分子标记辅助选育具抗稻瘟病Pi2基因的水稻新恢复系
2017-03-09余键吕德安郭承亮王世才
余键++吕德安++郭承亮++王世才
摘要:通过分子标记辅助选择,将早籼稻材料中的广谱抗稻瘟病基因Pi2通过杂交转入到中籼稻恢复系中,经过多代自交以及田间农艺性状的选择和自然诱发稻瘟病抗性鉴定,培育出了30个新的抗稻瘟病恢复系株系。通过对F6代抗性优良的株系与优良两系不育系C815S的大量杂交组合测配,选择出了3个高产、优质的新杂交组合,均比对照品种丰两优香1号增产5%以上,且对稻瘟病的抗性均优于对照品种,具有广阔的应用前景。
关键词:水稻(Oryza sativa L.);稻瘟病;恢复系;Pi2基因;分子标记辅助选择
中图分类号:S511;S336 文献标识码:A 文章编号:0439-8114(2017)01-0013-05
DOI:10.14088/j.cnki.issn0439-8114.2017.01.004
Developing New Restorer Lines with Pi2 Blast Resistance Gene by Molecular Marker Assistance Breeding in Rice
YU Jian1,2,L?譈 De-an1,2,GUO Cheng-liang1,2,WANG Shi-cai1,2,GENG Yue-ming1,2
(1.Hubei Provincial Seed Group Co. Ltd., Wuhan 430206, China;
2.Key Laboratory of India Rice Breeding and Seed Technology, Ministry of Agriculture, Wuhan 430206, China)
Abstract: In this research, we used molecular marker assistance breeding (MAS) to translate broad spectrum blast resistance gene Pi2 from early indica rice into restorer lines of medium indica rice. Through inbreeding of many generations,selection of good agronomic characters and identifying of the blast resistance under blast natural incidence,30 new restorer lines with blast resistance were cultivated. 3 new combinations with high yield, good quality were obtained from combinations between good two-line C815S and F6 excellent resistance restorer lines,and yields of these 3 combinations were higher than Fengliangyouxiang No.1 by more 5%,the blast resistance was also superior than control,which had extensive application prospect.
Key words: rice (Oryza sativa L.); blast; restorer lines; Pi2 Gene; molecular marker assistance breeding
水稻(Oryza sativa L.)稻瘟病是由子囊菌[Magnaporthe grisea (Hebert) Barrnov.]引起的真菌性病害,在全世界80多个国家和地区都有发生[1]。稻瘟病是限制水稻高产稳产的主要因素之一,给中国水稻的产量带来巨大的影响。20世纪90年代以来,中国稻瘟病的年发生面积在380万~400万hm2,造成稻谷年损失量达数亿千克,严重时高达10亿kg[2]。稻瘟病的变异性强,一般具有一定抗性的品种大面积推广3~5年后就会出现抗性下降或丧失,给生产带来极大的损失。长期的实践证明,发掘广谱、持久和稳定的抗稻瘟病新基因和培育新抗病性品種是亟待解决的问题,也是最经济有效的防治措施。
截至2013年8月,已至少报道了68个抗稻瘟病位点共83个主效基因。这些基因分布在除第3染色体以外的其他11条染色体上(2个隐性,其他显性)[3],其中第6、11、12号染色体上的抗性基因多且成簇分布[4],是3个最大的稻瘟病抗性基因簇。广谱抗稻瘟病基因Pi2位于第6染色体短臂近着丝粒附近[5],对从中国收集的792个稻瘟小种中的绝大部分表现抗性[6]。通过分子标记辅助选择已经在很多材料中得到应用[7-9]。本研究通过杂交将早籼稻材料Z219中的抗稻瘟病基因Pi2转入湖北省种子集团有限公司选育的7个品质优、配合力强的恢复系中,然后通过自交、分子标记辅助选择和田间自然诱发抗性鉴定,选择抗性好、农艺性状稳定的株系与不育系进行组合测配,拟在选育出含Pi2基因的新的抗稻瘟病优质恢复系材料。
1 材料与方法
1.1 植物材料与选育方法
供体亲本:早籼稻抗稻瘟病材料Z219,含有广谱抗稻瘟病的Pi2基因。受体亲本:优质中籼稻材料R201、R202、R203、R204、R205、R206和R207,都为湖北省种子集团有限公司自主选育。
从F2代开始结合分子标记辅助选择和表型鉴定,选择农艺性状优良、田间稻瘟病抗性良好的阳性单株,在F5~F6代进行稻瘟病自然诱发抗性鉴定、组合测配及组合抗性鉴定。
1.2 分子标记辅助选择
DNA的提取采用CTAB法;导入片段鉴定使用Pi2基因SSR标记RM527的序列合成引物。PCR反应体系总体积为20 μL,其中模板2 μL,去离子水12 μL,Buffer(10×)2 μL,Mg2+(25 mmol/L)1.8 μL,dNTP(5 mmol/L)1.8 μL,正反引物各0.2 μL,Taq酶0.2 μL。
扩增程序:95 ℃变性4 min;95 ℃变性30 s, 55 ℃退火30 s,72 ℃延伸45 s,35次循环;72 ℃延伸10 min。PCR扩增产物经聚丙烯酰胺凝胶(PAGE)电泳,银染检测。
1.3 稻瘟病田间抗性鉴定
稻瘟病田间抗性鉴定设于湖北省宜昌市远安县苟家垭镇望家乡以及恩施自治州白果乡两河村稻瘟病自然诱发鉴定圃,采用诱发品种和偏氮施肥及淹水保持高湿管理等措施辅助诱发。每份材料插2行,每行10株,单本插,株行距17 cm×17 cm。顺序排列,无重复。每组材料设抗、感品种和生产主栽对照品种各1个,供试材料及鉴定圃周围种植感病诱发品种。
叶瘟病于分蘖盛期对供试材料逐株调查发病病级,然后加权平均为发病等级;穗颈瘟病于收获前1周内分别调查发病率和损失率(统计穗数不少于100穗),分级标准参照湖北省水稻区域试验方法,即国家稻品种试验与审定规范《水稻抗主要病虫鉴定及评价标准试行方案》。
1.4 选种、组合测配的田间种植及考种
选种种植:每个小区种植20苗,双行区,每行10苗,单本插,密度16.7 cm×20.0 cm。记载生育期和整齐度。成熟时每个区选择1个以上的优良单株,收获中选株的全部种子。
组合测配种植:每个小区3次重复,30株,单本插,每行10苗,密度16.7 cm×20.0 cm,设丰两优香1号为中稻对照,成熟时混收每个小区的种子,测定小区产量,并取中间纵向的3株进行考种,考察其农艺性状。考察的农艺性状主要包括播始历期、株高、穗长、有效穗数、实粒数、空粒数、结实率、千粒重和小区产量等。
1.5 稻米品质分析
根据国家优质稻谷标准(GB/T 17891-1999)的测定方法,对配制的杂交组合以及2个对照品种进行稻米品质分析,包含的指标有糙米率、精米率、整精米率、长宽比、垩白粒率、垩白度、胶稠度和直链淀粉含量等8个性状。
2 结果与分析
2.1 Pi2基因的分子检测
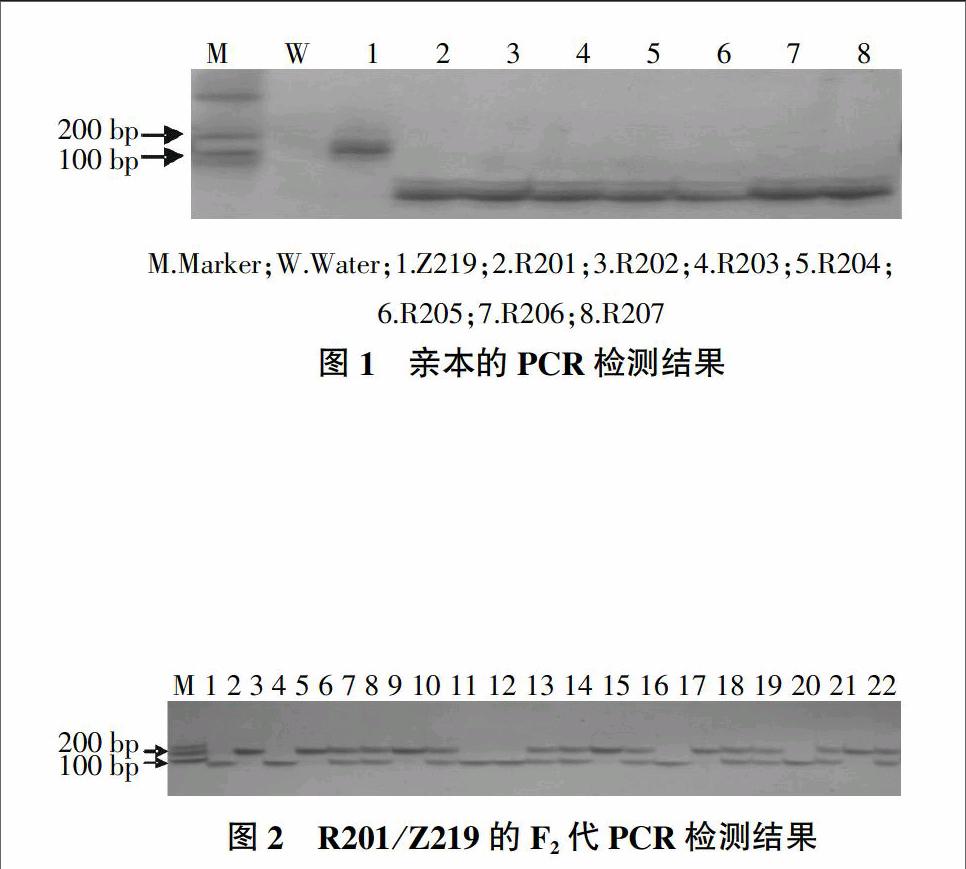
图1为Pi2基因亲本的分子检测结果,所用引物为共显性标记RM527。由图1可知,供体亲本和受体亲本之间有显著的多态性,可以用于Pi2基因的多态性检验。
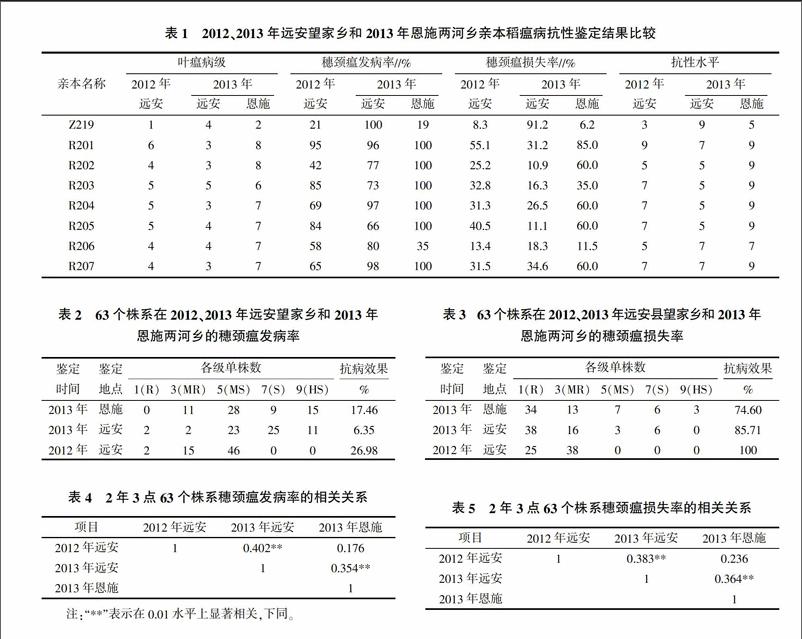
图2为R201/Z219的自交F2代分子标记检测结果,可以看出有明显的分离现象,而且3种带型符合1∶2∶1的孟德尔遗传规律,说明该引物可以对阳性纯合、杂合和阴性纯合个体进行准确的筛选。
2.2 亲本及育成株系的稻瘟病抗性
2.2.1 亲本的稻瘟病抗性 2012年和2013年在湖北省宜昌市远安县望家乡、2013年在恩施两河乡对8个亲本株系进行了2年3点的稻瘟病田间自然诱发抗性鉴定。由表1可知,供体亲本Z219的叶瘟病病级、穗颈瘟发病率和穗颈瘟损失率在2012年远安和2013年恩施点的鉴定结果相同,达到R-MS-MR级别,都与2013年远安的鉴定结果MR-HS-HS级别有显著差异。其他受体亲本除叶瘟病级在3点都有差异外,穗颈瘟发病率和穗颈瘟损失率与供体亲本的鉴定结果有一致的差异性。因此,认为2012年远安和2013年恩施的鉴定结果可靠,而且以穗颈瘟发病率和穗颈瘟损失率作为抗性鉴定的标准。
2.2.2 育成株系的稻瘟病抗性 通过2012年在远安县望家乡对育成含Pi2基因的自交F5代恢复系株系进行自然诱发稻瘟病抗性的鉴定,共筛选了19个综合抗性为1级、44个3级的抗性株系。2013年在远安县望家乡对这63份稻瘟病综合抗性3级及以上自交F6代恢复系株系进行重复抗性鉴定,同时在恩施两河乡进行异点的鉴定。
由表2穗颈瘟病发病率的抗病效果可知,2013年远安和恩施的抗病效果比2012年远安有明显下降,其中恩施下降了9.52个百分点,远安下降了20.63个百分点。2013年恩施出现了感(S)和高感(HS)的株系,占鉴定株系数的38.10%;2013年远安出现的感(S)和高感(HS)的株系数达57.14%。2年3点的穗颈瘟发病率的抗病效果都低于30%,可能與供体亲本的穗颈瘟发病率平均在感级别(S),受体亲本在高感级别(HS)有关,也可能是由于部分株系未完全纯合,从而出现分离,导致抗性差异。
表3为穗颈瘟损失率的抗病效果。由表3可知,2013年远安和恩施的穗颈瘟损失率的抗病效果都比2012年远安有明显的降低,其中远安下降了14.29个百分点,恩施下降了25.40个百分点。2013年远安抗穗颈瘟病效果下降和出现感病株系可能与第二次重复鉴定有关,2013年恩施鉴定结果与2012年远安的差异可能与两地自然条件不同有关。
通过表4和表5的穗颈瘟发病率和穗颈瘟损失率之间的相关关系可知,2012年远安和2013年远安、2013年远安和2013年恩施的穗颈瘟发病率之间和穗颈瘟损失率之间存在极显著的相关关系,2012远安和2013恩施的穗颈瘟发病率之间和穗颈瘟损失率之间没有相关性。这可能与2012年和2013年远安为同地点,环境相同,2013年恩施和远安为同年,气候相差不大有关;而2013年恩施和2012年远安环境和气候都不相同。
根据对2年3点的气候和抗性结果分析,认为2012年远安和2013年恩施的鉴定结果是比较可靠的。2013年远安鉴定田,在水稻成熟期出现低温,可能对鉴定结果的准确性具有一定的影响。表6是综合分析2012年远安和2013年恩施的鉴定结果,选出的抗性较好的株系。这30个株系的稻瘟病抗性明显比原来的中稻亲本有所提高,在今后杂交组合选育中,可能有较好的应用价值。
2.3 育成株系所配杂交组合的稻瘟病抗性鉴定结果
于2013年在远安县望家乡对2012年综合抗性级别达到1(R)级的19个恢复系株系与C815S所配的杂交组合进行田间自然诱发稻瘟病抗性鉴定。表7为所配组合抗性鉴定穗颈瘟发病率和穗颈瘟损失率的级别。由表7可知,各组合的穗颈瘟发病率绝大部分都达到了高感(HS);各组合的穗颈瘟损失率大部分都为中感(MS),少部分为中抗(MR),都明显低于亲本恢复系株系的抗性。这可能与杂交组合的抗性是由父母本共同的基因型决定有关,由于母本不具有稻瘟病抗性,甚至高感稻瘟病,从而导致了杂交组合的抗性显著下降。但与不育系C815S所配组合的穗颈瘟损失率级别都优于对照丰两优香1号,绝大部分都在中感(MS)及以上,说明这些组合具有一定的穗颈瘟损失率抗性。
2.4 育成株系所配增产杂交组合的生育期、产量和主要农艺性状表现
表8为4个高产中稻杂交组合的生育期、产量和主要农艺性状表现。由表8可知,播始历期4个杂交组合都长于或与对照相当;株高都低于对照;单株有效穗数都高于对照;平均穗长都短于对照;每穗总粒数除C815S/Q191-3低于对照外,其他杂交组合都高于对照;结实率和千粒重所有杂交组合都低于对照。杂交组合C815S/Q221-2-2与对照丰两优香1号产量相当。
2.5 育成株系所配增产杂交组合稻米品质表现
对由C815S所配的中稻杂交组合进行稻米品质分析,结果(表9)表明,增产的4个中稻杂交组合中,糙米率除C815S/Q213-2为国标2级外,其他3个组合均为国标1级,与对照丰两优香1号相当;整精米率均低于对照,仅C815S/Q191-3达国标3级水平;垩白粒率和垩白度均高于对照;粒型所有杂交组合均与对照相当;直链淀粉含量仅C815S/Q213-1达国标3级,其他3个组合均与对照相当;胶稠度均与对照相当,达国标1级水平。
3 讨论
本研究旨在创建抗稻瘟病的新恢复系,但是并没有选择双亲都具有抗性的亲本进行杂交,从而聚合抗稻瘟病基因,获得抗性更好的株系这种途径,而是通过含有广谱抗稻瘟病基因Pi2的早籼稻材料Z219与不含抗稻瘟病基因的优良恢复系杂交,通过分子标记辅助选择含有Pi2基因的株系自交,最后获得稳定的新的抗病恢复系。这样做的目的有两点:一是为了育种的需要,不需要改良某个特定的恢复系的抗性,而是创建新的恢复系,为育种提供新材料;二是育成的新株系可以直接用于组合测配,加快了抗病杂交育种的进程。
本研究育成的新株系分为3种类型:第一种是含有Pi2基因,但稻瘟病抗性却差于供体亲本Z219,可能原因是由于基因没完全纯合或基因的表达出现问题;第二种是含有Pi2基因,抗性与供体亲本相当,可能是Pi2基因的结构没有发生改变;第三种是含有Pi2基因,抗性优于供体亲本,可能是Pi2基因纯合,然后经过染色体的交换或重组,获得新的未知抗稻瘟病基因或QTL位点。具体原因还需进一步验证。
王久林等[10]认为杂种的稻瘟病抗性与不育系和恢复系都有关,且偏向于恢复系。郑轶等[11]认为杂交组合的叶瘟病和穗颈瘟病抗性与母本叶瘟病均呈极显著正相关关系,而与父本的相关性不显著,母本的抗性越好,则配制的杂交组合抗性更好。本研究通过对C815S与育成的稻瘟病综合抗性1级的恢复系配制的杂交组合进行自然诱发的稻瘟病抗性鉴定,结果表明杂交组合叶瘟病抗性处于抗-中抗级别,穗颈瘟发病率为感或高感级别,穗瘟损失率为中抗-中感级别,综合抗性中抗-中感级别,优于对照丰两优香1号(感)。
水稻叶瘟病与穗颈瘟病之间的关系一直存有争论,一种认为它们之间存在着正相关关系[12-14],另一种认为互相之间没有相关性,甚至是由不同的基因决定。本研究分析了2012年远安叶瘟发病率和穗颈瘟发病率之间的相关关系,结果表明叶瘟发病率和穗瘟发病率之间存在极显著的正相关关系。但是本试验中大部分叶瘟病鉴定表现为抗病级别的株系,但穗颈瘟发病率却表现为感病级别,即李桦[12]所认为的“R,S”型,这是因为新育成的株系抽穗不整齐且呈分离特点导致的。因此,需要对育成的株系进一步选择、纯化,以提高新育成株系整体的稻瘟病抗性水平。
本研究对两年同点和同年不同点的鉴定结果之间进行了相关性分析,结果表明,两年同点的叶瘟病级别之间以及与其他的抗性之间没有显著的相关关系,同年不同点的结果与两年同点的结果相似,说明叶瘟病在两年同点和同年不同点之间都不具有重复性;而穗颈瘟发病率与损失率在同年不同点及两年同点间都有显著的相关关系,所以在品种和新株系的稻瘟病抗性鉴定时,应以穗颈瘟发病率和损失率作为鉴定的标准,提高鉴定的准确性。
4 结论
本研究通过分子标记辅助选择将抗稻瘟病Pi2基因转入自育的恢复系中,育成了一批新的抗稻瘟病恢复系,通过2年3点的自然诱发田间抗性鉴定,获得了30个稻瘟病抗性明显优于原始受体亲本的新抗病恢复系,与不育系C815S测配杂交组合的抗性鉴定表明,杂交组合的稻瘟病抗性提升不明显,但要优于对照品种丰两优香1号,且产量、米质方面都比对照有所提高,具有一定的应用前景。
参考文献:
[1] 陈德西,李仕贵,马炳田,等.水稻抗稻瘟病育种研究进展[J].安徽农学通报,2008,14(17):206-209.
[2] 李落叶,井金学.稻瘟病抗性基因的分子定位及克隆[J].中国农学通报,2006,22(1):49-53.
[3] 谢 培,邓其明,王世全,等.水稻稻瘟病抗性基因研究进展[J].湖南农业科学,2011(5):79-81,82.
[4] BALLINI E,MOREL J B,DROC G,et al. A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance[J].MPMI,2008,21:859-868.
[5] ZHOU B,QU S,LIU G,et al. The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea[J].MPMI,2006,19:1216-1228.
[6] CHEN H L,CHEN B T,ZHANG D P,et al.Pathotypes of Pyricularia grisea in rice fields of central and southern China[J].Plant Disease,2001,85:843-850.
[7] 陈红旗.分子标记辅助聚合3个稻瘟病抗性基因[D].江苏扬州:扬州大学,2005.
[8] 郭艳丽.利用MAS技术培育寒区抗稻瘟病水稻品种五优稻1号(Pi1/Pi2)[D].哈尔滨:黑龙江图书馆,2009.
[9] 林 琳.稻瘟病抗性基因的多基因聚合研究[D].成都:四川农业大学,2010.
[10] 王久林,雷财林,毛世宏,等.杂交稻抗瘟性遗传表达特征的研究[J].福建稻麦科技,1995,13(2):12-17.
[11] 鄭 轶,涂诗航,周 鹏,等.杂交水稻稻瘟病抗性鉴定及其与父母本抗性的相关性[J].杂交水稻,2010,25(4):75-77.
[12] 李 桦.水稻稻瘟病苗瘟、叶瘟、穗颈瘟相关关系的探讨[J].植物保护学报,1991,18(4):293-297.
[13] 李 华.宁夏水稻抗稻瘟病育种存在的问题及鉴定方法的探讨[J].宁夏农林科技,1997(4):36-38.
[14] 陈福如,阮宏椿,杨秀娟,等.稻瘟病苗瘟叶瘟和穗颈瘟的相关性分析[J].中国农学通报,2006,22(7):440-443.
