不同基质对盆栽闭鞘姜光合特性和生长的影响
2017-03-10刘晓荣欧成宇李冬梅刘小飞
刘晓荣,欧成宇,李冬梅,刘小飞
(1.广东省农业科学院环境园艺研究所,广东 广州 510640;2.仲恺农业工程学院园艺园林学院,广东 广州 510225)
闭鞘姜(Costus speciosus)为姜科闭鞘姜属多年生草本植物,别名广商陆、水焦花、老妈妈拐棍[1],广布于东南亚、南亚热带地区,主要分布在广东、广西、云南等省区。喜温暖、湿润环境下生长,生长适温为20~30℃,在华南地区春、夏、秋三季均可生长,冬季呈半休眠状态。除药用、食用外,还可作鲜切花、干花和庭院绿化,丛植于庭院小区、公园、花坛等,红色革质状的穗状花序形状独特极为雅致,是极具开发前景的一类野生花卉资源。目前对其研究主要集中于药用价值,观赏性方面的研究甚少。建立温室下盆栽技术,进而推动多功能植物闭鞘姜在观赏方面的产业化发展,其意义重大。我国和东南亚、非洲一些国家对闭鞘姜的药用性方面开展了相关研究[2-4],栽培技术方面国内仅见曾凌云、赵国祥等[5-6]介绍。光合作用是植物生长的生理基础,可作为判断植物长势的指标[7],但目前有关闭鞘姜盆栽栽培技术方面的研究未见有报道。本研究以盆栽闭鞘姜为研究对象,分析不同基质栽培条件对其光合生长特性和生长发育的影响,为筛选闭鞘姜盆栽种植基质材料提供科学依据。
1 材料与方法
1.1 试验材料
供试材料为闭鞘姜一年生地下根茎,以1~2个芽点切断横走根茎若干块,每块大小约5 cm×5 cm,种植于盆内。
1.2 试验方法
试验在广东省农业科学院环境园艺研究所白云基地温室内进行,基质有红土、泥炭、椰糠、珍珠岩,按照不同比例混合而成,分别为S1:红土+珍珠岩(3∶1)、S2:泥炭∶松树皮∶珍珠岩 (3∶2∶1)、S3:泥炭∶椰糠∶珍珠岩(1∶1∶1)、S4:泥炭∶椰糠∶珍珠岩(1∶2∶2)、S5:泥炭∶椰糠∶珍珠岩(2∶1∶2)、S6:泥炭∶椰糠∶珍珠岩 (2∶2∶1)共6个处理。
选择处于生长期、生长状态良好植株,每个基质分选5株,选择叶位相近的成熟饱满的叶片,挂牌标记。2016年8月,选择3个连续晴天,在大棚里用Yaxin-1102便携式光合蒸腾仪测定不同基质净光合速率、蒸腾速率及环境因子的日变化,开放气路测定。从8:30 ~16:30,每隔2 h测定1次,共测3 d,每个处理测定5株,3次重复。测定指标包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci) 和叶片温度,同时测定大气温度和相对湿度(R H),以3 d测定的平均值进行相关分析。
1.3 数据分析
采用 SAS 8.1软件[8]和 Microsoft Excel进行数据统计分析和绘图。
2 结果与分析
2.1 光合有效辐射(PAR)和大气CO2摩尔分数日变化
图1显示,PAR日变化呈单峰型曲线,在观测时间内,初始直线性地升高,在12:30时达到最高(391.29 μmol/m2·s),之后快速下降;大气CO2摩尔分数日变化呈折线变化,先缓慢性地下降,12:30时降到366.06 μmol/mol,后又上升,在14:30出现峰值,随后又下降。
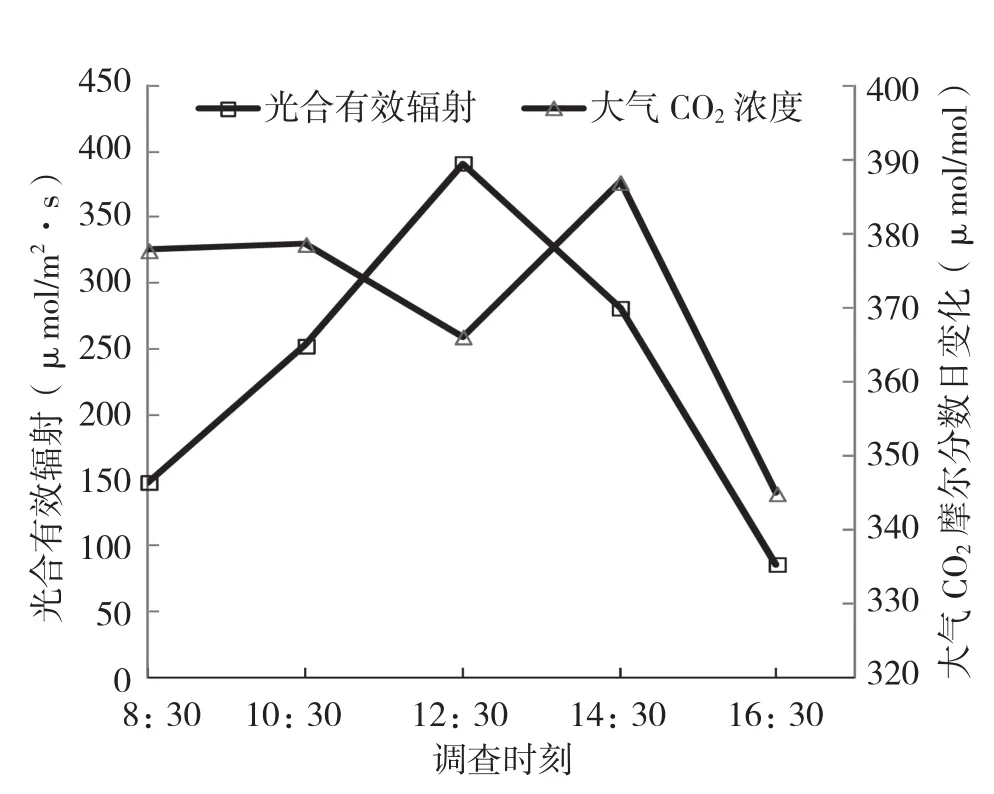
图1 光合有效辐射与大气CO2摩尔分数日变化
2.2 不同基质下盆栽闭鞘姜叶片气孔导度(Gs)和胞间CO2浓度(Ci)的日变化
由图2A可知,不同基质下闭鞘姜叶片的Gs日变化呈单峰曲线,其中S4、S5、S6在10:30达到峰值,而S1、S2、S3处理下的盆栽闭鞘姜Gs峰值出现在14:30。图2B显示,不同基质处理的盆栽闭鞘姜叶片的Ci日变化呈单峰变化,基质S4、S5和S6处理下的盆栽闭鞘姜Ci在10:30出现峰值,基质S2、S3处理下的Ci峰值在14:30出现,而基质S1处理下的Ci在12:30达到最高点。
2.3 不同基质下盆栽闭鞘姜叶片净光合速率(Pn)的日变化
基质S1处理下的盆栽闭鞘姜叶片Pn的日变化呈双峰曲线(图3),在10:30时形成第1次峰值、达11.48 μmol/m2·s,之后下降,第2次峰值13.35 μmol/m2·s出现在14:30;其他5个基质处理下的盆栽闭鞘姜叶片Pn日变化规律相似,均呈单峰型,在12:30达到峰值。6个不同基质处理下盆栽闭鞘姜叶片的Pn日均值分别为7.77、8.50、9.43、12.16、9.71、9.00 μmol/m2·s。
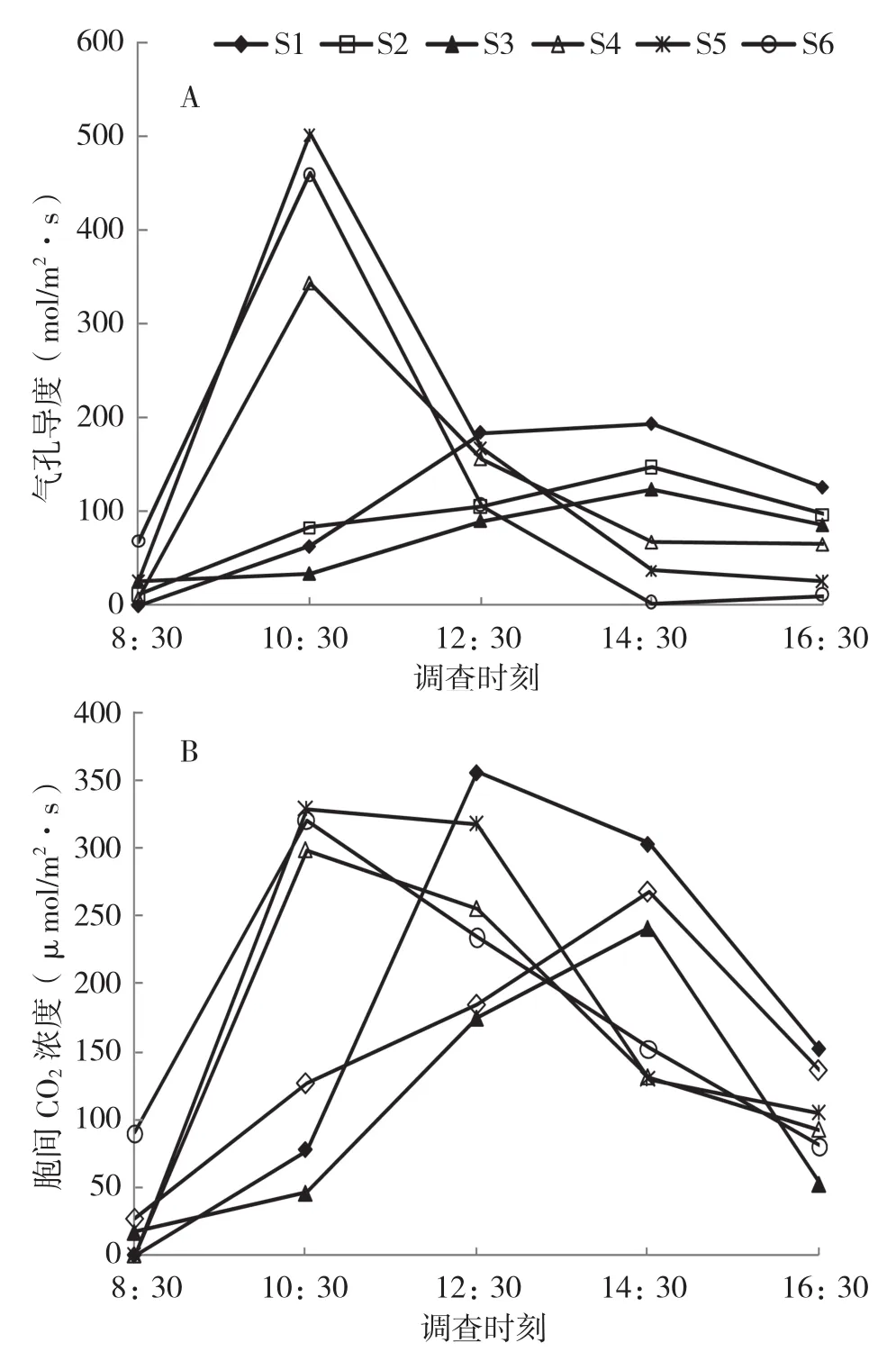
图2 不同基质下盆栽闭鞘姜叶片气孔导度和胞间CO2浓度的日变化
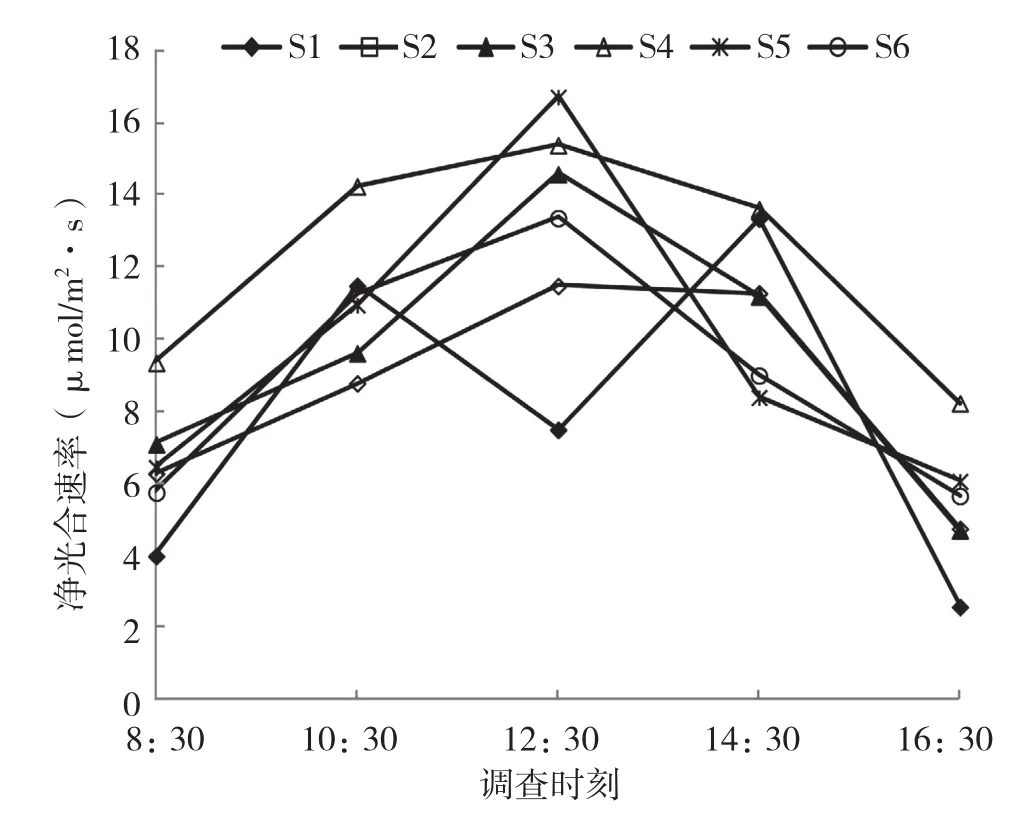
图3 不同基质下盆栽闭鞘姜叶片净光合速率的日变化
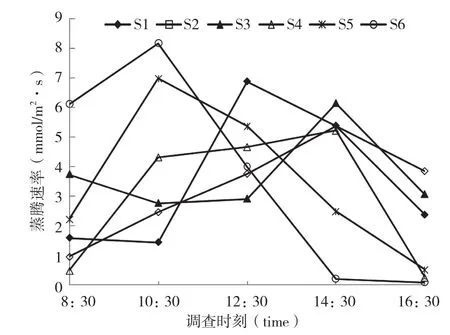
图4 不同基质下盆栽闭鞘姜叶片蒸腾速率的日变化
2.4 不同基质下盆栽闭鞘姜叶片蒸腾速率(Tr)的日变化
从图4可以看出,不同基质下盆栽闭鞘姜Tr日变化均呈单峰曲线,但出现峰值时间不同,其中S1在12:30出现峰值,S2、S3和S4在14:30达到最大,S5、S6在10:30出现峰值。基质S6的Tr最大(8.17 mmol/m2·s),基质S4的Tr最小(5.22 mmol/m2·s)。不同基质下盆栽闭鞘姜Tr日均值分别为3.53、3.28、3.72、2.99、3.50 和 3.70 mmol/m2·s。
2.5 不同基质下盆栽闭鞘姜叶片净光合速率、蒸腾速率与环境因子的相关性分析
对净光合速率与环境因子相关分析结果(表1)表明,栽培在基质S1、S2、S4的闭鞘姜叶片Pn与光合有效辐射分别呈显著正相关和极显著正相关;此外,基质S1处理的盆栽闭鞘姜叶片的Pn与相对湿度呈极显著正相关。
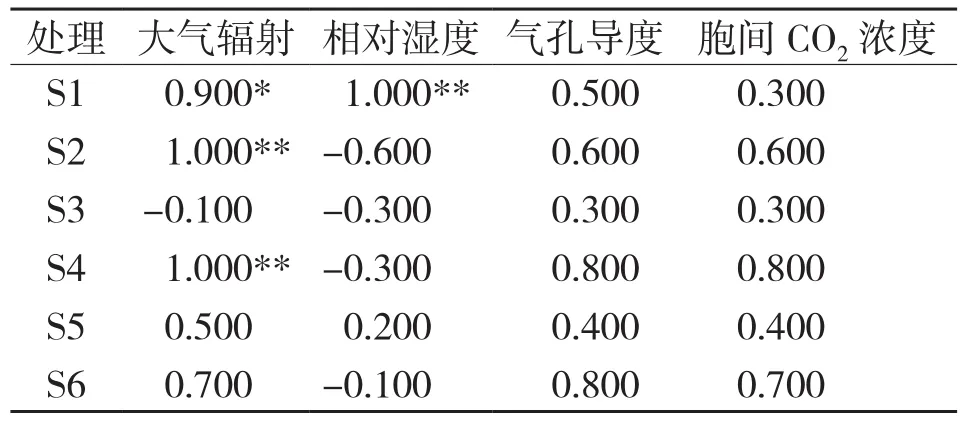
表1 不同基质下盆栽闭鞘姜叶片净光合速率与环境因子的相关系数
从蒸腾速率与环境因子相关分析结果(表2)可以看出,栽培在基质S4和S5的闭鞘姜叶片蒸腾速率与光合有效辐射PAR呈极显著正相关;栽培在S5的盆栽闭鞘姜叶片蒸腾速率与气孔导度和胞间CO2浓度分别呈显著正相关。
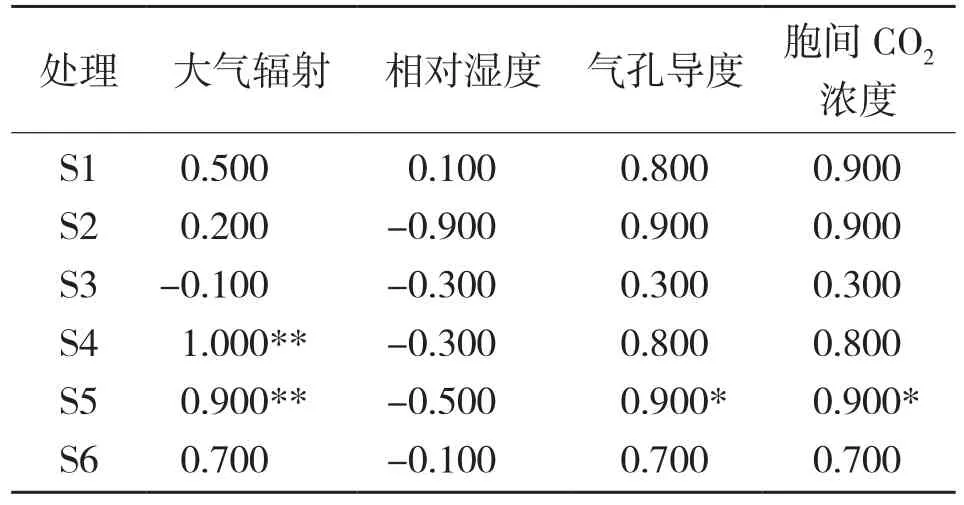
表2 不同基质下盆栽闭鞘姜叶片蒸腾速率与环境因子的相关系数
2.6 不同基质对盆栽闭鞘姜生长发育的影响
从表3可以看出,6种基质中闭鞘姜的株高存在显著性差异,其中S4处理的盆栽闭鞘姜株高显著高于其他基质,基质S3的株高次之;S4和S3处理的盆栽闭鞘姜茎基部粗显著高于其他基质;鲜重以及干重存在显著差异,S4的鲜重以及干重显著大于其他基质,S4的块茎鲜重比S1高58.9%,S4块茎干重比S6高115.2%。可见,S4较其他基质更适合闭鞘姜盆栽,该结果可为生产提供一定的理论依据。
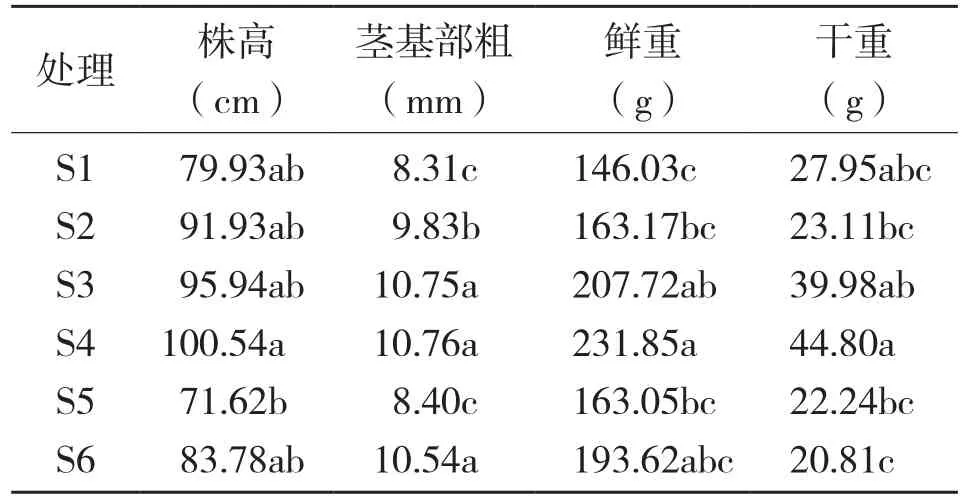
表3 不同基质对盆栽闭鞘姜生长发育的影响
3 结论与讨论
植物的光合特性与其生长环境密不可分,在生长过程中,植物逐渐形成了适应自身生长环境的光合特性,以保证其正常生长、发育、繁衍[9]。栽培基质是盆栽植物生长的介质,不同基质对花卉生长影响不同[10-11]。盆栽植物基质的物理性质容重、孔隙度影响植物根系的发育,进而影响其对营养元素(如N、P)的吸收[12],而N、P对光合作用影响显著。
在温度和光照辐射相同的天气及自然条件下,不同基质下植物的净光合特性日变化差异较大,植物光合作用的日变化曲线通常呈双峰型或单峰型[13],闭鞘姜6种不同基质盆栽光合特性研究结果显示:基质S1处理下的闭鞘姜的净光合速率日变化呈双峰型,其余5个基质呈单峰型曲线,这与张文斌等[14-15]研究结果相似。蒸腾作用是植物对水分吸收和运输的主要动力,陈兆波等[16]指出,香紫苏叶片的蒸腾速率变化是先升高后降低。本研究中,不同基质栽培的闭鞘姜叶片蒸腾速率变化的总体趋势可分为两大类:(1)基质S1、S3呈下降-上升-下降趋势;(2)基质S2、S4、S5和S6则呈上升-下降趋势。
在供试的6种基质中,盆栽闭鞘姜的净光合速率日均值分别为7.77、8.50、9.43、12.16、9.71和9.00 μmol/m2·s;蒸腾速率日均值分别为 3.53、3.28、3.72、2.99、3.50和 3.70 mmol/m2·s,可见不同基质对闭鞘姜的净光合速率和蒸腾速率影响不同,这与在其他作物[17-20]上的研究结果一致,其中S4处理下闭鞘姜的净光合速率与蒸腾速率日均值较其他基质高,这与盆栽基质的孔隙度和持水性有关,直接影响植物的生长发育与光合作用。环境因子PAR、相对湿度、气孔导度和胞间CO2浓度对不同基质中闭鞘姜叶片的净光合速率、蒸腾速率影响显著不同。基质S4种植出的植株在鲜重、干重、叶片数、茎基部粗细度、株高方面都优于其他基质栽培的植株。下一步我们将就不同基质的理化性质与叶片光合特性的关联性进行深入研究。
本研究结果表明,基质S4(泥炭∶椰糠∶珍珠岩=1∶2∶2)有利于提高盆栽闭鞘姜植株的净光合速率,增加单株株高和根状茎基部粗度,提高蒸腾作用和光合能力,从而有利于干物质累积,可应用于生产中。
[1]曾宋君,段俊. 姜目花卉[M]. 北京:中国林业出版社,2003.
[2]胡琳,陈娜娜,贺正山. 闭鞘姜化学成分及药理作用研究进展[J]. 云南民族大学学报(自然科学版),2014(1):18-23.
[3]苏新民,钟敏. 傣药材闭鞘姜药材质量研究[J]. 中国民族民间医药,2010(7):19-20.
[4]李忠琼,傅文,林瑞超,等. 闭鞘姜属植物化学成分及药理研究概况[J]. 中药材,2001(2):148-150.
[5]赵国祥,钱云,张光勇,等. 多功能植物闭鞘姜无公害人工栽培技术研究[J]. 热带农业科学,2009(4):43-46.
[6]曾凌云. 闭鞘姜及其人工栽培开发利用[J]. 蔬菜,2001(4):33-34.
[7]惠红霞,许兴,李前荣. 外源甜菜碱对盐胁迫下枸杞光合功能的改善[J]. 西北植物学报,2003,3(12):2137-2142.
[8]黄燕,吴平. SAS统计分析及应用[M]. 北京:机械工业出版社,2006.
[9]潘瑞炽. 植物生理学[M]. 第7版. 北京:高等教育出版社,2012.
[10]孙敏,姚海燕,陈伯清,等. 不同基质对几种花卉扦插生根及生长的影响[J]. 江苏农业科学,2009(1):179-181.
[11]张世超,陈少雄,彭彦. 无土栽培基质研究概况[J]. 桉树科技,2006(1):49-54.
[12]Gareia F,Cruse R M ,Blacker A M.Compaction and nitrogen placement efect on root growth,water depletion,and nitrogen uptake[J].Soil Science Society of America Journal,1998,52:792-798.
[13]邹琦. 作物抗旱生理生态研究[M]. 济南:山东科学技术出版社,1994.
[14]张文斌,许耀照,陈修斌. 不同茬口无土栽培基质水浸液对西葫芦幼苗生长及光合特性的影响[J]. 土壤与作物,2016(4):234-239.
[15]王岚,张宇斌,李建新. 不同栽培基质对七叶一枝花光合特性、生理特性的影响[J]. 江苏农业科学,2016(1):216-219.
[16]陈兆波,张翼,王沛,等. 香紫苏开花期蒸腾和光合作用日变化特征及其影响因子研究[J].西北植物学报,2007(6):1202-1208.
[17]徐德福,李映雪,郑建伟,等. 不同基质对黄菖蒲光合特性及净化能力的影响[J]. 环境科学,2011(9):2576-2581.
[18]许泽康,武华周,朱国鹏,等. 不同基质对鸟巢蕨生长和叶片生理特性的影响[J]. 热带作物学报,2015(1):41-46.
[19]陈凤真. 不同基质对小青菜穴盘苗生长和光合特性的影响[J]. 中国土壤与肥料,2014(1):75-78,100
[20]陈香,陆耀东,柯欢,等. 不同基质和生长调节剂对红桂木光合与水分利用效率的影响[J].亚热带植物科学,2011(1):5-8.
