山核桃天牛危害调查及种类的鉴别方法
2017-02-22张冬勇周书永樊建庭
张冬勇,周书永,朱 宁,樊建庭
(1.浙江农林大学 林业与生物技术学院,浙江 临安 311300;2.浙江省临海市森林病虫防治检疫站,浙江 临海317000)
山核桃天牛危害调查及种类的鉴别方法
张冬勇1,周书永2,朱 宁1,樊建庭1
(1.浙江农林大学 林业与生物技术学院,浙江 临安 311300;2.浙江省临海市森林病虫防治检疫站,浙江 临海317000)
为了掌握危害山核桃Carya cathayensis的天牛种类以及危害规律,对浙江省临安地区的山核桃林地进行了系统调查,研究了天牛危害种类、危害状以及不同种类的识别方法。结果表明:临安地区山核桃天牛种类主要包括桑天牛Apriona germari,星天牛Anoplophora chinensis和云斑天牛Batocera horsfieldi等3种;其中桑天牛幼虫分布在山核桃枝条部位(100%),星天牛幼虫分布在根部和地上树干30 cm以下(100%),云斑天牛幼虫分布在树干0~ 150 cm(93.85%);不同种类天牛的幼虫排粪状、幼虫蛀道和成虫产卵刻槽具有不同的典型症状;桑天牛、星天牛和云斑天牛雌雄成虫之间具有显著的形态特征,可以用来区分雌雄个体。另外,3种天牛幼虫的前胸背板具有不同的典型特征,可以通过此特征快速准确区分其幼虫种类,相应的分子鉴定结果也证明了此方法的准确性。由此推测:云斑天牛和星天牛主要危害成年山核桃,而桑天牛主要危害幼龄山核桃,而且不同种类天牛幼虫在山核桃树上占据不同的生态位。不同种类天牛危害症状和典型形态特征的研究,对于指导山核桃天牛害虫防控工作也具有重要的意义。图5表5参19
森林保护学;山核桃;天牛;调查;幼虫;鉴别
山核桃Carya cathayensis又称昌化山核桃,属胡桃科Juglandaceae山核桃属Carya,是中国特有的名优干果和木本油料作物,具有营养、保健、美容及药用价值[1],国家三级保护濒危植物[2],主产于浙、皖交界的天目山区,29°~30°N,118°~120°E。90%以上的山核桃生长于石灰岩土壤上[3],其中浙江省临安地区的山核桃林地面积达3.1万hm2,产量占全国的50%左右[4],是当地林农增收、脱贫致富的主要经济来源。山核桃产业的快速稳定健康发展对促进区域发展具有重要意义。受极端气候事件增多、经营强度加大和化肥使用量上升等因素的影响,山核桃林地有害生物的种类、分布区域、发生程度和危害情况均发生了重大变化,病虫害暴发频率逐年提高,发生范围不断扩大,经济损失也不断增加[5-6]。其中,山核桃天牛Cerambycidae危害日趋严重,主要以天牛幼虫蛀食寄主木质部进行危害,具有隐蔽性强,危害严重的特点,被称为 “心腹之患”,轻则造成风折木,重则造成整株枯死,成为限制山核桃产量的一个重要因素。山核桃寄主上同时有多种天牛混合发生,各自危害规律也不相同,至今还没有对山核桃天牛种类及其危害规律的系统调查。众所周知,天牛成虫的形态特征较为明显,容易辨识种类,但是天牛幼虫历期相对其他虫态来说更长,而且幼虫是主要危害虫态,因此,天牛幼虫的识别既是实际生产应用中的重点也是难点,如何通过幼虫危害状或者典型的形态特征快速准确地识别山核桃天牛种类,对于防控山核桃天牛这类害虫具有重要的指导意义。本研究对山核桃林地天牛种类和危害规律进行了系统调查,并且研究了天牛幼虫危害状和形态特征的简易鉴别方法。
1 研究方法
1.1 试验地概况
调查地点选择在浙江省临安市(29°56′~30°23′N,118°51′~119°52′E)的湍口镇、太阳镇和校园试验地,分别设置3个试验样地。临安地处浙江省西北部,属于季风气候,年均降水量为1 613.9 mm,降水日158.0 d,无霜期年平均为237.0 d,年平均气温由16.0℃降至9.0℃,年温差7.0℃,横跨亚热带和温带2个气候带。太阳镇和湍口镇的山核桃树均栽植在山坡上,湍口镇山核桃林下植被主要有桂花Osmanthus fragrans,油茶Camellia oleifera,南天竹Nandina domestica和黑麦草Lolium perenne等地被植物,林地面积120 hm2,山核桃树龄为20~30 a;太阳镇山核桃林下只有零星的草本植物,无其他藤本灌木植物,山核桃树龄为20~30 a,郁闭度0.7;校园试验地山核桃林下植物稀少,山核桃平均树龄为7~8 a。各地山核桃树都受到不同程度的山核桃天牛危害。
1.2 调查方法
在2015年5月到6月期间,在3个试验样地中,随机调查30株·样地-1有天牛危害的山核桃树,进行解剖调查,记录天牛幼虫采集部位(如枝条、树干、根部),采集高度(cm)等信息,同时用佳能相机(机身700D,EFS18~135 mm镜头)拍照记录不同种类天牛的形态特征和危害特征,活体天牛幼虫当即浸泡在体积分数为96%乙醇中保存,放置于实验室-20℃冰箱保存待用。
生态位宽度按照LEVINS提出的公式计算[7]。①生态位宽度其中:B为生态位宽度;S为生态位资源等级数;Pi为在一个资源集合中,物种在第i单元中所占的比例。②生态位重叠[8-10]aij=其中:aij=第i个物种对物种j的生态位重叠;Pih,Pjh分别是第i第j个物种在资源集第h个单元中的比例。Bi为第i种的生态位宽度。
1.3 天牛幼虫前胸背板特征
将浸泡标本取出,用吸水纸吸干虫体表面,置于电子显微镜(Nikon AZ100)下仔细观察其前胸背板的特征并拍照记录。
1.4 天牛幼虫COI基因鉴定
天牛幼虫基因组DNA的提取:采用QIAGEN公司的DNeasy Blood&Tissue Kit试剂盒。具体操作步骤:①将在体积分数为96%乙醇中浸泡的天牛幼虫标本清洗干净晾干后,幼虫取其腹部组织≤25.0 mg置于研钵中充分研磨,置于1.5 mL的离心管中。②向离心管中加入180.0 μL裂解液(缓冲液ATL)和20.0 μL蛋白酶K,充分混匀后置于56℃恒温水浴锅6 h直到组织完全被消化。③在离心管中加入200.0 μL裂解液(缓冲液 AL)和200.0 μL的无水乙醇涡旋混合均匀,混合液转至吸附柱,柱子放在2.0 mL的收集管上。在室温下,8 000 r·min-1离心1 min,移除废液,吸附柱重新置于新的收集管内。④在吸附柱内加入500.0 μL洗液(缓冲液AW1),在室温下,8 000 r·min-1离心1 min,移除废液,吸附柱重新置于新的收集管内。⑤在吸附柱内加入500.0 μL洗液(缓冲液 AW2),在室温下,12 000 r·min-1离心3 min(缓冲液 AE),在室温下,8 000 r·min-1离心1 min。⑦重复第6步,以便提高产量。将提取的最终产物使用质量分数为1%的琼脂糖凝胶进行电泳,使用凝胶成像系统对电泳条带进行观察和拍照。抽提的DNA溶液置于-20℃冰箱保存备用。聚合酶链式反应(PCR)扩增:反应体系15.0 μL,其中包括1.0 μL DNA模板,上下游引物LCO1490(5′-GGTCAACAAATCATAAAGATATTGG-3′)和HCO2198(5′-TAAACTTCAGGGTGACCAAAAAATCA-3′)各0.3 μL,1.5 μL缓冲液(10×PCR缓冲液),1.5 μL氯化镁(MgCl2),1.5 μL核苷酸(dNTPs), 0.13 μL酶(rTaq),8.77 μL灭菌水。 PCR条件:95℃预变性3 min,95℃变性30 s,50℃退火40 s,72℃延伸50 s,35个循环,最后72℃延伸10 min。COI序列测序及分析:根据PCR检测结果,将样品送往生工生物测序部(上海)公司测序,测序成功后得到COI序列。在保证测序结果正确的基础上,对序列进行比对,除去序列两端冗余,保留共有序列进行同源比对。将比对结果导入MEGA 3.0系统发育分析程序。采用除权配对法(unweighted pair group method witharithmetic mean,UPGMAM),运用Maximum Composite Likelihood模型,并利用自举检验(bootstrap test)估计所构建系统树的可靠性,重复1 000次估计各个分支的置信值,并计算相关系数。
2 结果与分析
2.1 天牛种群分布情况
调查结果显示:浙江临安太阳镇和湍口镇山核桃林地都存在桑天牛Apripona germari,星天牛Anoplophora chinensis和云斑天牛Batocera horsfieldi等3种天牛同时危害,其中云斑天牛是主要种群,在这2个地区的天牛总量中均占1/2以上,其次是星天牛,桑天牛最少。在校园山核桃林地,主要是桑天牛和星天牛危害,没有发现云斑天牛危害,其中桑天牛占天牛总量比例(78.00%),其次是星天牛(表1)。

表1 不同种类天牛幼虫在山核桃林地天牛总量中的比例Table 1 Proportion distribution of different larvae in Carya cathayensis longhorned beetles
由表2可知:云斑天牛幼虫仅分布于山核桃树的树干部位,而在根部和枝条部位没有分布,且在树干不同高度均有分布,其中0~150 cm高度范围内的分布最多,占据93.85%;星天牛幼虫在根部和树干30 cm以下部位均有分布,且在根部分布最多,占据56.60%;桑天牛幼虫则都分布在枝条部位。以山核桃树的树干、根部和枝条部位作为天牛栖息资源,以山核桃的垂直高度作为资源序列进行划分,进行3种山核桃天牛的生态位宽度分析,从生态位宽度来看,云斑天牛幼虫的生态位宽度值最大(0.6),说明云斑天牛幼虫的取食范围较广,主要集中在树干部一定区间内;星天牛幼虫的生态位宽度值(0.4)仅次于云斑天牛幼虫的生态位宽度值,星天牛的蛀食范围除在根部以外,在树干近基部范围内也有危害;桑天牛幼虫的生态位宽度值较小,危害的位置集中在枝条部位。云斑天牛对星天牛和星天牛对云斑天牛的生态位重叠值分别为0.100 0和0.066 7,桑天牛对星天牛和云斑天牛的生态位重叠值都为0。
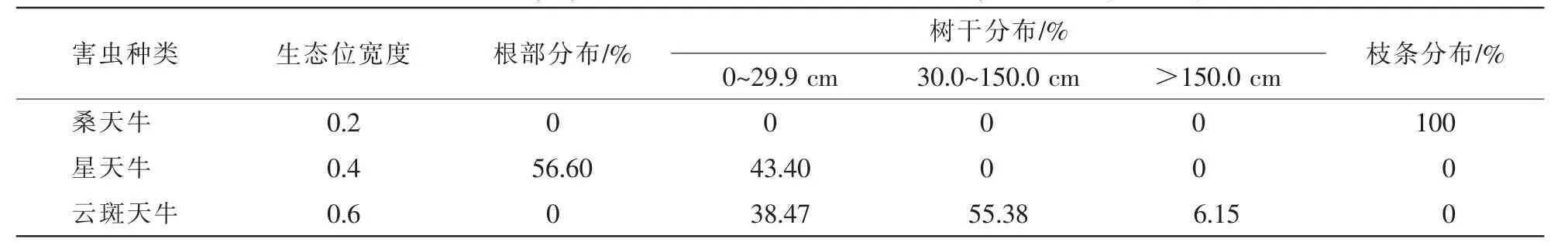
表2 天牛幼虫在山核桃树上不同部位的分布比例Table 2 Distribution proportion of larva of beetle on different parts of Carya cathayensis
2.2 天牛危害特征
调查结果显示:不同天牛幼虫在山核桃树上表现出不同的危害症状,如排粪状、蛀道和产卵刻槽特征(表3,图1,图2和图3)。

表3 不同天牛幼虫在山核桃上的危害特征Table 3 Damage features of different longhorned beetle larvae on Carya cathayensis

图1 山核桃不同天牛幼虫排粪状Figure 1 Damage shape of defecation of different longhorned beetle larvae on Carya cathayensis

图2 山核桃不同天牛幼虫蛀道Figure 2 Larvae burrow of different longhorned beetle larvae on Carya cathayensis

图3 山核桃不同天牛产卵刻槽Figure 3 Oviposition groove of different longhorned beetles on Carya cathayensis
桑天牛幼虫主要危害山核桃树的枝条部位。在幼虫排粪期,可明显观察到每隔一段距离就有一排粪孔。剥开蛀道可发现,其幼虫蛀道一般较通直,蛀道内往往伴随一股发酵的酒糟味。雌成虫在枝条上产卵时的刻槽常咬成回形针状。
星天牛幼虫主要危害山核桃树30 cm以下树干和根部部位。在幼虫排粪期,幼虫常常在树干基部地面土壤排出土堆状的虫粪。幼虫蛀道从近地面树干一直延伸到地下树根部位。星天牛雌成虫在树干基部的产卵刻槽为 “T”字型。
云斑天牛幼虫主要危害山核桃树的树干部位。幼虫危害后,树皮向外鼓起纵裂,虫粪为粗木丝状,常堆满整个地面。幼虫蛀道内部呈黑色,云斑天牛雌成虫在树干上的产卵刻槽一般为唇形。
2.3 不同山核桃天牛成虫雌雄鉴别特征
通过观察,发现不同种类天牛成虫雌雄之间都具有显著的形态特征,可以用来鉴别天牛雌雄个体(表4)。

表4 不同天牛成虫雌雄鉴别特征Table 4 Identification characteristic between male and female of different longhorned beetle adults
桑天牛雄成虫触角超出身体长的1/3;而雌成虫触角稍稍超出身体部位。因此,触角是否超过体长的1/3是判断桑天牛雌雄成虫的重要依据。
星天牛雄成虫触角长度接近超出身长的1/2;而雌成虫触角稍稍超出身体部位。雄成虫腹部末端较尖,宽度比雌成虫要窄;雌成虫腹部末端较钝平,宽度比雄成虫宽。另外,雄成虫的触角正面是黑色和灰白色相间结构,背面则是整条近乎的黑色;而雌成虫触角正反两面都是一样的黑色和灰白色相间结构。因此,通过观察触角背面是否为黑色,就可以简单的区分星天牛雌雄个体。
云斑天牛雄成虫触角的末端3节超出腹部末端;而雌成虫触角稍稍超出身体部位。另外,雄成虫腹部最后一节没有任何竖缝线;而雌成虫腹部最后一节具有一明显的竖缝线。因此,观察腹末是否有竖缝线是区别云斑天牛雌雄成虫的重要特征。
2.4 天牛幼虫前胸背板特征
在3个试验样地中收集到的天牛幼虫通过用电子显微镜观察并拍照记录其前胸背板特征。通过比较发现,桑天牛、星天牛和云斑天牛幼虫前胸背板具有显著的形态特征:桑天牛幼虫的前胸背板具有放射线状瘤点,星天牛幼虫前胸背板前方有2个褐色飞鸟纹,云斑天牛幼虫前胸背板前方有2个小黄点。通过这些简易的形态特征可以很方便地识别这3种天牛幼虫(表5和图4)。

表5 不同天牛幼虫的前胸背板特征Table 5 Pronotum characteristic of different longhorned beetle larvae
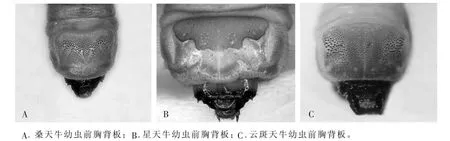
图4 不同天牛幼虫前胸背板的形态特征图Figure 4 Figure of pronotum morphological characteristic of different longhorned beetle larvae
2.5 基于COI序列的系统发育树分析
由图5 COI序列聚类结果显示:共比对了9种同属于沟胫天牛亚科常见天牛的COI基因序列,9种天牛分别属于粒肩天牛属Apriona,星天牛属Anoplophora,白条天牛属Batocera和墨天牛属Monochamus。经COI基因鉴定,其结果如下:上述A天牛幼虫基因比对为桑天牛A.germari,上述B天牛幼虫基因比对为星天牛A.chinensis,上述C天牛幼虫基因比对为云斑天牛B.horsfieldi。从亲缘关系来看,桑天牛和云斑天牛的亲缘关系更近,而这2种天牛与星天牛的亲缘关系相对较远。COI基因鉴定结果与上述通过幼虫前胸背板特征区分不同天牛幼虫的结果相一致,说明通过幼虫前胸背板特征快速区分桑天牛、星天牛和云斑天牛幼虫种类的方法是准确的。

图5 天牛的UPGMA系统发育树Figure 5 Phylogenetic tree revealed by UPGMA cluster analysis of genetic distance based on the sequence
3 结论与讨论
湍口镇和太阳镇的山核桃林地都发现了桑天牛、星天牛和云斑天牛,且以云斑天牛为主要种群,而校园山核桃林地只发现了桑天牛和星天牛,没有发现云斑天牛,且桑天牛占绝对种群。这可能跟林地山核桃的树龄有关,湍口镇和太阳镇的山核桃树龄一般为20~30年生,正值青壮年,处于产山核桃的高峰期,而校园林地的山核桃树龄一般为7~8年生,尚处于幼龄阶段。金凤等[11]研究发现:桑天牛的刻槽主要集中为2~3年生的枝条上,相关系数显著性检验表明:刻槽与木质部厚度、枝条直径极显著相关。刁志娥等[12]报道,在山东东营市,云斑天牛主要危害胸径在8 cm以上生长的白蜡树Fraxinus chinensis,这都说明不同种类天牛在山核桃树上选择其危害部位与其树木胸径有一定的要求,也可能与树皮厚度有一定关系。因此,我们推测,在成年山核桃林地,以云斑天牛和星天牛危害为主,而在低龄山核桃林地,以桑天牛危害为主。
桑天牛、星天牛和云斑天牛幼虫在山核桃树上占据着不同的分布位置,分别为枝条、根部和树干。3种山核桃天牛在山核桃上的生态位宽度揭示了3种物种之间的资源利用关系,云斑天牛的生态位宽度为0.6,星天牛的生态位宽度为0.4,云斑天牛对星天牛和星天牛对云斑天牛的生态位重叠值分别为0.100 0和0.066 7,两者之间资源利用上存在一定的生态位重叠,主要在树干部0~30 cm区间有重叠分布,而桑天牛对星天牛和云斑天牛的生态位重叠值都为0。PIANKA等[13]认为:竞争与生态位重叠之间是密切相关的,而COLWELL[14]与ABRAMS等[15]则认为竞争与生态位重叠无关。李契等[16]认为生态位重叠并不一定导致竞争,而资源量和供求比以及资源满足生物需要的程度对研究生态位重叠和竞争的关系非常重要,若资源很丰富,那么2个物种间就完全可共同利用同一资源而不给对方带来竞争。目前,暂时还没发现同一株山核桃树上同时存在星天牛和云斑天牛幼虫在危害,是否因为这2种天牛存在一定的生态位重叠,从而存在一定的竞争关系,互相排斥,还有待进一步的深入研究。桑天牛幼虫的分布与云斑天牛幼虫和星天牛幼虫的分布是相对分离的,对空间资源的共享程度小,说明它们对资源的需求具有较大的差异,这有利于桑天牛和其他2种天牛在竞争条件下的共存。
桑天牛、星天牛和云斑天牛幼虫在幼虫排粪状和蛀道等危害症状方面具有一些明显的区别特征。同时,这3种天牛成虫产卵刻槽也有着不同的特征,桑天牛在山核桃树上的产卵刻槽为回形针形,这与金凤等[11]报道桑天牛产卵刻槽为 “U”字形不太一致,星天牛产卵刻槽为 “T”字形,云斑天牛产卵刻槽为唇形或椭圆形,星天牛和云斑天牛的产卵刻槽特征和徐起等[17]和严敖金等[18]报道的星天牛和云斑天牛产卵刻槽特征结果相一致。这些危害特征都有助于我们初步判断是哪种天牛幼虫在进行危害。
星天牛雌成虫触角阴阳两面接近一样,为一半黑色与一半灰白色相间,雄成虫触角阳面为一半黑色与一半灰白色相间,触角阴面则近乎整条黑色。王涛等[19]报道了相似的方法可快速准确鉴别光肩星天牛雌雄成虫。我们通过研究发现了一种新的云斑天牛雌雄鉴别的方法,在整个成虫期都可以快速准确鉴别云斑天牛雌雄成虫,即云斑天牛雌成虫在其腹部最后一节具有一明显的竖缝线,雄成虫则在腹部最后一节没有任何竖缝线。严敖金等[18]通过交配斑来鉴别云斑天牛雌雄成虫,即已交配的雌成虫其鞘翅中上部绒毛磨损形成明显的交配斑,已交配的雄虫其中胸腹板及前、中足基节绒毛磨损形成相应的交配斑,通过交配斑只能鉴别出处于交配期的云斑天牛雌雄成虫,但在交配期前就不能根据此方法来鉴别。
通过天牛幼虫前胸背板特征可以快速准确鉴别出危害山核桃树的桑天牛、星天牛和云斑天牛幼虫,借助分子鉴定方法也进一步确认了此鉴别方法的准确性。
总之,不同种类天牛幼虫的主要危害症状和典型形态特征,都可以给我们提供重要的信息,用来鉴别山核桃天牛的种类。山核桃天牛分布规律、危害症状以及鉴别特征,对于指导山核桃天牛害虫防控工作具有重要的意义。
[1] 吴伟文,麻耀强,吴伟志.论杭州市山核桃产业的发展与对策[J].浙江林业科技,2003,23(3):57-60.
WU Weiwen,MA Yaoqiang,WU Weizhi.About development and countermeasures for hickory nut[J].J Zhejiang For Sci Technol,2003,23(3):57-60.
[2] 宋朝枢.自然保护区工作手册[M].北京:中国林业出版社,1988:41-43.
[3] 黎章矩,钱莲芳.山核桃科研成就和增产措施[J].浙江林业科技,1992,12(6):49-53.
LI Zhangju,QIAN lianfang.Achievements and measures of increase production of Carya cathayensis Sarg.[J].J Zhejiang For Sci Technol,1992,12(6):49-53.
[4] 吕秋菊,沈月琴,高宇列,等.山核桃产业的发展过程、动因及展望[J].浙江农林大学学报,2012,29(1):97-103.
LÜ Qiuju,SHEN Yueqin,GAO Yulie,et al.Development process,agents and prospect of hickory industry[J].J Zhejiang A&F Univ,2012,29(1):97-103.
[5] 童根平,王卫国,张圆圆,等.大田条件下山核桃林地土壤和叶片养分变化规律[J].浙江林学院学报,2009,26(4):516-521.
TONG Genping,WANG Weiguo,ZHANG Yuanyuan,et al.Seasonal changes of soil and leaf nutrient levels in a Carya cathayensis orchard[J].J Zhejiang For Coll,2009,26(4):516-521.
[6] 慕康国,赵秀琴,李健强,等.矿质营养与植物病害关系研究进展[J].中国农业大学学报,2000,5(1):84-90.
MU Kangguo,ZHAO Xiuqin,LI Jianqiang,et al.Progressing on the relation between mineral nutrients and plant disease[J].J China Agric Univ,2000,5(1):84-90.
[7] LEVINS R.Evolution in Changing Environments:Some Theoretical Explorations[M].Princeton:Princeton University Press,1968.
[8] 徐汝梅.昆虫种群生态学[M].北京:北京师范大学出版社,1987.
[9] 陈辉,唐明,叶宏谋,等.秦岭华山松小蠹生态位研究[J].林业科学,1999,35(4):40-44.
CHEN Hui,TANG Ming,YE Hongmou,et al.Niche of bark beetles with in Pinus armandi ecosystem in Inner Qinling mountains[J].Sci Silv Sin,1999,35(4):40-44.
[10] 吕仲贤,杨樟法,王桂跃,等.玉米螟和桃蛀螟在玉米上的生态位及其种间竞争[J].浙江农业学报,1995,7(1):31-34.
LÜ Zhongxian,YANG Zhangfa,WANG Guiyue,et al.The niche of corn borer and peach borer and their interspecific competition in corn[J].Acta Agric Zhejinag,1995,7(1):31-34.
[11] 金凤,嵇保中,刘曙雯,等.桑天牛雌成虫产卵器解剖和刻槽及产卵习性研究[J].林业科学研究,2007,20(3):394-398.
JIN Feng,JI Baozhong,LIU Shuwen,et al.Studies on the anatomical structure of ovipositor and the gnawing nidus and oviposition habits of female adult Apriona germari[J].For Res,2007,20(3):394-398.
[12] 刁志娥,丁福波.云斑天牛在白蜡树上的发生与防治研究[J].华东昆虫学报,2004,13(2):49-52.
DIAO Zhie,DING Fubo.Occurrence and control of white-stiped longhorn,Batocera horsfieldi(Hope)(Coleoptera:Cerambycidae)on Chinese ash,Fraxinus chinensis Roxb.[J].Entom J East China,2004,13(2):49-52.
[13] PIANKA E R.Competition and niche theory[G]//USHER M B,MAY R M.Theoretical Ecology:Principles and Applications.Oxford:Black-Well Scientific Publications,1976:114-141.
[14] COLWELL R K,FUTUYMA D J.On the measurement of niche breadth and overlap[J].Ecology,1971,52(4):567 -576.
[15] ABRAMS P.Some comments on measuring niche overlap[J].Ecology,1980,61(1):44-49.
[16] 李契,朱金兆,朱清科.生态位理论及其测度研究进展[J].北京林业大学学报,2003,25(1):100-107.
LI Qi,ZHU Jinzhao,ZHU Qingke.A review on niche theory and niche metrics[J].J Beijing For Univ,2003,25(1):100-107.
[17] 徐起.木麻黄害虫星天牛生活习性及防治[J].林业科学研究,1997,10(5):551-555.
XU Qi.Habits of Anoplophora chinensis which causes harm to Casuarina equisetifolia and its control[J].For Res, 1997,10(5):551-555.
[18] 严敖金,嵇保中,钱范俊,等.云斑天牛Batocera horsfieldi(Hope)的研究[J].南京林业大学学报(自然科学版),1997,21(1):1-6.
YAN Aojin,JI Baozhong,QIAN Fanjun,et al.A study on Batocera horsfieldi(Hope)[J].J Nanjing For Univ Nat Sci Ed,1997,21(1):1-6.
[19] 王涛,温俊宝,许志春.鉴别光肩星天牛雌雄成虫的一种简便方法[J].中国森林病虫,2004,23(5):42-44.
WANG Tao, WEN Junbao, XU Zhichun.A simple and convenient method used in identifying the sex of Anoplophora glabripennis(Mostsch.)adults[J].For Pest Dis,2004,23(5):42-44.
Damage investigation and an identification method for different species of Carya cathayensis longhorned beetles
ZHANG Dongyong1,ZHOU Shuyong2,ZHU Ning1,FAN Jianting1
(1.School of Forestry and Biotechnology,Zhejiang A&F University,Lin’an 311300,Zhejiang,China;2.Forest Pest Control and Quarantine Station of Linhai City,Linhai 317000,Zhejiang,China)
A variety of longhorned beetles simultaneously occur on Carya cathayensis host with increasingly serious damage,which become an important factor limiting the pecan production.To explain the damage profile from longhorned beetle species and to help prevent and control damage on a Carya cathayensis forest,we carried on a systematic research on a C.cathayensis forest in Lin’an to study damage from the species,recognition of typical damage shapes,and identification methods of different species by host anatomy,morphological identification and molecular identification method.Results showed that there were mainly three longhorned beetle species,Apriona germari,Anoplophora chinensis,and Batocera horsfieldi in the C.cathayensis forest of Lin’an. A.germari larvae were 100%distributed on branches,A.chinensis larvae were 100%on roots and trunks 30 cm below ground,and B.horsfieldi larvae were 93.9%distributed on the trunk 0-150 cm above ground.Different species of longhorned beetle larvae produced different typical symptoms for the shape of the damage due to defecation,larvae burrows,and oviposition grooves.There were strong morphological differences between male and female adults of A.germari,A.chinensis,and B.horsfieldi.In addition,the larvae pronotum of these three species of longhorned beetles had different typical characteristics making it possible to quickly and accuratelydistinguish the type of longhorned beetle species.Corresponding molecular identification results also proved the accuracy of this method.We speculate that B.horsfieldi and A.chinensis mainly damage adult C.cathayensis trees,and A.germari mainly damage young ones with different species of longhorned beetle larvae occupying different niches.In addition,different typical symptoms and morphological characteristics could be important in preventing and controlling C.cathayensis longhorned beetles.[Ch,5 fig.5 tab.19 ref.]
forest protection;Carya cathayensis;longhorned beetles;investigation;larvae;identification
S763.3
A
2095-0756(2017)01-0042-08
2016-01-15;
2016-03-02
国家林业公益性行业科研专项(201304403)
张冬勇,从事森林害虫控制研究。E-mail:979224878@qq.com。通信作者:樊建庭,副教授,博士,从事入侵生物学、化学生态学和森林害虫控制研究。E-mail:fanjt123@sina.com
浙 江 农 林 大 学 学 报,2017,34(1):50-55
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2017.01.008
