水稻糙米重金属镉(Cd2+)含量的QTL定位分析
2017-02-15郭小蛟蒋开锋杨乾华游书梅曹应江李昭祥万先齐郑家奎四川省农业科学院水稻高粱研究所农业部西南水稻生物学与遗传育种重点实验室四川德阳68000国家水稻改良中心泸州分中心四川泸州64600
郭小蛟,杨 莉,*,张 涛,,蒋开锋,,杨乾华,游书梅,曹应江,李昭祥,罗 婧,高 磊,万先齐,郑家奎,(.四川省农业科学院水稻高粱研究所/农业部西南水稻生物学与遗传育种重点实验室,四川 德阳 68000;.国家水稻改良中心泸州分中心,四川 泸州 64600)
水稻糙米重金属镉(Cd2+)含量的QTL定位分析
郭小蛟1,杨 莉1,2*,张 涛1,2,蒋开锋1,2,杨乾华1,游书梅1,曹应江1,李昭祥1,罗 婧1,高 磊1,万先齐2,郑家奎1,2
(1.四川省农业科学院水稻高粱研究所/农业部西南水稻生物学与遗传育种重点实验室,四川 德阳 618000;2.国家水稻改良中心泸州分中心,四川 泸州 646100)
本研究利用日本晴与泸恢99构建的188个重组自交系群体,根据已构建的遗传连锁图谱,采用混合线性模型方法,在高镉含量环境A和低镉含量环境B进行糙米Cd2+含量的QTL定位分析。在A环境中检测到2个控制糙米Cd2+含量的QTLs,分别位于第7、8染色体,贡献率分别为29.8%、8.87%;在B环境中检测到1个控制糙米Cd2+含量的QTL,位于第7染色体,贡献率为7.35%;A、B两个环境下的增效基因均来自于亲本泸恢99。试验表明:镉浓度对该基因的表达有影响,糙米Cd2+含量QTL基因更易在Cd2+高浓度环境中表达,因此,Cd2+低吸收水稻材料创制及选育应在Cd2+含量较高的土壤中进行。
水稻;糙米Cd2+含量;QTL定位
水稻(Oryzasativa)是世界上最主要的粮食作物,有50%以上的人口以稻米为主食。近年来,随着工业“三废”排放量的增加,各种化学产品、农药、化肥的广泛无节制使用,使农田土壤中有毒重金属含量急剧增加,我国面临的土壤环境及粮食安全问题日益突出。中国水稻研究所与农业部2010年的研究称,全国受到镉污染的耕地涉及11个省份25个地区。在湖南、江西等长江以南地带,这一问题更加突出。南京农大潘根兴团队在全国多个县级以上市场随机采购样品,结果表明,10%左右的市售大米镉超标。2008年以来,相继出现了湖南、广州等地“镉大米”事件,在社会上引起了强烈反响。重金属污染特别是Cd2+污染已严重危及食物安全,因此引起世界各国的广泛重视。大量研究表明,不同水稻品种由于遗传上的差异,在对稻田重金属元素的吸收和分配上存在很大差异,这种差异不仅存在于种间(不同种和属),而且在种内(不同变种或品种)也存在[1、2、3、4、5、6、7、8]。针对重金属污染土壤的治理方法,国内外学者作了大量研究,取得了一系列的成功技术,但这些方法在有效性、持久性及经济性方面难以达到预期效果。日本冈山大学教授马建锋等[9]研究发现,种植不含Nramp5基因的水稻,可以有效地解决水稻中Cd2+富集的问题,但是却破坏了摄取锰元素的能力,导致产量比正常水稻降低了近90%,这在生产上显然是不能接受的。而通过品种选择,将作物食用部位的重金属浓度控制在允许范围被认为是治理污染的有效途径,并在向日葵(Helianthus annuus)和硬质小麦上成功应用[10、11、12],这为该方面的深入研究奠定了基础。因此,针对土壤重金属污染,开展作物耐重金属污染基因的发掘、分子标记及利用研究,开展作物耐重金属污染品种的筛选、鉴定和选育研究,必将是有效、持久和经济的重金属污染土壤治理策略的发展趋势。近年来,有关糙米中Cd2+含量的QTL定位研究也取得了一定的进展[13、14、15],但总的来说定位的QTL还很少,且贡献率均不大,还不足以用于分子标记辅助选择育种。本研究将利用日本晴×泸恢99构建的RIL群体分析控制糙米中Cd2+含量的QTL,旨在发掘更多有利基因,为分子标记辅助选择Cd2+低吸收品种提供一定的理论和技术指导。
1 材料与方法
1.1 试验材料
以测序品种日本晴(粳稻)为母本,与我所选育的恢复系泸恢99(籼稻)杂交,F2代始经单粒传法获得369个株系的F8代重组自交系群体(Recombinant inbred lines,RIL),选取188个株系为试验材料。
1.2 田间试验与表型分析
2012年夏季,选取2个土壤Cd2+含量不同的田块进行了试验,其中A环境土壤Cd2+含量为1.45mg/kg,B环境土壤Cd2+含量为0.15mg/kg(国家规定,pH值≤7.5土壤安全标准为0.3mg/kg,pH值 ≥7.5时为0.6mg/kg,大米中的安全镉含量为0.2mg/kg)。2个亲本及188个株系于4月1日播种,5月8日移栽。每个株系种植4行,每行10株,株行距16.67cm×26.67cm,两环境田间肥水管理、病虫害防治同常规大田管理。
正季稻成熟时,收取2、3行各5株种子,自然风干后储存3个月,送四川省农业科学院分析测试中心,采用GB/T5009.15-2003方法检测糙米中的Cd2+含量。
1.3 数据分析和QTL定位
用SPSS统计软件进行表型数据的统计分析。根据已构建的遗传连锁图谱(赵建国,2013)[16],利用基于混合线性模型[17]的QTL Network2.0[18]软件进行QTL分析。以P = 0.005为统计检测阈值, 即当标记的P 值小于统计检测阈值时, 认为该标记处存在1个QTL;最后将检测到的所有QTL 以及它们之间的上位性互作以及与环境的互作整合到一个全QTL 模型中, 用基于Gibbs 抽样的Bayesian 方法估计遗传效应。QTL的命名参照McCouch[19]水稻QTL命名方法。
2 结果分析
2.1 亲本及群体表型
在2个不同Cd2+含量的土壤环境下(表1),双亲糙米Cd2+含量性状差异均达极显著水平,其中籼稻泸恢99糙米Cd2+含量明显比粳稻日本晴高。A环境下,糙米Cd2+含量性状在RIL群体中表现为连续变异,株系间变异幅度很大,变异范围在0.18~5.54mg/kg之间,差异明显。大多数株系性状值居于双亲中间,部分株系呈超亲优势,个别株系甚至比土壤背景值高出几倍,糙米Cd2+含量性状表现正态分布(图1),表明糙米Cd2+含量是受多基因控制的数量性状。B环境中性状亦呈连续变异,株系间的变异范围为0.0039~0.27mg/kg,幅度变较大,但大多数株系糙米Cd2+含量很低,大约60%的株系Cd2+含量集中在0.0039~0.037mg/kg之间,株系间差异不明显,性状表现出一定的偏分离,不符合正态分布,这可能与B环境土壤Cd2+含量低有关。从本试验的结果来看,土壤背景值对水稻籽粒Cd2+含量这一性状的表达有影响,Cd2+含量高的土壤背景下更有利于鉴别株系间的性状差异。
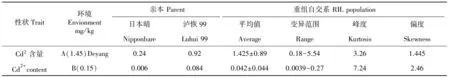
表1 双亲及群体在两种环境下的性状表现值
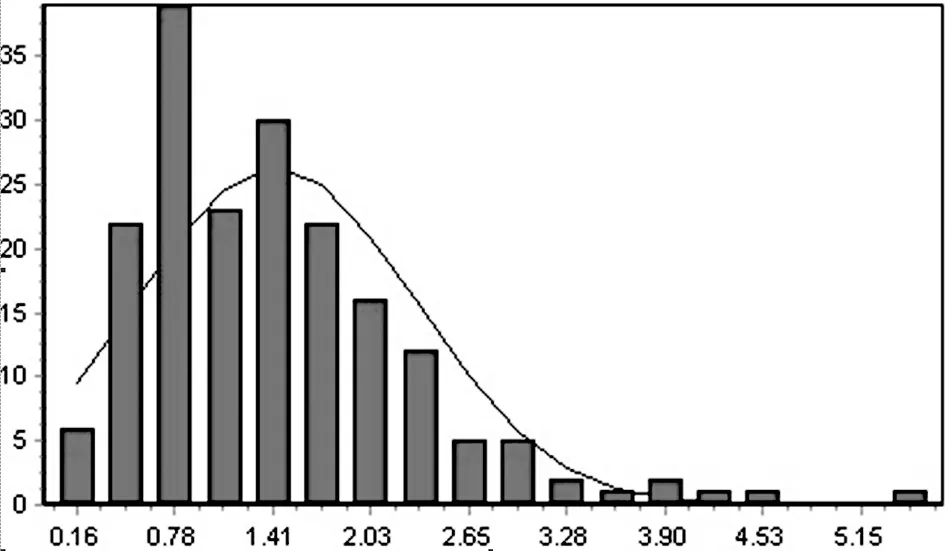
图1 A环境188个家系糙米Cd2+含量的正态分布图
2.2 QTL定位分析
根据已构建的遗传连锁图谱[16],该图谱包含有覆盖水稻全基因组12条染色体的207个DNA分子标记。其中SSR标记136个,STS标记24个,Indel标记47个。连锁图谱全长为2397cM,标记间平均距离为12.29cM。在A环境中检测到2个糙米Cd2+含量的QTL,即位于第7、8染色体上的qCCBR-7-1和qCCBR-8,均表现出增效作用,增效基因均来自泸恢99。贡献率分别为29.8%和8.87%,加性效应分别为-0.4943和-0.2154。同时还检测到qCCBR-7-1和qCCBR-8之间存在上位性互作,但效应值较低仅为2.37%。B环境中检测到1个QTL,qCCBR-7-2位于第7染色体SSR标记RM214与RM11之间,与A环境中检测到qCCBR-7-1位置相近(图2),贡献率为7.35%,增效基因来自于亲本泸恢99。
3 讨论
3.1 QTL定位结果比较
数量性状易受环境影响,同一数量性状的QTL在不同定位群体、不同环境及不同年份等QTL检测结果均会有所不同。Ishikawa等[13]利用含39个片段代换系的图谱检测到位于第3、6、8染色体上的3个控制糙米中Cd2+含量的QTLs。沈希宏等[14]利用协青早B×中恢9308构建的RIL群体在不同的土壤环境下分别检测到2个、1个控制糙米中Cd2+含量的QTLs,分别位于第5、7和11染色体上,贡献均在9%左右。陈志德[15]利用韭菜青×IR26的重组自交系群体(RIL),在5 mg/kg Cd2+胁迫下进行糙米Cd2+浓度的QTL定位,在第11染色体上检测到2个控制糙米Cd2+浓度的QTL。本研究利用日本晴与R99的RIL群体,在土壤Cd2+浓度较高的A环境中在第7和第8染色体上各检测到了1个QTL,B环境中第7染色体上检测到1个QTL。与前人的研究比较发现,是3个新的糙米Cd2+含量QTL位点。位于第7染色体的2个QTL在物理位置上比较近,qCCBR-7-1(镉较高浓度)的贡献率较大29.8%,沈希宏等[14]在第7染色体上也定位了1个控制糙米Cd2+含量QTL位点,可能该区域是控制糙米Cd含量的基因区域。而qCCBR-7-2(低浓度)贡献率小,可能是因为糙米Cd含量QTL基因在土壤低Cd浓度下不易表达。因此,在此研究基础上,进一步开展qCBBR-7-1基因的精细定位和克隆研究,开展水稻与Cd2+的分子生物学研究,探明控制水稻糙米Cd含量基因的遗传、分子和生理生化机制,发掘更多有利基因,最终利用分子标记辅助选择或转基因技术改良杂交水稻不育系和恢复系亲本,选育水稻籽粒Cd2+低吸收品种,对于解决我国面临的粮食生产安全问题具有理论和技术指导意义。

表2 QTL定位结果及遗传参数估算
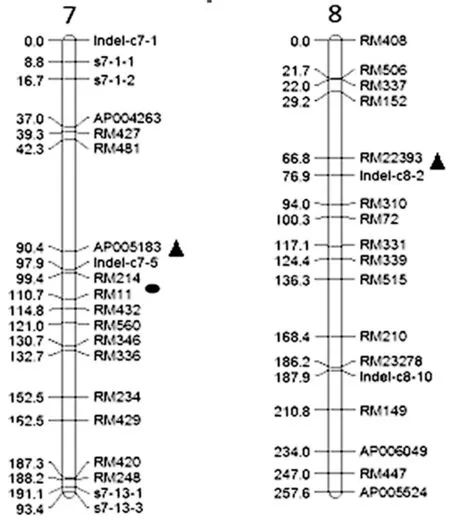
图2 控制糙米中Cd2+含量的QTL图谱(▲环境A ●环境B)
3.2 性状与环境的相关性
沈希宏等[14]在不同Cd2+污染程度土壤中,检测到了显著的QTL与环境互作效应,说明环境对基因表达有显著影响。本实验中,环境A和环境B土壤Cd2+浓度分别为1.45 mg/kg和0.15mg/kg。A环境中出现了比亲本高数倍的株系,株系间的性状差异明显,而在B环境中株系间差异不如A环境显著,尤其是吸收量低的株系所占比例较大,这很可能与B环境的土壤Cd2+浓度较低导致了糙米Cd2+含量QTL基因不易表达有关。从本研究结果看,土壤Cd2+浓度高更有利于Cd2+含量QTL基因的表达。杨莉[8]曾利用不完全双列杂交材料在较高镉环境下研究了镉的遗传效应,结果表明,籽粒低Cd2+杂交稻组合的选育,双亲的遗传改良和组合的评价筛选都很重要,不育系改良的效果比恢复系好。因此,Cd2+低吸收材料(品种)的鉴定及选育最好选择在Cd2+含量较高的环境中进行。
4 结论
(1)发现3个新的糙米Cd2+含量QTL位点,其中第7染色体的2个QTL在物理位置上比较近,qCCBR-7-1(镉较高浓度)的贡献率较大29.8%,值得做进一步研究。第7染色体上可能存在一个Cd2+含量基因区域。
(2)土壤Cd2+浓度高更有利于Cd2+含量QTL基因的表达,因此Cd2+低吸收材料(品种)的鉴定及选育最好选择在Cd2+含量较高的环境中进行。
[1]王荣凯,龚惠群.两种基因型水稻对环境Cd2+吸收与再分配差异性比较研究[J].农业环境保护,1996,15:145-149,176-176.
[2]吴启堂,陈卢,王广寿.水稻不同品种对Cd2+吸收累积的差异和机理研究[J].生态学报,1999,19 :104-107.
[3]李花粉,张福锁,李春俭,等.Fe对不同品种水稻吸收Cd的影响[J].应用生态学报,1998,9(1):110-112.
[4]刘敏超,李花粉,夏立江,等.根表铁锰氧化物胶膜对不同品种水稻吸收Cd2+的影响[J].生态学报,2001,20:592-596.
[5]李坤全,刘建国,陆小龙,等.水稻不同品种对Cd2+吸收与分配的差异[J].农业环境科学学报,2003,22:503-504.
[6]Lin JG,Li KQ,Liang JS,etal.Zhu QS Interaction of Cd and five mineral nutrients for uptake and accumulation in different rice cultivars and genotypes[J].FieldCropsResearch,2003a,83:271-281.
[7]Lin JG, Li KQ,Liang JS,etal.Zhu QSLead toxicity ,uptake,and translocation in different rice cultivars[J].PlantScience,2003b,165:793-862.
[8]杨莉 ,郑家奎,蒋开锋,等.杂交水稻籽粒Cd2+和砷含量的遗传效应分析[J].杂交水稻,2010,5:80-83.
[9]Akimasa Sasaki,Naoki Yamaji,Kengo Yokoshoand,etal. Nramp5 Is a Major Transporter Responsible for Manganese and Cadmium Uptake in Rice[J].ThePlantCellMay,2012,112.
[10]Li YM, Channey LR, Schneiter AA (1995) Genotypic variation in kernel cadmium concentration in sun-flower germplasm under varying soil conditions[J].Crop Science,35: 137-141.
[11]Penner GA, Clarke J,Bezte LJ,etal. Iden-tification of PAPD markers linked to a gene gov-erning cadmium uptake in durum wheat[J].Genome,1995,38: 543-547.
[12]Wang KR. Tolerance of cultivated plants tocadmium and their utilization in polluted farmland soils[J]. Acta Biotechnology,2002,22:189-198.
[13]Satoru Ishikawa, Noriharu Ae.Masahiro Yano Chromosomal regions with quantitative trait loci controlling cadmium concentration in brown rice (Oryza sativa)[J].Newphytologist.,2005,168(2):345-350.
[14]沈希宏,曹立勇,邵国胜,等.水稻籽粒中5种微量元素的QTL定位[J].分子植物育种,2008,6(6)1061-1067.
[15]陈志德,王州飞, 贺建波, 等.水稻糙米重金属 Cd2+含量的 QTL 分析[J].遗传,2009,31(11)1135-1140.
[16]赵建国, 蒋开锋,杨莉,等.水稻产量相关性状QTL定位[J].中国水稻科学,2013,27(4):344-352.
[17]Wang D L, Zhu J, Li Z L,etal. Mapping QTLs with epistatic effects and QTL × environment .interactions by mixed linear model approaches[J].TheoreticalandAppliedGenetics, 1999, 99(7): 1255-1264.
[18]Yang J, Zhu J. Predicting superior genotypes in multiple environments based on QTL effects[J].TheoreticalandAppliedGenetics, 2005, 110(7): 1268-1274.
[19]Mccouch S R, Teytelman L, Xu Y,etal. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.)[J].DNAresearch,2002, 9(6): 199-207.
2016-11-19
四川省科技支撑计划项目(2015SZ0048)、四川省财政现代农业技术创新与示范项目。
郭小蛟(1988-),女,研究实习员,硕士,专业方向为水稻遗传育种。E-mail:gxiaojiao@163.com。*为通讯作者。
