紫茎泽兰在不同群落中的生长与繁殖特征
2017-03-09韦春强唐赛春潘玉梅李象钦吕仕洪广西壮族自治区中国科学院广西植物研究所广西桂林541006
韦春强, 唐赛春, 潘玉梅, 李象钦, 吕仕洪(广西壮族自治区中国科学院广西植物研究所,广西桂林 541006)
紫茎泽兰(Ageratinaadenophora)为菊科(Compositae)泽兰属(Eupatorium)多年生草本或半灌木状植物,是一种危害严重的世界性恶性杂草,原产于中、南美洲,现已扩散到亚、非、欧、美、澳等各大洲的30多个国家和地区[1-2]。紫茎泽兰约于20世纪40年代由中缅边境传入云南省,经过半个多世纪的传播扩散,现已广泛分布在我国西南地区的云南、贵州、广西、四川、重庆、西藏等省(市、区),并以较快的速度继续向东和向北传播蔓延,给当地的农业、畜牧业和生态环境造成严重的危害[3-6]。在2003年时紫茎泽兰已被列入国家环境保护总局和中国科学院联合发布的我国首批外来入侵物种名录中,且排在第1位[7]。
目前对紫茎泽兰的生物学特性、入侵机制、入侵的生态学效应、防治与利用等方面都有了较多的研究[8-13]。对其在不同环境下生长与繁殖的研究也有少量报道,赵相健等的研究发现,不同环境下紫茎泽兰生长和繁殖特征均存在显著差异[14-15]。然而目前紫茎泽兰已入侵了各种各样的生境,并造成了严重的危害,生境的多样化也就须要更多的研究来解析生境变化给紫茎泽兰带来的影响,同时研究不同群落中紫茎泽兰的生长和繁殖变化,有助于从生态角度提出更为科学的防治紫茎泽兰的方法。目前对紫茎泽兰的研究多集中在云南、贵州和四川等省份[16-19],而在广西开展的研究较少[20]。因此,本试验通过对广西境内最常见的且具有代表性的紫茎泽兰入侵地群落进行调查,研究紫茎泽兰在不同生境条件下的生态对策,分析这些特性与其入侵性的关系,为深入研究紫茎泽兰入侵、扩散机制以及制定有效的防控措施提供更充足的科学依据。
1 材料与方法
1.1 研究地点概况
研究地点位于广西隆林县德峨乡(105°7′55″E、24°39′5″N),海拔1 521 m,距县城36 km,是隆林南部乡镇以及滇、黔、桂3省(区)的重要交通枢纽。属云贵高原余峰,日照时数 1 384 h,日最高温度24 ℃,平均气温14.42 ℃,年均降水量约为1 500 mm,为多雾阴雨高寒山区。
1.2 方法
于2017年4月在紫茎泽兰种子成熟季节选择广西境内最常见的且具有代表性的紫茎泽兰入侵地群落进行研究,群落1,开阔地,紫茎泽兰单优势群落,紫茎泽兰盖度为80%~95%;群落2,桉树(Eucalyptusspp.)林,郁闭度0.3左右,属于中度郁闭,紫茎泽兰盖度为65%~85%;群落3,马尾松(Pinusmassoniana)林,郁闭度0.6左右,属于中度郁闭,紫茎泽兰盖度为30%~40%;群落4,白栎(Quercusfabri)、黄檀(Dalbergiahupeana)混交林,郁闭度0.8左右,属于高度郁闭的密林,紫茎泽兰盖度为10%~25%。以随机样方法在每个群落中随机设置5个2 m×2 m的样方,测量各样方内紫茎泽兰株高、总分蘖数、总头状花序花数、各小花内所含种子数量(每样方选测5朵小花)、地上部分植株鲜质量(野外全株称量)、总花序鲜质量、单分蘖地上部分湿质量以及该单分蘖上头状花序鲜质量(野外称量鲜质量后带回实验室并测其干质量,每个样方选测4株分蘖)。
1.3 数据分析
用Excel软件对数据进行整理、计算并绘图,以平均数代表样本各性状的整体水平,以标准差反映样本的绝对变异度。采用SPSS 13.0统计分析软件对紫茎泽兰各参数进行描述性分析及单因素方差分析(One-Way ANOVA)与多重比较,Pearson相关系数法进行相关性分析。
2 结果与分析
2.1 不同群落中紫茎泽兰的营养生长比较
由图1可知,群落2中的紫茎泽兰株高显著高于其他群落,群落4中的紫茎泽兰株高显著低于其他群落,而群落1与群落3无显著差异。群落1和群落2中紫茎泽兰单位面积的分蘖数无显著差异,但显著多于群落3和群落4。4个群落中紫茎泽兰的单分蘖地上生物量大小依次为:群落1>群落2>群落3>群落4。
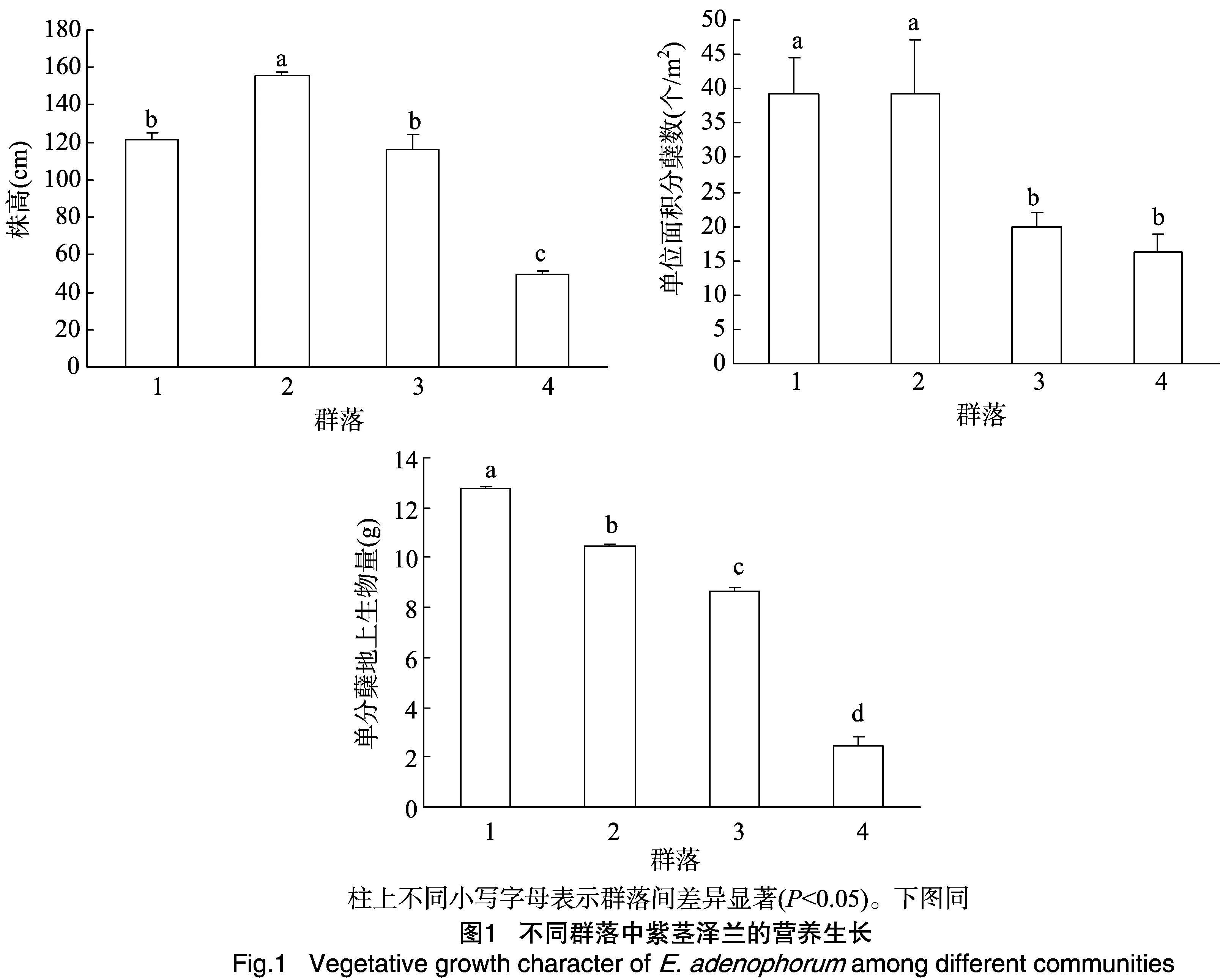
2.2 不同群落中紫茎泽兰的繁殖特征比较
由图2可知,群落中紫茎泽兰的单分蘖小花数、单花种子数均表现为群落1最高,群落2和群落3次之,群落4最低。单分蘖的小花数和单花种子数决定单面积种子产量,因此各群落中紫茎泽兰的单面积种子产量变化趋势与单分蘖的小花数和单花种子数一样。
2.3 不同群落中紫茎泽兰地上组织的生物量积累与繁殖分配比较
由图3可知,不同群落对紫茎泽兰的生物量积累有不同影响。群落1和群落2的单位面积地上生物量显著高于群落3和群落4,且群落4的单位面积地上生物量最少,表明郁闭度较高的密林不利于紫茎泽兰的生长。各群落间紫茎泽兰单位面积花生物量存在显著差异,且随郁闭度的升高而降低。群落1中紫茎泽兰的繁殖分配显著大于其他群落,主要是由于群落1为开阔地,受到的限制因素较少,紫茎泽兰可以将更多的资源分配到繁殖器官,以利于其传播扩散。
2.4 郁闭度与紫茎泽兰生长繁殖特性的相关性
从表1可以看出,群落的郁闭度与紫茎泽兰的株高、单位面积分蘖数、单分蘖地上生物量、单分蘖小花数、单花种子数、单位面积种子产量、单位面积地上生物量、单位面积花生物量以及繁殖分配都呈显著的负相关关系,说明群落郁闭度影响紫茎泽兰的生长与繁殖。
3 讨论
自然界中不同立地条件下的微环境变化较大,对植物的生长也产生一定的影响[21]。处于不同立地条件下的植物,会在存活、生长、繁殖之间作出权衡,形成一定的生活对策特征,以适应环境的变化[22]。因此,在异质生境中,植物种群的营养生长和繁殖特征往往表现出明显差异[23]。赵相健等对云南省9个不同地理种群紫茎泽兰的野外生长繁殖特性进行调查发现,处于不同条件下野外不同地理种群的紫茎泽兰之间的植株生长和繁殖特征均存在显著差异[14]。徐高峰等在薇甘菊(Mikaniamicrantha)常见的4种入侵生境中进行调查研究发现,薇甘菊对异质环境条件具有不同的生长与繁殖适应对策[24]。



表1 郁闭度与紫茎泽兰生长繁殖特性的相关性Table 1 Correlation between canopy density and growth and reproduction characters of E. adenophorum
注:***表示在0.01水平上显著。
本研究调查结果表明,紫茎泽兰在不同群落中的生长与繁殖同样表现出明显差异。在开阔地,紫茎泽兰将更多的资源投入到繁殖中,增加其繁殖分配比例,以便于种群的扩散传播;而在林地中,环境选择压力较大,使得紫茎泽兰在生存与繁殖之间更倾向于选择生存,减少对繁殖的投入,因而单分蘖头状花序数、单花种子数以及繁殖分配比例都相应减少,而将更多的资源投入营养生长中,增强其个体的存活能力,以适应不利的环境。在高郁闭度的林地中,紫茎泽兰不仅生殖生长受到严重影响,营养生长也受到严重影响,但是仍能存活下来,并且产生少量花和种子,说明紫茎泽兰具有很强的适应能力,在条件有利时,可产生大量的种子,快速占领生境,而在条件不利时,则维持其基本的生存和繁殖,等待有利条件的出现,这是紫茎泽兰能快速入侵的策略之一。
从相关性分析结果来看,紫茎泽兰的生长和繁殖与群落的郁闭度有很大的相关性,也就是说光线强度对紫茎泽兰的生长和繁殖有很大的影响,许多研究也证实光照度显著影响紫茎泽兰的生长和繁殖[15,25-28]。在郁闭度越高的群落中,紫茎泽兰的生长越差,郁闭度较高的群落对紫茎泽兰有较好的控制作用,因而可以通过进行群落改造来增加群落的郁闭度,进而有效地控制紫茎泽兰生长和扩散。
[1]强 胜. 世界性恶性杂草——紫茎泽兰研究的历史及现状[J]. 武汉植物学研究,1998,16(4):366-372.
[2]Auld B A. The distribution ofEupatoriumadenophorumSpreng. on the far north coast of New South Wales[J]. Journal and Proceedings of the Royal Society of New South Wales,1969,102:159-161.
[3]肖正清,周冠华,权文婷. 恶性外来入侵植物紫茎泽兰在云南的分布格局[J]. 自然灾害学报,2009,18(5):82-87.
[4]卢志军,马克平. 地形因素对外来入侵种紫茎泽兰的影响[J]. 植物生态学报,2004,28(6):761-767.
[5]刘伦辉,谢寿昌,张建华. 紫茎泽兰在我国的分布、危害与防除途径的探讨[J]. 生态学报,1985,5(1):1-6.
[6]Lu Z J,Ma K P. Spread of the exotic crofton weed (Eupatoriumadenophorum) across southwest China along roads and streams[J]. Weed Science,2006,54(6):1068-1072.
[7]国家环境保护总局,中国科学院.中国外来入侵物种名单:第一批[EB/OL]. (2003-01-10)[2017-05-04]. http://www.zhb.gov.cn/download/ 1055369430488.doc.
[8]万方浩,刘万学,郭建英,等. 外来植物紫茎泽兰的入侵机理与控制策略研究进展[J]. 中国科学(生命科学),2011,41(1):13-21.
[9]李霞霞,张钦弟,朱珣之. 近十年入侵植物紫茎泽兰研究进展[J]. 草业科学,2017,34(2):283-292.
[10]于兴军. 紫茎泽兰入侵生态学研究[D]. 武汉:武汉大学,2005.
[11]王崇云,潘 燕,朱晓媛,等. 紫茎泽兰在弃耕地上的种子风传播与种群建立特征[J]. 杂草科学,2015,33(3):1-5.
[12]鲁 萍,桑卫国,马克平. 外来入侵种紫茎泽兰研究进展与展望[J]. 植物生态学报,2005,29(6):1029-1037.
[13]卢向阳,曹坳程,欧阳灿彬,等. 葛藤定植及替代控制紫茎泽兰的技术研究[J]. 杂草科学,2015,33(4):18-22.
[14]赵相健,刘文耀,周 蒙,等. 不同地理种群紫茎泽兰生长繁殖特征的比较研究[J]. 西北植物学报,2009,29(6):1252-1258.
[15]朱文达,曹坳程,颜冬冬,等. 不同林木种群对紫茎泽兰营养生长和生殖生长的影响[J]. 生态环境学报,2013,22(11):1790-1794.
[16]周 蒙,刘文耀,马文章,等. 不同地理种源紫茎泽兰的生态适应性比较[J]. 应用生态学报,2009,20(7):1643-1649.
[17]苏秀红,强 胜,宋小玲. 不同地理种群紫茎泽兰耐热性差异的比较分析[J]. 西北植物学报,2005,25(9):1766-1771.
[18]黄梅芬,徐 驰,曹后英,等. 不同生境条件对紫茎泽兰营养生长的影响[J]. 热带作物学报,2009,30(10):1429-1436.
[19]欧国腾,赵宇翔,江 赢,等. 贵州南部山区紫茎泽兰的替代控制研究[J]. 中国森林病虫,2012,31(2):23-26.
[20]贾桂康. 外来入侵种紫茎泽兰在广西的分布与危害[J]. 百色学院学报,2007,20(3):90-95.
[21]山 仑,徐炳成,杜 峰,等. 陕北地区不同类型植物生产力及生态适应性研究[J]. 水土保持通报,2004,24(1):1-7.
[22]董 力. 不同生境条件下头花蓼的生长繁殖特征研究[D]. 重庆:西南大学,2011.
[23]张治来,拓 飞,尚 华,等. 不同立地类型大果沙棘生长及生态适应对策差异[J]. 西南林学院学报,2009,29(5):12-15.
[24]徐高峰,申时才,张付斗. 异质环境下入侵植物薇甘菊的适应性与繁殖特性[J]. 生态环境学报,2014,23(8):1258-1264.
[25]王俊峰,冯玉龙,梁红柱. 紫茎泽兰光合特性对生长环境光强的适应[J]. 应用生态学报,2004,15(8):1373-1377.
[26]刘伦辉,刘文耀,郑 征,等. 紫茎泽兰个体生物及生态学特性研究[J]. 生态学报,1989,9(1):66-70.
[27]孙锡治,全骏纯,饶维维,等. 紫茎泽兰个体生物学特性与防除技术[J]. 云南农业科技,1992,(4):13-15.
[28]Cabin R J,Weller S G,Lorence D H,et al. Effects of light,alien grass,and native species additions on Hawaiian dry forest restoration[J]. Ecological Applications,2002,12(6):1595-1610.
