天然产物合成生物学体系的优化策略
2017-02-21匡雪君邹丽秋孙超陈士林中国医学科学院北京协和医学院药用植物研究所北京0093中国中医科学院中药研究所北京00700
匡雪君邹丽秋孙超陈士林(. 中国医学科学院北京协和医学院药用植物研究所,北京 0093;. 中国中医科学院中药研究所,北京 00700)
天然产物合成生物学体系的优化策略
匡雪君1邹丽秋1孙超1陈士林2
(1. 中国医学科学院北京协和医学院药用植物研究所,北京 100193;2. 中国中医科学院中药研究所,北京 100700)
天然产物是新药研发的重要源泉。天然产物合成生物学通过设计、重构目标化合物的高效生物合成途径,借助宿主改造,利用发酵生产目标化合物,可以有效弥补有机合成化学在复杂天然产物类药物生产方面的不足。虽然合成生物学已经取得了一些进展,但是通过合成生物学技术使目标产物的产量达到工业化生产水平依然是一项非常具有挑战性的任务。综述了天然产物合成生物学体系的优化策略,通过综合运用单个元件、外源代谢途径、底盘系统和发酵条件的优化技术,可以实现生物合成系统的最优化,最大化目标产物的产量,为来源稀缺的复杂天然产物的开发提供持续、稳定、经济的原料供给,推动天然产物类新药的研发。
天然产物;合成生物学;优化;底盘
天然产物在药物的发现和研制中发挥着重要作用,以天然产物为基础研制和开发新药一直是化学界和医药界重点关注的领域[1,2]。 天然产物虽然在整个已知化合物中所占比例很小,但以之为基础发展成为新药的比例却很大。1981-2014年间批准上市的1 562个药物中约有一半与天然产物有关;1940-2014年间,国际上共有175个小分子药物被批准用于癌症治疗,其中有85个(即约49%)来源于天然产物或其衍生物[3]。这些有价值的天然产物通常在原物种中含量很低,化学提取不仅成本高,而且会对环境资源造成破坏;有些天然产物由于含有多个手性中心或结构不稳定,用化学方法全合成难度很大,因此,利用合成生物学技术合成具有药用价值的天然产物受到越来越多的关注。目前,已有学者在工程菌中成功异源合成了萜类化合物青蒿素和人参皂苷、生物碱类化合物诺斯卡品、黄酮类化合物儿茶素和柚皮素等重要天然产物[2,4]。
虽然天然产物合成生物学研究进展迅速,但是使异源合成的目标产物的产量达到工业化生产水平依然是一项富有挑战性的任务。由于底盘细胞是非常复杂的系统,外源途径的导入会使底盘系统产生一系列反应,包括生长速率的调节、热激反应、压力反应、严谨反应等[5],进而引起质粒的不稳定、细胞的裂解和细胞遗传信息的改变,导致产物无法合成或产量极低。本文提出了天然产物合成生物学体系的优化策略(图1),综合运用单个元件、外源代谢途径、底盘系统和发酵条件的优化技术,对宿主细胞进行改造,人工精确调控外源基因,实现多基因的联合协同表达,使生物合成系统最优化,最大化目标产物的产量,为天然药物的研发提供新的先导化合物。

图1 天然产物合成生物学体系的优化策略
1 单个元件的优化
合成生物学中的元件包括核糖体结合位点(Ribosome-binding site,RBS)序列、复制起始位点、启动子、终止子、功能蛋白等[6]。单个元件的微调能有效改善基因线路的性能,增加途径中酶基因的表达量和生化反应的效率。
1.1 RBS优化
核糖体结合位点是调节翻译强度的重要元件,人工设计的RBS文库可以方便地通过引物设计和PCR扩增来实现与特定基因的连接,从而在翻译水平上对基因的表达强度进行调节[7]。可以通过RBS计算器(RBS Calculator)[8]或者RBS 设计器(RBS Designer)[9]对RBS文库的范围和强度进行预测。需要注意的是,此方法一般用于大肠杆菌系统,在酵母中不适用。目前,已有研究者成功在大肠杆菌中通过修改途径中关键酶的RBS优化了抗癌类化合物紫苏醇、抗肿瘤类物质司帕霉素和脱氧紫色杆菌素在工程菌中的产量[10]。
1.2 启动子修饰
启动子是结构基因上游起始转录的一段序列。启动子结构会影响其本身与RNA聚合酶的亲和力,从而影响该启动子对基因的转录效率。启动子的强度决定了结构基因的转录强度[11]。对启动子进行修饰(如对启动子进行突变、改变启动子序列的长度或使用强启动子)可以调控代谢途径中酶基因的表达效率[12]。Redding-Johanson 等[13]的实验表明,使用强启动子能增加甲羟戊酸激酶和磷酸甲羟戊酸激酶的表达活性,进而提高工程菌中紫穗槐-4,11-二烯的产量。Anthony等[14]通过优化启动子的长度来平衡甲羟戊酸激酶(MK)和紫穗槐-4,11-二烯合酶(ADS)基因的表达,使抗癌药物的前体紫穗槐-4,11-二烯(AD)的产量提高了5倍。
1.3 质粒拷贝数优化
增加基因的拷贝数是调节异源基因在底盘系统中表达最常用的工具之一。将基因克隆到多拷贝的表达质粒上增加基因的拷贝数能使基因获得高水平表达,但会给细胞带来更大的生理负担,所以质粒拷贝数不是越高越好[15,16]。Jawed等[17]在大肠杆菌中建立了丁酸的生物合成途径,他们发现质粒的拷贝数能对丁酸的产量产生影响,使用中等拷贝数的质粒能取得最好的效果,使丁酸的产量达到1.5 g/L。
1.4 密码子优化
采用宿主偏爱的密码子,减少或避免使用稀有密码子是提高途径酶基因异源表达水平的重要手段[18]。3-类固醇脱氢酶(3-ketosteroid-Δ1-dehydrogenase,KSDD)能将雄甾-4-烯-3,17-二酮催化为雄甾-1,4-二烯-3,17-二酮,由于KSDD是途径中的限速酶,途径重构时较低的酶活性使这步反应受到限制。Shao等[19]采用密码子优化的方法将KSDD在枯草芽孢杆菌( Bacillus subtilis)中的酶活性提高了7倍,达到12.3 U/mg,结合发酵条件的优化,使雄甾-1,4-二烯-3,17-二酮的产量达到8.76 g/L。
2 代谢途径的系统优化
对生物合成途径单一元件进行修饰尽管一定程度上能提高目的产物的产量,但局部的改善往往导致中间产物的积累,增加细胞负担,甚至可能会对细胞产生生长抑制毒性,不利于目标产物的获得。对于代谢途径的平衡优化,主要采用一种基于全局的数学算法和生物信息分析,实现整条途径代谢流的平衡[20]。
2.1 多元模块代谢工程
多元模块代谢工程(Multiple module metabolic engineering,MMME)是将整个代谢途径分为不同的模块,再通过系统地改造各个模块的复制起始位点、启动子或RBS序列来协调不同模块的表达强度,只需少数模块的组合,就可以在大范围内优化代谢通途径[21]。Wu 等[22]将松属素合成途径分为4个模块,通过改变每个模块的质粒拷贝数,限制了毒性物质肉桂酰辅酶A的积累,使松属素的产量达到40.02 mg/L。Liu等[23]在枯草芽孢杆菌中构建了n-乙酰氨基葡糖(GlcNAc)合成途径,并把整个途径分为3个模块:异源的n-乙酰氨基葡糖合成模块、n-乙酰氨基葡糖醣酵解模块和肽聚糖合成模块。使用调控小RNA来抑制n-乙酰氨基葡糖醣酵解和肽聚糖的合成,通过同时平衡3个模块中内源和外源基因的表达,使n-乙酰氨基葡糖的产量达到36.35 g/L。
2.2 多元自动化基因组工程
多元自动化基因组工程(Multiplex automated genome engineering,MAGE)是一种高通量修饰基因组的工具,能用目标序列同时替换基因组中多个位点。人工设计的寡核苷酸双末端均带有与基因组插入位点同源的序列,中间区域含有需要替换的DNA片段,在宿主DNA复制过程中,在λ噬菌体Red重组酶介导下进行高效同源重组[26]。由于MAGE通过启动子替换或者核糖体结合位点序列替换这两种方式调节基因的表达,所以寡核苷酸链中间区域含有启动子序列或核糖体结合位点序列。MAGE能同时调节途径中多个基因(>20)的表达[27]。Sharan等[26]利用不同T7启动子来替换吲哚合成途径中的12个基因的启动子,最优菌株中靛蓝的产量比原始菌株提高4倍。Wang等[28]利用不同的RBS序列替换番茄红素合成途径中20个基因中的原始RBS序列,找到了最优的组合,使番茄红素的产量提高了5倍。
2.3 多元循环质粒工程
与多元自动化基因组工程类似,多元循环质粒工程(Multiplex Iterative Plasmid Engineering,MIPE)也是利用寡核苷酸介导的λ噬菌体Red重组酶的同源重组,将携带多个基因的质粒和针对不同基因设计的多个寡核苷酸共同转化进入宿主细胞,用寡核苷酸目的片段替换质粒上的相应区域,使质粒产生点突变。该法与限制酶切介导的共选择(Restriction digestion mediated co-selection,RD CoS)策略相结合,每个循环用一个限制酶切位点作为共选择标记就能用于质粒多个循环的筛选[29]。Li等[29]运用MIPE对核黄素的生物合成途径中5个基因进行了组合优化,首先从大肠杆菌基因组中克隆出这5个基因,并连接到质粒的同一个操纵子中。然后将含有不同的核糖体结合位点的寡核苷酸混合物、携带基因的质粒与用于筛选的寡核苷酸共转化大肠杆菌,质粒上核糖体结合位点区域经同源重组发生突变,经过3轮循环后,产生了1×107种组合,最后将核黄素的产量提高了2.67倍。
2.4 组合转录工程优化法
在真核微生物中(如酿酒酵母),运用启动子工程能对途径中基因进行转录水平的优化,通常使用随机突变的方法来得到一系列不同强度的启动子[30]。采用组合转录工程优化法(Customized optimization of metabolic pathways by Combinatorial transcriptional engineering,COMPACTER)将组装技术成功应用于代谢途径的组合优化,能同时调节多个酶的表达水平。该技术通过将不同强度的启动子与不同代谢途径的基因进行组合,再用高通量的筛选方法来构建和筛选出最高效的代谢途径,由此实现对代谢途径中多个基因表达强度的组合优化[31]。COMPACTER技术已经应用到酵母木糖利用途径的3个酶和纤维二糖利用途径中2个酶的优化,使工程菌株中纤维二糖的消耗加速了2.1倍,并且使乙醇的产量增加2.3倍[32]。
2.5 可调控基因间序列优化
通过可调控基因间序列(Tunable intregenic regions,TIGR)能在一个操纵子内对多个基因表达强度同时进行调节,且在原核生物和真核生物中都适用[33]。TIGRs由多种控制元件组成,它们是:mRNA二级结构、核糖核酸酶断裂位点和RBS隔离序列等[34]。Pfleger等[35]在大肠杆菌中对甲羟戊酸途径中3个基因(atoB、HMGS、tHMGR)构建TIGR文库,优化多种控制元件,使甲羟戊酸的产量提高了7倍。采用TIGR法人工设计操纵子,能影响mRNA的稳定性或翻译起始效率,并消除不同基因的干扰,不需要多个启动子就能在转录后水平同时对多个基因的表达强度进行调节。
3 底盘系统优化
一些常见的模式微生物,如大肠杆菌、酿酒酵母、枯草芽胞杆菌等是近年来被广泛使用的异源合成底盘细胞[1]。它们均具备作为一个优良底盘细胞所需的特征,如生长快速、易于基因工程操作、适合大规模培养、工业化控制简便等。底盘细胞的初生或次生代谢产物能为异源生物合成途径的构建提供前体,优化异源宿主能增加主流代谢途径中的底物供应,促进途径通量的重新分配,增加天然产物的产量。
3.1 目标代谢途径的上调
除了增加外源基因的表达量外,使宿主内源基因的表达上调也能促进产物的生物合成。Fatma等[36]通过计算机软件分析,在大肠杆菌中选出可能参与长链脂肪醛转化为脂肪醇的35个内源性酶。研究发现敲除YbbO基因会使脂肪醇的含量下降90%,证实该基因在脂肪醇合成中具有重要作用。通过优化YbbO基因的表达,使长链脂肪醇的产量达到169 mg/L,此外,通过调节脂肪酸和磷脂的生物合成使脂肪醇的产量进一步增加60%。通过调节途径中的转录因子也能使目标代谢途径上调,Santos等[37]使用全转录工程法(Global transcription machinery engineering,gTME),对rpoA和rpoD转录因子进行突变,使酪氨酸的产量达到13.8 g/L。
3.2 竞争性代谢途径的抑制
下调或阻断竞争性代谢支路能够限制或减少流入竞争代谢途径中的底物供应,从而维持重要前体或中间产物在底盘细胞中的含量。可以通过基因敲除或使用弱启动子来抑制或下调竞争代谢途径以获得更多的前体,促进途径通量的重新分配[40]。Paradisee等[41]通过敲除竞争性甾体合成途径中的ERG9基因,减少进入甾体生物合成途径中的 FPP代谢流,从而使进入青蒿酸生物合成途径的 FPP流量增加,进而增加了目的产物AD和青蒿酸的产量。Westfall等[42]采用多种策略来提高丙二酰辅酶A在大肠杆菌中的含量,例如过表达乙酰辅酶A合酶基因和敲除竞争途径,通过消除丙二酰辅酶A降解途径,最终使丙二酰辅酶A在胞内的产量提高了15倍。有时同一个前体可能同时作为不同次生代谢途径中的底物用于不同产物的合成,如缬氨霉素和菌丝霉素合成途径竞争一个共同的前体物质2-酮异戊酸,通过使菌丝霉素合成的基因簇和相关基因失活,缬氨霉素的产量比野生型菌株提高了4倍[45]。
3.3 底盘基因组的简化
理想的底盘细胞应该具有最小化基因组,即维持细胞的生长繁殖所必须的最少基因(必需基因)。基因组的适当精简将为重要天然产物的异源生物合成提供理想的底盘细胞。基因组的简化可使细胞代谢途径得以优化,不仅能改善细胞对底物、能量的利用效率,更好地耐受引入的各种酶和代谢产物的代谢负担;还能更好地保持外源基因网络,大大提高细胞生理性能的预测性和可控性[46]。采用CRISPR-Cas9技术、多元自动化基因组工程、基于位点特异性重组酶的同源重组等基因组编辑技术,能在基因组范围内对任意的多个位点进行同步修饰,实现底盘细胞的改造[47]。2010年,Komatsu等[48]利用阿维链霉菌(S. avermitilis)宿主分别异源重构了链霉素和头霉素C基因簇,阿维链霉菌的基因组大小为9.02 Mb,采用双交换重组与位点特异性重组技术将阿维链霉菌染色体左端亚端粒区域与次生代谢产物合成相关的基因与转座子酶基因删除(共约1.4 Mb),得到了一系列突变菌株。链霉素是一种氨基糖苷类抗生素,可由灰色链霉菌(S. griseus)产生,但是阿维链霉菌不能合成,故将灰色链霉菌中链霉素生物合成的基因簇(约41.2 kb)导入阿维链霉菌野生型和突变菌株,结果发现基因组简化后的突变菌株中链霉素含量明显高于野生型阿维链霉菌菌株;头霉素C是一类β内酰胺类抗生素,将棒状链霉菌(S. clavuligerus)中的头霉素C合成相关的基因簇(约35 kb)导入阿维链霉菌菌株,结果也和前述一致,进一步证明了宿主基因组简化的重要性。由于基因组简化后的链霉菌菌株删除掉了多种主要的内源性次生代谢产物相关的基因,积累了更多的前体可用于链霉素或头霉素C的合成,故产量得到提高。2013年,该课题组又将20多个基因簇引入阿维链霉菌,大部分实现了产物的成功表达,证明了阿维链霉菌拥有作为次生代谢产物通用宿主的潜能[49]。
4 发酵条件优化
除了对代谢途径自身进行优化外,发酵条件对异源宿主中目标产物及其前体的产量也有很大影响。即使天然产物工程菌的性能优良,如果缺乏合理的发酵工艺也不能将其潜力发挥到极致[50]。通过前体饲喂、培养基优化、温度控制和诱导物添加等方式均能有效增加菌株中目标产物的产量。
4.1 前体饲喂
前体饲喂是提高目标产物产量的最有效的方法之一[51,52]。前体或中间产物的有效供应是天然产物的合成和生产的先决条件,前体的增加对于次生代谢产物的积累有促进作用,通过中间产物的添加,降低中间产物合成的能量消耗,从而促进代谢流向目标产物[53]。Dhakal等[54]在诺卡氏菌中重构了阿根诺卡菌素途径,通过饲喂前体物质脯氨酸与葡萄糖并调节饲喂时间,阿根诺卡菌素的产量达到84.9 mg/L,与没有饲喂前体的菌株相比,增加了24倍。
4.2 培养基优化
目前发酵产品的大规模工业化生产多采用液体培养基进行深层发酵。使用的培养基必须满足微生物生长、繁殖的需要,有利于微生物大量合成有机物[55]。培养基成分对工程菌发酵具有重大影响,不同的生物合成途径采用不同的碳源进行发酵,常用的碳源有:葡萄糖、木糖、甘油和酵母提取物等[56]。Westfall等[42]研究发现,将乙醇和葡萄糖作为培养基的混合碳源时,发酵产生的紫穗槐-4,11-二烯产量显著增加,而将碳源变成半乳糖时,发酵产物中的青蒿酸产量上升明显。
4.3 温度控制
每种微生物都有最适生长温度,对于某种特定的微生物而言,只能在一定的温度范围内生长,微生物的不同生理活动也需要在不同的温度条件下进行,所以发酵速度、生长速率和代谢产物积累的最适温度往往不同[57]。例如,乳酸链球菌在40℃时发酵速度最快,在34℃时繁殖速度最快,25-30℃时细胞产量最高[58]。所以,研究不同微生物在生长或积累代谢产物阶段时的最适温度,对于用变温发酵来提高发酵生产效率具有重要意义。
4.4 诱导物添加
诱导物的种类、浓度和诱导起始时机、诱导时间均能影响产物的产量[59]。因此,兼顾工程菌生理状态和增殖能力选择适当的诱导时机、诱导物用量和诱导时间,能够有效增加工程菌中外源途径蛋白的表达,提高目标产物的产量。Sassi等[60]利用解脂耶氏酵母作为细胞工厂生产重组蛋白,油酸能作用于诱导型启动子pLIP2控制基因的表达,把油酸与葡萄糖的混合物(3∶2)添加到培养基中,重组蛋白的产量增加了10倍。
5 总结与展望
表1总结了近年来在天然产物合成生物学体系优化研究领域的主要进展。在体系优化过程中,一个途径中取得最大产量的方法可能在另一个途径中并不适用,找到正确的优化方法是增加产量的关键。可以选择一种适合高效的优化技术或是将不同技术相结合,以此来取得产量最大化,为大规模工业化生产奠定基础。例如,Alper等[61]将基因敲除技术与gTME方法相结合来优化番茄红素的产量。
质粒拷贝数优化、启动子替换、核糖体结合位点优化、密码子优化是常用于提高基因表达水平的方法,已在多个天然产物的途径优化中得到应用(表1),尽管单一元件的修饰一定程度上能提高目标产物的产量,但天然产物代谢途径通常相对复杂,包含多个关键酶基因与调控基因,对单个元件的修饰可能只会增加部分酶的表达量,对整个代谢途径的影响不明显,甚至中间产物的过度积累有时可能会使细胞产生毒性反应,影响细胞正常的生长和繁殖;而采用组合优化的方法对途径中多个元件进行同时调节能有效避免这种情况的发生。如果目标产物的合成途径涉及的基因数目不多,可以尝试优化可调控基因间序列,将途径中几个基因置于一个操纵子内,进行协同控制;当途径复杂,涉及多个代谢支路,可以使用多元模块代谢工程、多元自动化基因组工程、多元循环质粒工程等方法同时修饰多个元件,使整体的代谢达到平衡,以获得异源合成体系的最优化[62]。
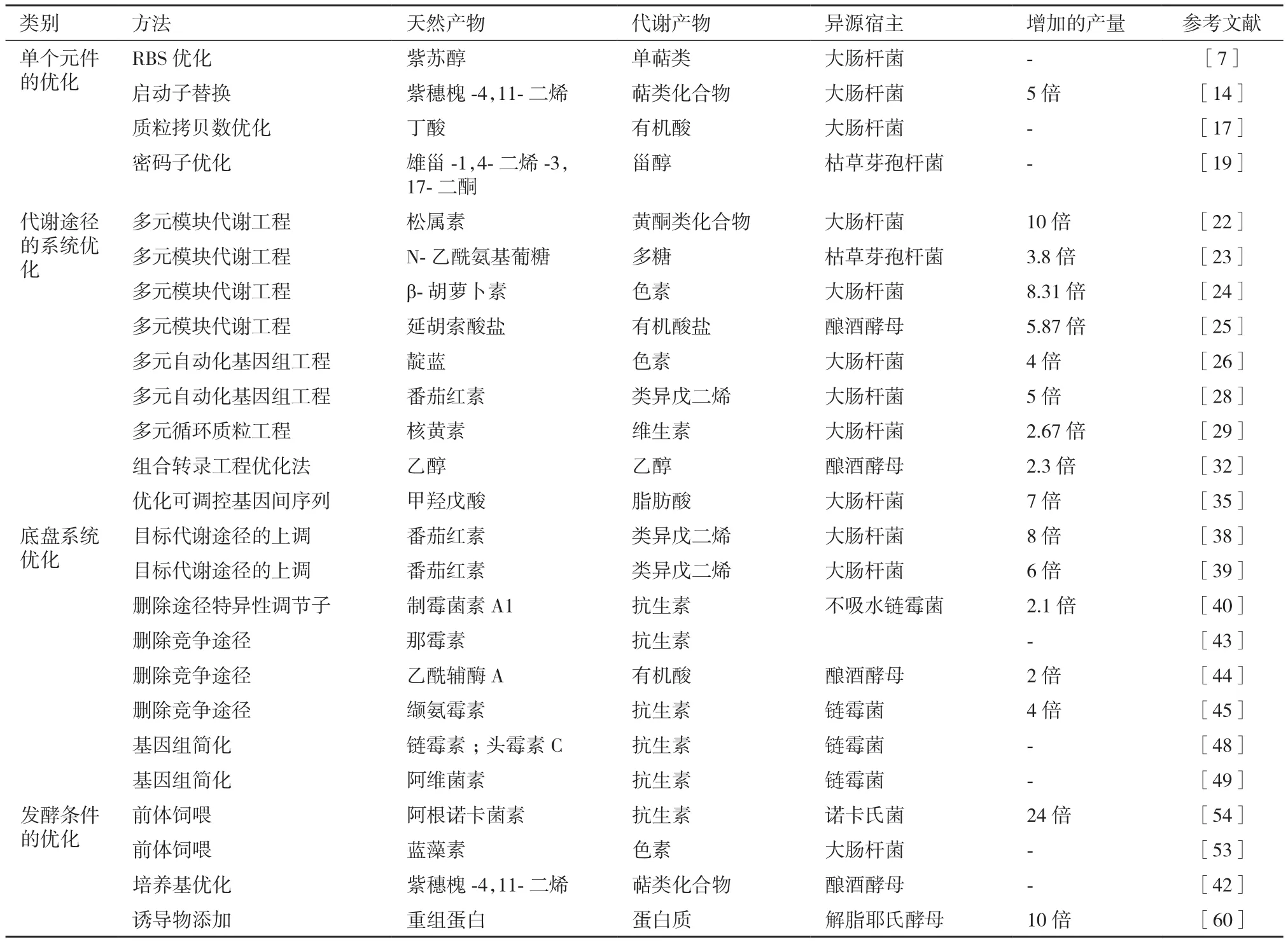
表1 天然产物合成生物学体系的优化研究进展
途径的异源重构常用的工程化宿主有:大肠杆菌、酿酒酵母、链霉菌等,宿主中存在的内源性初生代谢途径能为目标次生代谢产物的合成提供前体物质[10,63]。通过对宿主初生代谢途径进行调节,并与引入的外源代谢途径之间达到平衡,以获得更多的前体物质。删除异源宿主中与目标化合物合成不相关的非必需基因,例如转座子和插入序列基因和次生代谢产物基因簇相关的基因,可以提高细胞的遗传稳定性,减少代谢负担[63]。
目前,途径优化还存在一些瓶颈:首先,途径中不同的基因与方法(例如使用不同拷贝数、启动子和RBS长度)的组合多样,即使是一个小型途径,要找到最优的组合也费时费力。但幸运的是,大型文库的高通量筛选、动态平衡系统和计算机辅助的建模方法进展迅速,极大地促进了途径优化进程。其次,将小规模体系按比例扩大用于目标产物高细胞密度的大规模发酵时,可能会导致系统通量的不平衡,小规模体系所用的发酵条件(如温度、氧气饱和度、细胞密度等)需要进行大幅度的改变,以适应产物工业生产的需要。令人振奋的是,最近出现了多种可用于天然产物生物合成研究的新技术和新方法,如生物传感器用于途径的实时监控,使微生物系统达到动态的平衡[64];CRISPR-Cas9介导的基因组工程技术用于调节基因的强度[65];使用合成支架或融合蛋白实现多个酶空间结构的合理优化[66]、对代谢产物和途径酶进行细胞器水平的区室化,以实现底物和酶的充分利用等[67]。可以预见随着合成生物学理论和技术的快速发展,在不久的将来会开发出更高效的工程工具,用来优化天然产物合成生物学体系,从而实现复杂天然产物的工程化生产。
[1]Jhelum P, Reddy RG, Kumar A, et al. Natural product based novel small molecules with promising neurotrophic, neurogenic and anti-neuroinflammatory actions can be developed as stroke therapeutics[J]. Neural Regen Res, 2016, 11(6):916-917.
[2]Venisetty RK, Ciddi V. Application of microbial biotransformation for the new drug discovery using natural drugs as substrates[J]. Curr Pharm Biotechnol, 2003, 4(3):153-167.
[3] Newman DJ, Cragg GM. Natural products as sources of new drugs from 1981 to 2014[J]. J Nat Prod, 2016, 79(3):629-661.
[4] Jones JA, Toparlak ÖD, Koffas MA. Metabolic pathway balancing and its role in the production of biofuels and chemicals[J]. Curr Opin Biotechnol, 2015, 33:52-59.
[5] Sun H, Zhao D, Xiong B, et al. Engineering Corynebacterium glutamicum for violacein hyper production[J]. Microb Cell Fact, 2016, 15(1):148.
[6] Breitling R, Takano E. Synthetic biology of natural products[J]. Cold Spring Harb Perspect Biol, 2014, doi: 10. 1002/9781118794 623. ch19 .
[7]Alonso-Gutierrez J, Chan R, Batth TS, et al. Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production[J]. Metab Eng, 2013, 19:33-41.
[8]Salis HM, Mirsky EA, Voigt CA. Automated design of synthetic ribosome binding sites to control protein expression[J]. Nat Biotechnol, 2009, 27(10):946-950.
[9]Na D, Lee D. RBSDesigner:software for designing synthetic ribosome binding sites that yields a desired level of protein expression[J]. Bioinformatics, 2010, 26(20):2633-2634.
[10]Ahmadi MK, Pfeifer BA. Recent progress in therapeutic natural product biosynthesis using Escherichia coli[J]. Curr Opin Biotechnol, 2016, 42:7-12.
[11]Santos CN, Koffas M, Stephanopoulos G. Optimization of a heterologous pathway for the production of flavonoids from glucose[J]. Metab Eng, 2011, 13(4):392-400.
[12]Na D, Kim TY, Lee SY. Construction and optimization of synthetic pathways in metabolic engineering[J]. Curr Opin Microbiol, 2010, 13(3):363-370.
[13]Redding-Johanson AM, Batth TS, Chan R, et al. Targeted proteomics for metabolic pathway optimization:application to terpene production[J]. Metab Eng, 2011, 13(2):194-203.
[14]Anthony JR, Anthony LC, Nowroozi F, et al. Optimization of the mevalonate-based isoprenoid biosynthetic pathway in Escherichia coli for production of the anti-malarial drug precursor amorpha-4, 11-diene[J]. Metab Eng, 2009, 11(1):13-19.
[15]Chlebowicz-Sledziewska E, Sledziewski AZ. Construction ofmulticopy yeast plasmids with regulated centromere function[J]. Gene, 1985, 39(1):25-31.
[16]Lian J, Jin R, Zhao H. Construction of plasmids with tunable copy numbers in Saccharomyces cerevisiae and their applications in pathway optimization and multiplex genome integration[J]. Biotechnol Bioeng, 2016, 113(11):2462-2473.
[17]Jawed K, Mattam AJ, Fatma Z, et al. Engineered production of short chain fatty acid in Escherichia coli using fatty acid synthesis pathway[J]. PLoS One, 2016, 11(7):e0160035.
[18]Kong JQ, Wang W, Wang LN. The improvement of amorpha-4, 11-diene production by a yeast-conform variant[J]. J Appl Microbiol, 2009, 106(3):941-951.
[19]Shao M, Sha Z, Zhang X, et al. Efficient androst-1, 4-diene-3, 17-dione production by coexpressing 3-ketosteroid-Δ1-dehydrogenase and catalase in Bacillus subtilis[J]. J Appl Microbiol, 2016, doi:10. 1111/jam. 13336.
[20]Orlenko A, Teufel AI, Chi PB, et al. Selection on metabolic pathway function in the presence of mutation-selection-drift balance leads to rate-limiting steps that are not evolutionarily stable[J]. Biol Direct, 2016, 11:31.
[21]Chen X, Zhu P, Liu L. Modular optimization of multi-gene pathways for fumarate production[J]. Metab Eng, 2016, 33:76-85.
[22]Wu J, Du G, Zhou J, et al. Metabolic engineering of Escherichia coli for(2S)-pinocembrin production from glucose by a modular metabolic strategy[J]. Metab Eng, 2013, 16:48-55.
[23]Liu Y, Zhu Y, Li J, et al. Modular pathway engineering of Bacillus subtilis for improved N-acetylglucosamine production[J]. Metab Eng, 2014, 23:42-52.
[24]Zhao J, Li Q, Sun T, et al. Engineering central metabolic modules of Escherichia coli for improving β-carotene production[J]. Metab Eng, 2013, 17:42-50.
[25]Chen X, Zhu P, Liu L. Modular optimization of multi-gene pathways for fumarate production[J]. Metab Eng, 2016, 33:76-85.
[26]Sharan SK, Thomason LC, Kuznetsov SG, et al. Recombineering:A homologous recombination-based method of genetic engineering[J]. Nat Protoc, 2009, 4(2):206-223.
[27]Carr PA, Wang HH, Sterling B, et al. Enhanced multiplex genome engineering through co-operative oligonucleotide co-selection[J]. Nucleic Acids Res, 2012, 40(17):e132.
[28]Wang HH, Isaacs FJ, Carr PA, et al. Programming cells by multiplex genome engineering and accelerated evolution[J]. Nature, 2009, 460(7257):894-898.
[29]Li Y, Gu Q, Lin Z, et al. Multiplex iterative plasmid engineering for combinational optimization of metabolic pathways and diversification of protein coding sequences[J]. ACS Synth Biol, 2013, 2(11):651-661.
[30]Santos CN, Stephanopoulos G. Combinatorial engineering of microbes for optimizing cellular phenotype[J]. Curr Opin Chem Biol, 2008, 12(2):168-176.
[31]Liu W, Jiang R. Combinatorial and high-throughput screening approached for strain engineering[J]. Appl Microbiol Biotechnol, 2015, 99(5):2093-2104.
[32]Du J, Yuan Y, Si T, et al. Customized optimization of metabolic pathways by combinatorial transcriptional engineering[J]. Nucleic Acids Res, 2012, 40(18):e142.
[33]Klein-Marcuschamer D, Yadav VG, Ghaderi A, et al. De novo metabolic engineering and the promise of synthetic DNA[J]. Adv Biochem Eng Biotechnol, 2010, 120:101-131.
[34]Li XR, Tian GQ, Shen HJ, et al. Metabolic engineering of Escherichia coli to produce zeaxanthin[J]. J Ind Microbiol Biotechnol, 2015, 42(4):627-36.
[35]Pfleger BF, Pitera DJ, Smolke CD, et al. Combinatorial engineering of intergenic regions in operons tunes expression of multiple genes[J]. Nat Biotechnol, 2006, 24(8):1027-1032.
[36]Fatma Z, Jawed K, Mattam AJ, et al. Identification of long chain specific aldehyde reductase and its use in enhanced fatty alcohol production in E. coli[J]. Metab Eng, 2016, 37:35-45.
[37]Santos CN, Xiao W, Stephanopoulos G. Rational, combinatorial, and genomic approaches for engineering L-tyrosine production in Escherichia coli[J]. Proc Natl Acad Sci U S A, 2012, 109(34):13538-13543.
[38]Rodríguez-Villalón A, Pérez-Gil J, Rodríguez-Concepción M. Carotenoid accumulation in bacteria with enhanced supply of isoprenoid precursors by upregulation of exogenous or endogenous pathways[J]. J Biotechnol, 2008, 135(1):78-84.
[39]Zhou Y, Nambou K, Wei L, et al. Lycopene production in recombinant strains of Escherichia coli is improved by knockout of the central carbon metabolism gene coding for glucose-6-phosphate dehydrogenase[J]. Biotechnol Lett, 2013, 35(12):2137-2145.
[40]Cui H, Ni X, Shao W, et al. Functional manipulations of the tetramycin positive regulatory gene ttmRIV to enhance the production of tetramycin A and nystatin A1 in Streptomyces anygroscopicus[J]. J Ind Microbiol Biotechnol, 2015, 42(9):1273-1282.
[41]Paradise EM, Kirby J, Chan R, et al. Redirection of flux through the FPP branch-point in Saccharomyces cerevisiae by down-regulating squalene synthase[J]. Biotechnol Bioeng, 2008, 100(2):371-378.
[42]Westfall P, Pitera DJ, Lenihan JR, et al. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin[J]. Proc Natl Acad Sci USA, 2012, 109(3):E111-118.
[43]Bunet R, Song L, Mendes MV, et al. Characterization and manipulation of the pathway-specific late regulator AlpW reveals Streptomyces ambofaciens as a new producer of Kinamycins[J]. J Bacteriol, 2011, 193(5):1142-1153.
[44]Lian J, Si T, Nair NU, et al. Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains[J]. Metab Eng, 2014, 24:139-149.
[45]Lee DW, Ng BG, Kim BS. Increased valinomycin production in mutants of Streptomyces sp. M10 defective in bafilomycin biosynthesis and branched-chain alpha-keto acid dehydrogenase complex expression[J]. J Ind Microbiol Biotechnol, 2015, 42(11):1507-1517.
[46]Kelkar YD, Ochman H. Genome reduction promotes increase in protein function complexity in bacteria[J]. Genetics, 2013, 193(1):303-307.
[47]Csörgõ B, Nyerges Á, Pósfai G. System-level genome editing in microbes[J]. Curr Opin Microbiol, 2016, 33:113-122.
[48] Komatsu M, Uchiyama T, Omura S, et al. Genome-minimized Streptomyces host for the heterologous expression of secondary metabolism[J]. Proc Natl Acad Sci U S A, 2010, 107(6):2646-2651.
[49]Komatsu M, Komatsu K, Koiwai H, et al. Engineered Streptomyces avermitilis host for heterologous expression of biosynthetic gene cluster for secondary metabolites[J]. ACS Synth Biol, 2013, 2(7):384-396.
[50]Lau J, Frykman S, Regentin R, et al. Optimizing the heterologous production of epothilone D in Myxococcus xanthus[J]. Biotechnol Bioeng, 2002, 78(3):280-288.
[51] Bonartsev AP, Zharkova II, Yakovlev SG, et al. Biosynthesis of poly(3-hydroxybutyrate)copolymers by Azotobacter chroococcum 7B:A precursor feeding strategy[J]. Prep Biochem Biotechnol, 2016:1-12.
[52]Saha SP, Patra A, Paul AK. Incorporation of polyethylene glycol in polyhydroxyalkanoic acids accumulated by Azotobacter chroococcum MAL-201[J]. J Ind Microbiol Biotechno, 2006, 33(5):377-383.
[53]Du J, Li L, Zhou S. Enhanced cyanophycin production by Escherichia coli overexpressing the heterologous cphA gene from a deep sea metagenomic library[J]. J Biosci Bioeng, 2016, pii:S1389-1723(16)30208-0.
[54]Dhakal D, Chaudhary AK, Yi JS, et al. Enhanced production of nargenicin A1 and creation of a novel derivative using a synthetic biology platform[J]. Appl Microbiol Biotechnol, 2016, 100(23):9917-9931.
[55]Ujor V, Agu CV, Gopalan V, et al. Glycerol supplementation of the growth medium enhances in situ detoxification of furfural by Clostridium beijerinckii during butanol fermentation[J]. Appl Microbiol Biotechnol, 2014, 98(14):6511-6521.
[56]Yi JS, Kim MS, Kim SJ, et al. Effects of sucrose, phosphate, and calcium carbonate on the production of pikromycin from Streptomyces venezuelae[J]. J Microbiol Biotechnol, 2015, 25(4):496-502.
[57]Cheigh CI, Choi HJ, Park H, et al. Influence of growth conditions on the production of a nisin-like bacteriocin by Lactococcus lactis subsp. lactis A164 isolated from kimchi[J]. J Biotechnol, 2002, 95(3):225-235.
[58]Chander H, Batish VK, Babu S, et al. Amine production by Streptococcus lactis under different growth conditions[J]. Acta Microbiol Pol, 1988, 37(1):61-64.
[59]Ran H, Wu J, Wu D, et all. Enhanced production of recombinant Thermobifida fusca isoamylase in Escherichia coli MDS42[J]. Appl Biochem Biotechnol, 2016, 180(3):464-476.
[60]Sassi H, Delvigne F, Kar T, et al. Deciphering how LIP2 and POX2 promoters can optimally regulate recombinant protein production in the yeast Yarrowia lipolytica[J]. Microb Cell Fact, 2016, 15(1):159
[61]Alper H, Stephanopoulos G. Global transcription machineryengineering:a new approach for improving cellular phenotype[J]. Metab Eng, 2007, 9(3):258-267.
[62]Carbonell P, Currin A, Jervis AJ, et al. Bioinformatics for the synthetic biology of natural products:integrating across the Design-Build-Test cycle[J]. Nat Prod Rep, 2016, 33(8):925-932.
[63]Billingsley JM, DeNicola AB, Tang Y. Technology development for natural product biosynthesis in Saccharomyces cerevisiae[J]. Curr Opin Biotechnol, 2016, 42:74-83.
[64]Tsao CY, Hooshangi S, Wu HC, et al. Autonomous induction of recombinant proteins by minimally rewiring native quorum sensing regulon of E. coli[J]. Metab Eng, 2010, 12(3):291-297.
[65]Pankowicz FP, Barzi M, Legras X, et al. Reprogramming metabolic pathways in vivo with CRISPR/Cas9 genome editing to treat hereditary tyrosinaemia[J]. Nat Commun, 2016, 7:12642.
[66]Dueber JE, Wu GC, Malmirchegini GR, et al. Synthetic protein scaffolds provide modular control over metabolic flux[J]. Nat Biotechnol, 2009, 27(8):753-759.
[67] Farhi M, Marhevka E, Masci T, et al. Harnessing yeast subcellular compartments for the production of plant terpenoids[J]. Metab Eng, 2011, 13(5):474-481.
(责任编辑 马鑫)
Optimization Strategies for Synthetic Biological Systems of Natural Products
KUANG Xue-jun1ZOU Li-qiu1SUN Chao1CHEN Shi-lin2
(1. Institute of Medicinal Plant Development,China Academy of Medical Sciences and Peking Union Medical College,Beijing 100193;2. Institute of Chinese Materia Medica,China Academy of Chinese Medical Sciences,Beijing 100700)
Natural products have become an important source of new drugs. However,there are some deficiencies while producing complex natural products by classical organic synthesis,one measure to efficiently make up those deficiencies is to apply synthetic biology in the biosynthesis of natural product,i.e.,designing and reconstructing efficient biosynthetic pathways of target compounds,re-engineering it in host cells,and producing target compounds by fermentation. However,achieving the yield of target product to the industrial level through technology of synthetic biology is still very challenging though there have been advances in synthetic biology. Here,we review various pathway optimization strategies for the synthetic biological systems of natural products. Via optimization technologies of fine-tuning individual component,exogenous metabolic pathways,the chassis systems,and fermentation conditions,the synthetic biological system may be optimized,and the yield of target product may be maximized,thus providing continuous,stable and economical raw material supplies for the manufacture of complex natural products while source is rare,and promoting the research and development of new drugs similar to natural products
natural products;synthetic biology;optimization;chassis
10.13560/j.cnki.biotech.bull.1985.2017.01.005
2016-10-22
国家自然科学基金面上项目(81273485,81573704)
匡雪君,女,硕士研究生,研究方向:天然药物的代谢工程与合成生物学;E-mail:kuangxuejun@126.com
孙超,男,博士,教授,研究方向:基于组学分析的药用植物有效成分生物合成途径解析及天然药物的代谢工程与合成生物学;E-mail:csun@implad.ac.cn;陈士林,男,博士,E-mail:slchen@icmm.ac.cn
