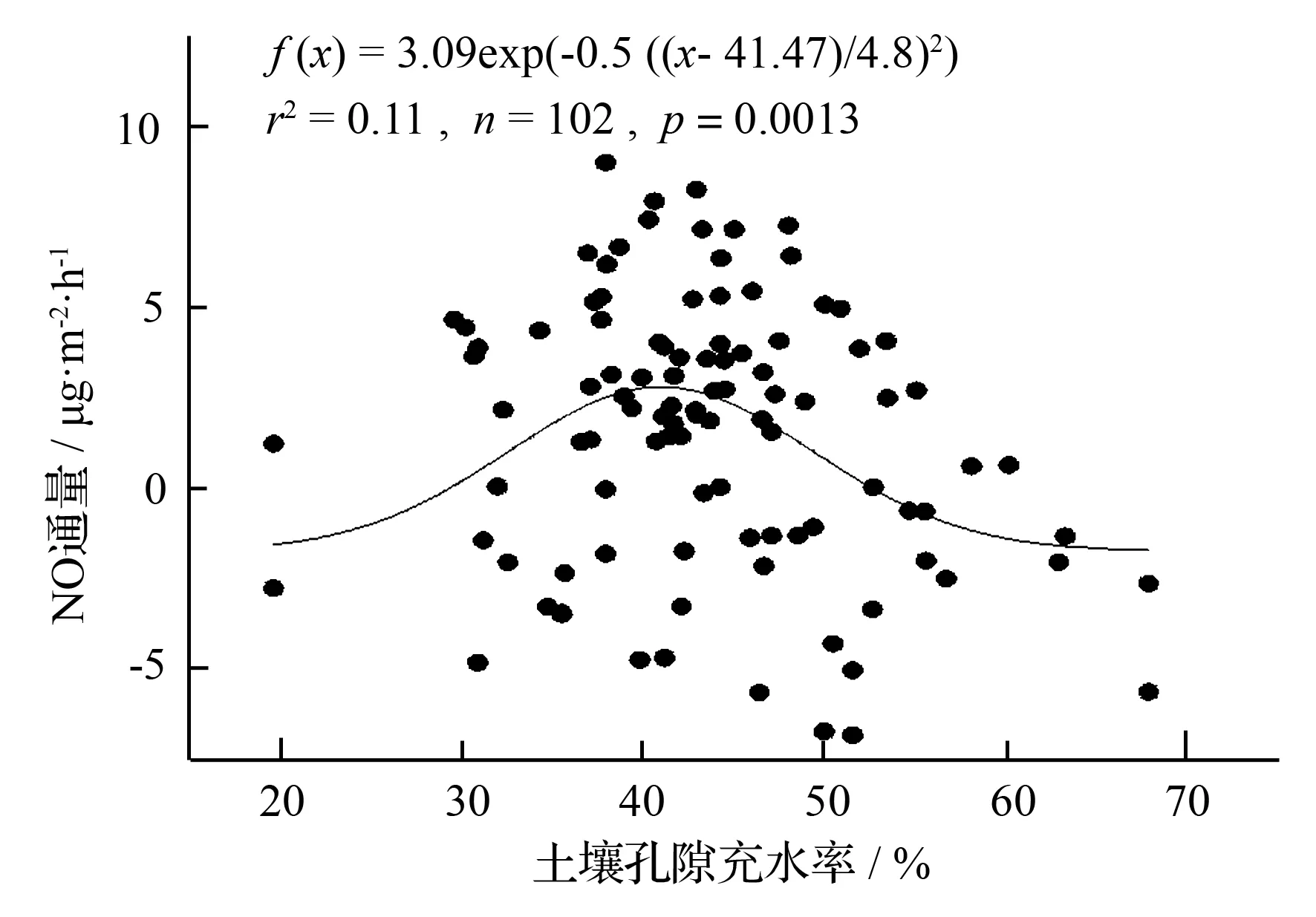
图7 土壤NO排放与WFPS(20~40 cm)之间的相互关系
4 讨论
除华山松外,NO年排放总量随海拔的升高而减少,最大排放量在海拔1 560 m,为0.19 kg·ha-2·a-1;最小排放量在海拔2 160 m,为0.06 kg·ha-2·a-1。从海拔1 560 m到2 040 m,土壤有机质和黏粒含量随着海拔的升高而增加,因此高海拔土壤持水率较高。本研究中,NO排放主要集中在植物生长季,而生长季降雨较多,高海拔土壤土壤孔隙充水率一直保持在较高水平,导致土壤中所产生的NO难于扩散到大气中[28],而在缺氧条件下产生的NO被反硝化细菌消耗的量较高[29],因此高海拔NO排放低。海拔1 963 m华山松林土壤NO年排放量最低,可能是由于大小蠹的爆发导致华山松叶片损失、生长延缓,影响华山松林木营养和水分的运输,并且也导致枯落物的分解处于低至中等水平[30],从而影响其碳氮循环[31],最终影响土壤NO排放。
Ludwig et al[32]指出,在基质和水分等其他条件不受限制时,NO排放随着温度的升高而增加,因为对于生物过程来说,在一定范围内酶活性随温度升高而增加。本研究中我们得出土壤NO排放随地温呈指数增长(图6),这与Pilegaard et al[11]、Van Dijk和Meixner[33]、Schindlbacher et al[34]的研究结果相一致。土壤孔隙充水率可以从多个途径影响NO的排放,例如,可以通过调节硝化细菌和反硝化细菌的活性,从而决定是硝化反应还是反硝化反应占主导地位;显著影响基质的产生以及基质和产物在土壤中的迁移;显著影响NO在土壤中的产生和消耗过程[35]。Ludwig et al[32]得出最有利于土壤NO排放的土壤孔隙充水率为20%左右,Pilegaard et al[11]和Wu et al[17]也指出最大的NO排放通量出现在中等偏低的土壤孔隙充水率条件下。在我们的研究中,相对较大的NO排放出现在40%~45%土壤孔隙充水率(图7),这与Li et al[19]、Gasche和Papen[36]的结果相一致。但是,Schindlbacher et al[34]通过对欧洲不同森林的研究发现,不同的土壤最适土壤孔隙充水率不同,变化范围15%~65%,这可能是由不同的土壤质地、容重或微生物群落等引起的。
土壤NO的排放或吸收是多种微生物以及化学反应综合作用的结果,不同环境条件下有不同的反应过程[6-7]。Medinets et al[7]指出,以下三个过程可能会导致土壤NO吸收:(1)反硝化作用。此反应在厌氧条件下作用更大,同时土壤有机碳含量和硝态氮含量比较高时也会促进此反应的进行;(2)异养硝化菌的反硝化作用。此反应多发生于酸性针叶林中,土壤中有机质含量和土壤C/N较高;(3)DNRA(异化的硝酸根还原为铵根)过程。此反应多发生在厌氧,土壤黏粒含量高的土壤中,同时土壤中的有机质含量高。一般来说,土壤的黏粒含量越高,土壤的肥力越强,但土壤的透水性和透气性却越差[37]。本研究中,各样地均监测有NO吸收尤其是在非生长季。红桦林土壤NO吸收最大值为-4.80 μg·m-2·h-1,2015年1月22日其表层土壤(0~20 cm)土壤孔隙充水率为43.33%,土壤黏粒含量为8.4%,同时土壤有机碳含量为43.04 g·kg-1,硝态氮和铵态氮的含量较低,所以该NO吸收可能是由“DNRA”过程引发的。青杄林土壤NO吸收最大值为-6.71 μg·m-2·h-1,发现出现该最大值的2014年11月3日其表层土壤(0~20 cm)土壤孔隙充水率为47.19%,硝态氮含量为26.41 μg·g-1,土壤有机碳含量高达94.04 g·kg-1,有利于反硝化作用。华山松林土壤NO吸收最大值发生在2015年2月8日,其表层土壤(0~20 cm)土壤孔隙充水率为72.13%,而且土壤的冻结使得土壤团聚体分裂,为反硝化作用提供了可利用的碳源[38]。同时表层土壤pH为6.51,C/N高达22.44,促进了异养硝化菌的反硝化作用。油松林表层土壤pH为6.29,有机碳含量为54.47 g·kg-1,C/N为15.61,发生异养硝化菌的反硝化作用的同时可能伴随有“DNRA”过程。同样,2015年3月10日锐齿栎林表层土壤的土壤孔隙充水率较高,土壤C/N为21.71,而且冻融交替期土壤的冻结使得土壤团聚体分裂,为反硝化作用提供了可利用的碳源[38]。
在一些温带地区,森林土壤NO排放比较低,Kitzler et al[39]认为,这可能与当地大气氮沉降和土壤pH有关。火地塘林区受到人为干扰比较少,而且只有一条公路穿过,车流量小,大气氮沉降低。Ormeci et al[40]研究了pH值在4.3~8.4范围内NO排放变化特征(20 ℃,43%土壤孔隙充水率),发现pH<5时,NO排放最高,而pH介于5~8之间时,NO排放极低,当pH>8时,NO排放稍有增加。本研究中土壤pH介于5.75~7.27之间,是该地区土壤NO排放低的重要原因之一。另外,在植物非生长季,土壤温度很低,抑制了土壤微生物的活性,并且可能伴随有反硝化作用、异养硝化细菌反硝化作用以及“DNRA”过程等,故土壤净NO排放很低甚至出现NO吸收。虽然土壤NO排放主要集中在植物生长季,但在植物生长季,土壤铵态氮含量较低,不利于硝化反应,而且高海拔地区土壤孔隙充水率一直保持在较高水平,所以整个监测期NO排放较低。
5 结论
土壤排放NO是微生物活动以及化学反应综合作用的结果,且NO排放具有较大的时空变异性。本地区土壤NO通量保持在较低水平,土壤NO排放主要集中在植物生长季,在非生长季出现大量的NO吸收。不同海拔森林土壤NO年排放总量分别为2 160 m红桦林0.06 kg·ha-2·a-1,2 040 m青杄林0.08 kg·ha-2·a-1,1 963 m华山松林0.02 kg·ha-2·a-1,1 585 m油松林kg·ha-2·a-1,1 560 m锐齿栎林0.19 kg·ha-2·a-1。
NO通量受到地温、土壤含水量以及土壤有效氮的影响。不同海拔森林土壤NO通量均与地温显著相关(P<0.05)。华山松林土壤NO通量与土壤孔隙充水率负相关(P<0.05)。油松林土壤NO通量与铵态氮质量分数负相关(P<0.05)。
[1] BRADLEY M J, JONES B M. Reducing global NOxemissions: Developing advanced energy and transportation technologies[J]. AMBIO,2002,31(2):141-149.
[2] KHAN S, CLOUGH T J, GOH K M, et al. Influence of soil pH on NOxand N2O emissions from bovine urine applied to soil columns[J]. New Zealand Journal of Agricultural Research,2011,54(4):285-301.
[3] VITOUSEK P M, MOONEY H A, LUBCHENCO J, et al. Human domination of earth’s ecosystems[J]. Science,1997,277:494-499.
[4] CONRAD R, SMITH K A. Soil microbial processes and the cycling of atmospheric trace gases[J]. Philosophical Transactions: Physical Sciences and Engineering,1995,351:219-230.
[5] WILLIAMS E J, HUTCHINSON G L, FEHSENFELD F C. NOxand N2O emissions from soil[J]. Global Biogeochemical Cycles,1992,6(4):351-388.
[6] PILEGAARD K. Processes regulating nitric oxide emissions from soils[J]. Philosophical Transactions of the Royal Society B: Biological Sciences,2013,368:1-8.
[7] MEDINETS S, SKIBA U, RENNENBERG H, et al. A review of soil NO transformation: Associated processes and possible physiological significance on organisms[J]. Soil Biology and Biochemistry,2015,80(80):92-117.
[8] VERCHOT L V, DAVIDSON E A, CATTNIO H, et al. Land use change and biogeochemical controls of nitrogen oxide emissions from soils in eastern Amazonia[J]. Global Biogeochemical Cycles,1999,13(1):31-46.
[9] DAVIDSON E A, KELLER M, ERICKSON H E, et al. Testing a Conceptual Model of Soil Emissions of Nitrous and Nitric Oxides[J]. Bioscience,2000,50(8):667-680.
[10] ROSENKRANZ P, BRÜGGEMANN N, PAPEN H, et al. N2O, NO and CH4exchange and microbial N turnover over a Mediterranean pine forest soil[J]. Biogeosciences,2006,3(2):121-133.
[11] PILEGAARD K, SKIBA U, AMBUS P, et al. Factors controlling regional differences in forest soil emission of nitrogen oxides (NO and N2O)[J]. Biogeosciences,2006,3(4):651-661.
[12] PURBOPUSPITO J, VELDKAMP E, BRUMME R, et al. Trace gas fluxes and nitrogen cycling along an elevation sequence of tropical montane forests in Central Sulawesi, Indonesia[J]. Global Biogeochemical Cycles,2006,20(3):1-11.
[13] DAVIDSON E A, KINGERLEE W. A global inventory of nitric oxide emissions from soils[J]. Nutrient Cycling in Agroecosystems,1997,48(1):37-50.
[14] VENTEREA R T, GROFFMAN P M, VERCHOT L V, et al. Nitrogen oxide gas emissions from temperate forest soils receiving long-term nitrogen inputs[J]. Global Change Biology,2003,9(3):346-357.
[15] GOLDBERG S D, GEBAURE G. N2O and NO fluxes between a Norway spruce forest soil and atmosphere as affected by prolonged summer drought[J]. Soil Biology and Biochemistry,2009,41(9):1986-1995.
[16] BARGSTEN A, FALGE E, PRITSCH K, et al. Laboratory measurements of nitric oxide release from forest soil with a thick organic layer under different understory types[J]. Biogeosciences,2010,7(5):1425-1441.
[17] WU X, BRUGGEMANN N, GASCHE R, et al. Environmental controls over soil-atmosphere exchange of N2O, NO and CO2in a temperate Norway spruce forest[J]. Global Biogeochemical Cycles,2010,24:1-16.
[18] ERICKSON H E, PERAKIS S S. Soil fluxes of methane, nitrous oxide, and nitric oxide from aggrading forests in coastal Oregon[J]. Soil Biology and Biochemistry,2014,76(1) :268-277.
[19] LI D J, WANG X M, MO J M, et al. Soil nitric oxide emissions from two subtropical humid forests in south China[J]. Journal of Geophysical Research,2007,112(D23):209-232.
[20] LUO G J, BRÜGGEMANN N, WOLF B, et al. Decadal variability of soil CO2, NO, N2O and CH4fluxes at the Höglwald Forest, Germany[J]. Biogeosciences,2012,9(11):1741-1763.
[21] BURKE I C, LAUENROTH W K, RIGGLE R, et al. Spatial variability of soil properties in the short grass steppe: the relative importance of topography, grazing, microsite, and plant species in controlling spatial patterns[J]. Ecosystems,1999,2(5):422-438.
[22] GROFFMAN P M, ZAK D R, CHRISTENSEN S, et al. Early spring nitrogen dynamics in a temperate Forest landscape[J]. Ecology,1993,74(5):1579-1585.
[23] ZAK D R, HAIRSTON A, GRIGAL D F. Topographic influences on nitrogen cycling within an upland Pin Oak Ecosystem[J]. Forest Science,1991,37(1):45-53.
[24] 李鹏,李占斌,郑郁.不同海拔对干热河谷地区土壤理化性质及可蚀性的影响[J].水土保持通报,2011,31(4):103-107.
[25] HUTCHINSON G L, MOSIER A E. Improved method for field measurement of nitrous oxide fluxes[J]. Soil Science Society of America Journal,1981,45(2):311-316.
[26] MERINO A, BATALLN P, MACAS F. Responses of soil organic matter and greenhouse gas fluxes to soil management and land use changes in a humid temperate region of southern Europe[J]. Soil Biology and Biochemistry,2004,36(6):917-925.
[27] DU R, LU D R, WANG G C. Diurnal, seasonal and inter-annual variation of N2O fluxes from native semi-arid grassland soils of inner Mongolia[J]. Soil Biology and Biochemistry,2006,38(12):3474-3482.
[28] VAN DIJK S M, GUT A, KIRKMAN G A, et al. Biogenic NO emissions from forest and pasture soils: Relating laboratory studies to field measurements[J]. Journal of Geophysical Research: Atmospheres,2002,107(D20):251-269.
[29] YE R W, AVERILL B A, TIEDJE J M. Denitrification: production and consumption of nitric oxide[J]. Applied and Environmental Microbiology,1994,60(4):1053.
[30] CUNNINGHAM S A, PULLEN K R, COLLOFF M J. Whole-tree sap flow is substantially diminished by leaf herbivory[J]. Oecologia,2009,158(4):633-640.
[31] COBB T P, HANNAM K D, KISHCHUK B E, et al. Wood-feeding beetles and soil nutrient cycling in burned forests: implications of post-fire salvage logging[J]. Agricultural and Forest Entomology,2010,12(1):9-18.
[32] LUDWIG J, MEIXNER F X, VOGEL B, et al. Soil-air exchange of nitric oxide: An overview of processes, environmental factors, and modeling studies[J]. Biogeochemistry,2001,52(3):225 -257.
[33] VAN DIJK S M, MEIXNER F X. Production and consumption of NO in forest and pasture soils from the Amazon basin[J]. Water Air and Soil Pollution,2001,1(5):119-130.
[34] SCHINDLBACHER A, ZECHMEISTER-BOLTENSTERN S, BUTTERBACH-BAHL K. Effects of soil moisture and temperature on NO, NO2and N2O emissions from European forest soils[J]. Journal of Geophysical Research,2004,109(D17):1-12.
[35] DAVIDSON E A. Soil water content and the ratio of nitrous oxide to nitric oxide emitted from soil//Oremland RS. Biogeochemistry of Global Change: Radiatively Active Trace Gases[M]. London: Chapman and Hall,1993:369-386.
[36] GASCHE R, PAPEN H. A 3-year continuous record of nitrogen trace gas fluxes from untreated and limed soil of a N-saturated spruce and beech forest ecosystem in Germany: 2.NO and NO2fluxes[J]. Journal of Geophysical Research,1999,104(D15):18505-18520.
[37] 孙向阳.土壤学[M].北京:中国林业出版社,2005.
[38] GROFFMAN P M, HARDY J P, DRISCOLL C T, et al. Snow depth, soil freezing, and fluxes of carbon dioxide, nitrous oxide and methane in a northern hardwood forest[J]. Global Change Biology,2006,12(9):1748-1760.
[39] KITZLER B, ZECHMEISTER-BOLTENSTERN S, HOLTERMANN C, et al. Controls over N2O, NOxand CO2fluxes in a calcareous mountain forest soil[J]. Biogeosciences,2006,3(4):383-395.
[40] ORMECI B, SANIN S L, PEIRCE J J. Laboratory study of NO flux from agriculture soil: effects of soil moisture, pH, and temperature[J]. Journal of Geophysical Research,1999,104(D1):1621-1629.
NO Flux in Forest Soil at Different Elevation of Huoditang Area in the Qinling Mountains//
Li Yutian, Pang Junzhu, Peng Changhui, Zhang Shuoxin(Northwest A&F University, Yangling 712100, P. R. China); Hou Lin, Chen Shujun(State Key Field Observation and Research Station of Qingling Ecosystem); Wang Xiaoke, Zhang Hongxing(Research Center for Eco-environmental Sciences, Chinese Academy of Sciences)//
Journal of Northeast Forestry University,2017,45(2):42-49,56.
Qinling Mountains; Forests; Soil; NO flux
1)国家自然科学基金项目(41101182);国家级创新项目;西北农林科技大学基本科研业务费(Z109021106);西北农林科技大学博士科研启动基金项目(Z109021102)。
李玉田,男,1991年4月生,西北农林科技大学林学院,硕士研究生。E-mail:65737568@qq.com。
庞军柱,西北农林科技大学林学院,讲师。E-mail:pangjunzhu@126.com。
2016年11月4日。
S718.5
责任编辑:潘 华。
