Smad3对非小细胞肺癌A549细胞和宫颈癌HeLa细胞迁移的影响
2017-02-15韦荣飞李梦媛杨星九朱瑞敏徐大模
韦荣飞,李梦媛,杨星九,朱瑞敏,徐大模,高 苒
(中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,北京 100021)
Smad3对非小细胞肺癌A549细胞和宫颈癌HeLa细胞迁移的影响
韦荣飞,李梦媛,杨星九,朱瑞敏,徐大模,高 苒*
(中国医学科学院医学实验动物研究所,北京协和医学院比较医学中心,北京 100021)
目的 过表达、敲低或抑制Smad3的活性,探讨Smad3对非小细胞肺癌A549细胞和宫颈癌HeLa细胞迁移的影响。方法 设计扩增Smad3基因全长编码序列cDNA的引物,通过分子克隆技术构建pCMV-Myc-Smad3过表达载体,同时设计并合成靶向Smad3的siRNA序列,在A549和HeLa细胞中过表达或敲低Smad3,或者利用Smad3的抑制剂SIS3 2 μM处理细胞,采用细胞划痕的方法研究Smad3对A549和HeLa细胞迁移的影响。结果 利用分子克隆技术成功构建pCMV-Myc-Smad3过表达载体。过表达Smad3促进A549和HeLa细胞的迁移。敲低Smad3抑制A549和HeLa细胞的迁移。SIS3抑制Smad3的活性导致A549和HeLa细胞的迁移速率变慢。结论 Smad3促进A549和HeLa细胞的迁移。
Smad3;过表达载体;RNA干扰;SIS3;细胞迁移
TGF-β(transforming grpwth factor beta,转化生长因子β)信号通路是调节细胞生长的重要的信号通路,机体对TGF-β信号通路响应的敏感性降低,将会导致肿瘤的形成[1]。经典的TGF-β信号通路由TGF-β I型和II型受体(TGFBR1,TGFBR2)介导,TGFBR1和TGFBR2与TGF-β结合后能够磷酸化下游R-Smads(receptor-activated Smads)蛋白,包括Smad2和Smad3[2,3]。激活的R-Smads蛋白随后与Smad4结合并转移至细胞核中调控基因的转录[4-6]。正常的生理条件下,在多种组织器官中,包括乳房组织,Smad3能够抑制细胞的增殖并促进细胞凋亡,而在乳腺癌晚期,Smad3能够促进乳腺癌细胞的转移[7]。2015年,Rodney B Luwor等在MolecularCancer杂志发表文章揭示,与非迁移细胞相比,迁移的人乳腺癌MDA-MB-231细胞中,Smad3具有更高的活性[8]。除了乳腺癌细胞,Smad3是否也能影响其他类型的癌细胞的迁移呢?为探讨该科学问题,我们通过构建pCMV-Myc-Smad3过表达载体,检测过表达Smad3后对宫颈癌HeLa细胞和非小细胞肺癌A549细胞迁移的影响。同时,通过RNAi干扰及Smad3抑制SIS3处理细胞敲低Smad3的蛋白水平或抑制Smad3的活性,进一步验证Smad3对宫颈癌HeLa细胞和非小细胞肺癌A549细胞迁移的影响。
1 材料和方法
1.1 细胞及主要试剂
载体构建PCR引物由上海维基生物科技有限公司合成;Taq polymerase、dNTP购自宝生物工程(大连)有限公司;限制性内切酶Sal I、Not I及T4 DNA连接酶购自美国NEB公司;感受态细胞DH5α购自天根生化科技(北京)有限公司;质粒提取盒购自德国Qiagen公司;胎牛血清、DMEM、胰酶购自美国Gibco公司;RIPA裂解液购自Sigma公司;蛋白Marker及发光底物购自Thermo公司;Lipofectamine 2000及RNAiMax转染试剂购自Life Technologies公司。HEK-293T、HeLa和A549细胞系为本实验室所冻存。
1.2 实验方法
1.2.1 pCMV-Myc-Smad3过表达载体的构建
1.2.1.1 Smad3 DNA目的片段的获得
提取HeLa细胞的总RNA,并进行cDNA扩增;根据CDS序列(NM_005902.3)设计含有Sal I和Not I限制性内切酶位点的引物序列;以反转录的Smad3 cDNA作为模板,扩增Smad3 DNA目的片段,条件为95℃预变性5 min,95℃ 30 s,60℃ 30 s,72℃ 1 min 20 s,共35个循环,72℃ 5 min。将PCR产物进行琼脂糖凝胶电泳鉴定,紫外灯下切割含有目的条带的凝胶块,利用凝胶回收试剂盒回收目的片段。
1.2.1.2 酶切
利用Sal I和Not I限制性内切酶对PCR获得的Smad3 DNA目的片段及pCMV-Myc-vector进行酶切,酶切条件为37℃ 过夜。将酶切产物进行琼脂糖凝胶电泳鉴定,紫外灯下切割含有目的条带的凝胶块,利用凝胶回收试剂盒回收目的片段。
1.2.1.3 连接及转化
利用T4 DNA连接酶将酶切回收后的Smad3 DNA片段和线性pCMV-Myc-vector进行连接,连接条件为16℃过夜。连接产物转化感受态DH5α细胞,在含有氨苄霉素的LB平板上筛选克隆。挑取阳性克隆,过夜摇菌扩增。
1.2.1.4 质粒提取、酶切鉴定及测序
利用质粒提取试剂盒提取阳性菌中的质粒。采用Sal I和Not I限制性内切酶对重组质粒pCMV-Myc-Smad3进行双酶切鉴定,并以1%琼脂糖凝胶电泳验证片段大小,同时对重组质粒进行基因测序鉴定。
1.2.1.5 HeLa细胞和A549细胞的培养
HEK-293T、HeLa和A549细胞培养基为含10%胎牛血清、1%双抗的DMEM高糖培养基。培养于37℃,5% CO2的细胞培养箱。
1.2.1.6 质粒的表达检测
用胰酶将HEK-293T细胞消化成单细胞悬液并对24孔板进行铺板,24 h后,通过Lipofectamine 2000将0.8 μg的pCMV-Myc-vector和重组质粒pCMV-Myc-Smad3分别转染。24 h后,RIPA裂解液裂解细胞,加入等体积的2×loading buffer,沸水煮15 min,得到蛋白样品。随后利用Myc标签抗体进行Western blot检测重组质粒pCMV-Myc-Smad3的表达情况。
1.2.2 划痕实验
用胰酶将HeLa和A549细胞消化成单细胞悬液并对6孔板进行铺板,24 h后,通过Lipofectamine 2000或RNAiMax转染试剂将3 μg的重组质粒pCMV-Myc-Smad3或Smad3的siRNA分别进行转染。40 h后,用20 μL枪头沿直线对6孔板中长满的细胞进行划痕,0 h、24 h及48 h时分别拍照记录细胞的迁移情况。Smad3抑制剂SIS3(工作浓度2 μM)加入细胞培养基中培养细胞,划痕及记录方法同上。
1.3 统计学方法
细胞迁移划痕实验每个时间点随机选取5个视野拍照,并利用ZEN 2软件测量划痕间距,以时间点0 h划痕宽度作为标准即100%,其他时间点划痕间距与0 h划痕间距的比值即为该时间点划痕两侧的相对距离(百分比),相对距离越小,则说明相对迁移速率越大。利用Graphpad Prism 7对结果进行t检验,P< 0.05表示有统计学意义。
2 结果
2.1 pCMV-Myc-Smad3过表达载体的构建
Smad3蛋白由425个氨基酸组成,其主要包含两个结构域:MH1和MH2结构域(图1A)。MH1和MH2对于Smad3正常生物学功能的发挥至关重要。我们以cDNA作为模板,用引物Smad3-F/R(表1)进行扩增,选择Sal I和Not I分别为上下游内切酶切割位点(图1B),得到与预期片段大小相符的特异性片段(图2A)。利用Sal I和Not I对PCR产物进行酶切后,通过T4 DNA连接酶将酶切产物连接至pCMV-Myc载体上。经测序比对及图2B酶切鉴定结果显示,Smad3 DNA全长片段已成功插入pCMV-Myc载体上。如图2C,Western blot结果显示,与HEK-293T细胞中只转染了Myc-vector的对照组相比,转染Myc-Smad3过表达质粒24 h后,在约55 kDa大小处检测到Smad3的过表达条带,表明pCMV-Myc-Smad3过表达载体构建成功。

表1 pCMV-Myc-Smad3过表达载体构建所需引物
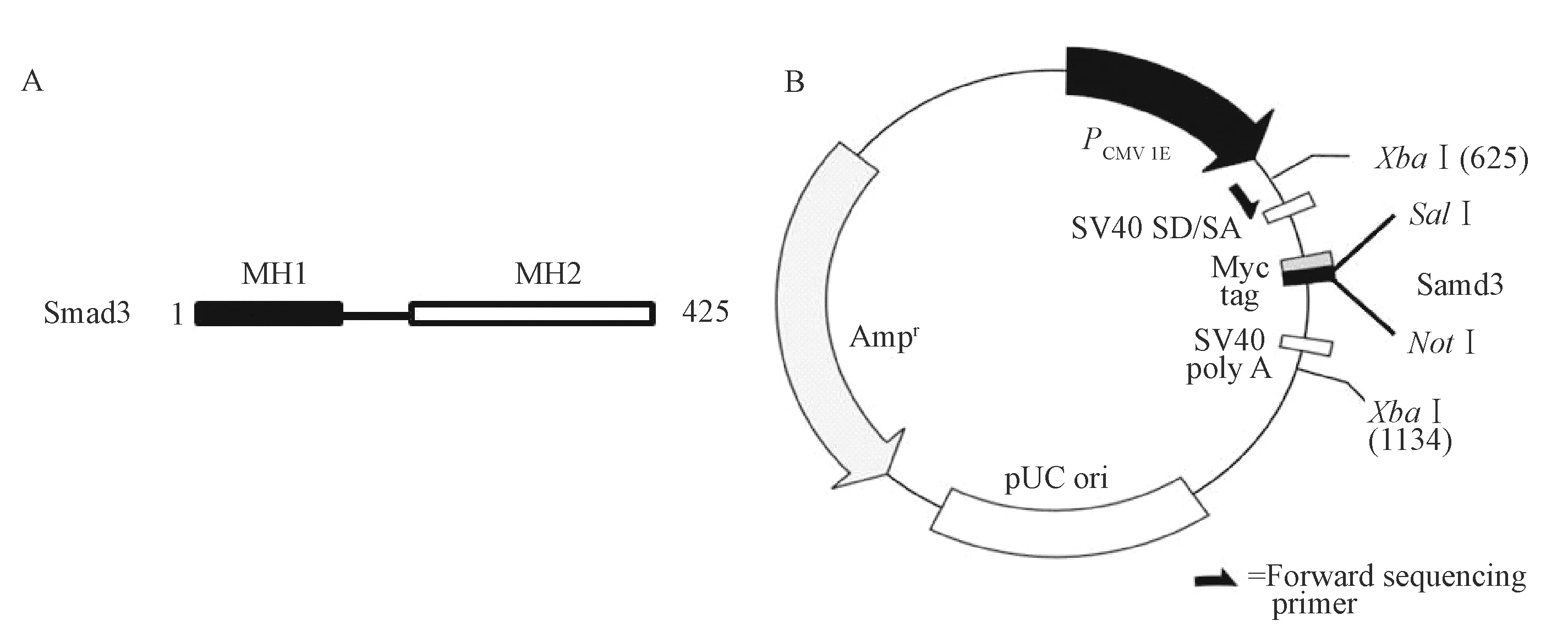
注:(A)为Smad3蛋白的结构示意图,主要包括MH1和MH2结构域;(B)为pCMV-Myc-Smad3过表达载体构建示意图,其中,选取Sal I和Not I分别为上下游限制性内切酶切割位点。图1 Smad3蛋白和pCMV-Myc-Smad3过表达载体构建示意图 Note.(A)indicates the structure diagram of Smad3, including MH1 and MH2 domain;(B)indicates the plasmid profile of pCMV-Myc-Smad3.Fig.1 Structure diagram of Smad3 and pCMV-Myc-Smad3 plasmid construction.

注:(A)为利用引物Smad3-F/R扩增Smad3 DNA片段;(B)为Sal I/Not I酶切鉴定pCMV-Myc-Smad3质粒;(C)为HEK-293T细胞中转染pCMV-Myc-Smad3过表达质粒,Myc-vector作为阴性对照,利用Myc标签抗体进行Western blot检测Smad3的过表达情况。图2 pCMV-Myc-Smad3过表达载体的构建及蛋白表达检测Note.(A)indicates PCR for amplification of Smad3 DNA;(B)indicates the restriction enzyme digestion for pCMV-Myc-Smad3 by Sal I + Not I;(C)indicates the detection of protein expression level of pCMV-Myc-Smad3 in HEK-293T by Western blot.Fig.2 Construction of pCMV-Myc-Smad3 plasmid
2.2 Smad3抑制剂SIS3对非小细胞肺癌A549细胞和人宫颈癌HeLa细胞迁移的影响
SIS3是Smad3的抑制剂,其能有效抑制Smad3的磷酸化,进而阻止Smad3进入细胞核参与基因的转录调控。为研究SIS3对细胞迁移的影响,首先,在A549细胞中检测SIS3对Smad3的抑制作用,以证明SIS3的活性。如图3所示,SIS3能够有效抑制Smad3的磷酸化。与DMSO处理组相比,含2 μM SIS3的DMEM培养基培养细胞,划痕后0、24、48 h,A549(图4A)和HeLa(图4B)细胞迁移结果显示,SIS3能够有效抑制细胞的迁移。

注:上图分别为2 μM SIS3处理A549细胞(A)和HeLa细胞(B)后进行划痕,0、24、48 h时拍照观察细胞的迁移情况并进行统计学分析,DMSO处理组作为对照组。与对照组比较,*P<0.05,**P<0.01,***P < 0.001。图4 Smad3抑制剂SIS3能够有效抑制A549细胞和HeLa细胞的迁移Note. The above results indicate the inhibitor of Smad3, SIS3, significantly inhibits cell migration of A549 (A) and HeLa (B) cells;*P < 0.05,**P < 0.01,***P < 0.001,compared with the normal group.Fig.4 The inhibitory effect of SIS3 on cell migration of A549 and HeLa cells

注:上图为Western blot检测A549细胞中Smad3抑制剂SIS3对Smad3活性(磷酸化)的抑制作用。图3 Smad3抑制剂SIS3对Smad3活性的抑制效应Note. The above result shows the inhibitor of Smad3, SIS3, has significant inhibitory effect on Smad3 activity. Fig.3 The inhibitory effect of SIS3 on Smad3
2.3 敲低或过表达Smad3对A549和HeLa细胞迁移的影响
如图5所示,与对照组相比,通过RNAi干扰技术敲低A549(图5A)和HeLa(图5B)细胞中内源的Smad3后,划痕后0、24、48 h细胞迁移结果显示,敲低Smad3明显抑制细胞的迁移。相反,在A549(图5A)和HeLa(图5B)细胞中过表达Smad3,划痕后0、24、48 h细胞迁移结果显示,过表达Smad3明显促进细胞迁移。以上结果表明,Smad3促进A549和HeLa细胞的迁移。

注:上图分别为A549细胞(A)和HeLa细胞(B)中敲低或过表达Smad3后进行划痕,0、24、48 h时拍照观察细胞的迁移情况并进行统计学分析,Con组作为对照组。与对照组比较,*P< 0.05,**P < 0.01,***P < 0.001。图5 敲低或过表达Smad3能够明显抑制或促进A549细胞和HeLa细胞的迁移Note. The above results show silencing of endogenous Smad3 or overexpression of Smad3 in A549 (A) or HeLa(B)cells can effectively inhibits or promotes cell migration;*P < 0.05,**P < 0.01,***P < 0.001,compared with the normal group.Fig.5 Knockdown or overexpression of Smad3 inhibits or promotes cell migration
3 讨论
癌症是导致人类死亡的主要疾病之一,其中,约90%以上的癌症患者死于癌细胞的转移。近年来,人们对肿瘤形成的分子机制展开了大量的研究,为人类治疗癌症提供了多个潜在的治疗靶点[9-11]。前期报道揭示,TGF-β与多种肿瘤的发展密切相关,包括神经胶质瘤、乳腺癌等[12-14]。在正常的生理条件下,TGF-β信号能够抑制上皮细胞的增殖,然而,在肿瘤的发展进程中,TGF-β则诱导肿瘤细胞的增殖、侵袭及血管生成[15,16]。Smad3是TGF-β信号通路中的一个重要分子,前期报道揭示,在乳腺癌晚期,Smad3异常活化,促进乳腺癌细胞的转移[7,8]。SIS3作为Smad3的抑制剂,能够抑制TGF-β1诱导的Smad3的磷酸化及Smad3-Smad4的相互作用,进而抑制相关基因的转录[17]。本研究通过过表达、敲低Smad3或利用SIS3处理A549和HeLa细胞抑制内源Smad3的活性,然后检测细胞迁移能力的变化,从而阐明Smad3对非小细胞肺癌和宫颈癌细胞迁移的影响,结果显示,Smad3促进A549和HeLa细胞的迁移。Smad3调控细胞迁移的详细作用机制有待深入研究。
综上所述,结合前期相关报道,我们发现,Smad3不仅能够促进乳腺癌细胞的迁移,也能促进非小细胞肺癌和宫颈癌细胞的迁移。这提示,在人类多种肿瘤类型中,Smad3促进肿瘤细胞的迁移具有一定的普遍性。因此,后续靶向Smad3活性抑制药物的开发将具有很大的临床应用前景,也为肿瘤治疗提供新的途径。
[1] David CJ, Huang Y-H, Chen M,etal. TGF-β Tumor Suppression through a Lethal EMT [J]. Cell, 2016, 164(5): 1015-1030.
[2] Abdollah S, Macías-Silva M, Tsukazaki T,etal. TβRI phosphorylation of Smad2 on Ser465 and Ser467 is required for Smad2-Smad4 complex formation and signaling[J]. J Biol Chem, 1997, 272(44): 27678-27685.
[3] Souchelnytskyi S, Tamaki K, Engström U,etal. Phosphorylation of Ser465 and Ser467 in the C terminus of Smad2 mediates interaction with Smad4 and is required for transforming growth factor-β signaling [J]. J Biol Chem, 1997, 272(44): 28107-28115.
[4] Shi Y, Hata A, Lo RS,etal. A structural basis for mutational inactivation of the tumour suppressor Smad4 [J]. Nature, 1997, 388(6637): 87-93.
[5] Chacko BM, Qin B, Correia JJ,etal. The L3 loop and C-terminal phosphorylation jointly define Smad protein trimerization [J]. Nat Struct Mol Biol, 2001, 8(3): 248-253.
[6] Chacko BM, Qin BY, Tiwari A,etal. Structural basis of heteromeric smad protein assembly in TGF-β signaling [J]. Mol Cell, 2004, 15(5): 813-823.
[7] Liu F, Xie W, Liu Z,etal. Role of Smad3 linker phosphorylation in breast cancer progression [J]. Cancer Res, 2015, 75(15 Supplement): 4940-4940.
[8] Luwor RB, Hakmana D, Iaria J,etal. Single live cell TGF-β signalling imaging: breast cancer cell motility and migration is driven by sub-populations of cells with dynamic TGF-β-Smad3 activity [J]. Mol Cancer, 2015, 14(1): 1.
[9] Holland EC. Gliomagenesis: genetic alterations and mouse models [J]. Nature Rev Genet, 2001, 2(2): 120-129.
[10] Bosch A, Eroles P, Zaragoza R,etal. Triple-negative breast cancer: molecular features, pathogenesis, treatment and current lines of research [J]. Cancer Treat Rev, 2010, 36(3): 206-215.
[11] Subbaramaiah K, Dannenberg AJ. Cyclooxygenase 2: a molecular target for cancer prevention and treatment [J]. Trends Pharmacol Sci, 2003, 24(2): 96-102.
[12] Pasche B. Role of transforming growth factor beta in cancer [J]. J Cell Physiol, 2001, 186(2): 153-168.
[13] Rich J. The role of transforming growth factor-beta in primary brain tumors [J]. Front Biosci, 2003, 8: 245-260.
[14] Korpal M, Yan J, Lu X,etal. Imaging transforming growth factor-β signaling dynamics and therapeutic response in breast cancer bone metastasis [J]. Nat Med, 2009, 15(8): 960-966.
[15] Derynck R, Zhang YE. Smad-dependent and Smad-independent pathways in TGF-β family signalling [J]. Nature, 2003, 425(6958): 577-584.
[16] Siegel PM, Massagué J. Cytostatic and apoptotic actions of TGF-β in homeostasis and cancer [J]. Nat Rev Cancer, 2003, 3(11): 807-820.
[17] Jinnin M, Ihn H, Tamaki K. Characterization of SIS3, a novel specific inhibitor of Smad3, and its effect on transforming growth factor-β1-induced extracellular matrix expression [J]. Mol Pharmacol, 2006, 69(2): 597-607.
Effect of Smad3 on cell migration of A549 and HeLa cells
WEI Rong-fei, LI Meng-yuan, YANG Xing-jiu, ZHU Rui-min, XU Da-mo, GAO Ran*
(Institute of Laboratory Animal Science, Chinese Academy of Medical Sciences &Comparative Medical Center, Peking Union Medical College, Beijing 100021,China)
Objective To investigate the effect of Smad3 on cell migration of A549 and HeLa cells. Methods Primers for pCMV-Myc-Smad3 plasmid construction and siRNA targeting Smad3 were designed and synthesized. pCMV-Myc-Smad3 plasmid was constructed with molecular cloning techniques. Overexpression of Smad3 with Myc-tag or silencing of endogenous Smad3, and then scratch assay was used to detect the migration ability of A549 and HeLa cellsinvitro. Results pCMV-Myc-Smad3 plasmid was successfully constructed. Overexpression of Smad3 significantly up-regulated the migration rate of A549 and HeLa cells. Conversely, in the same cells, silencing of endogenous Smad3 or treatment with Smad3 inhibitor, SIS3, down-regulated the migration rate. Conclusions Smad3 promotes cell migration of A549 and HeLa cells.
Smad3; Overexpression vector; RNAi; SIS3; Cell migration
协和青年基金资助,中央高校基本科研业务费专项资金资助(3332016077);中国医学科学院医学实验动物研究所基本科研业务费专项资助(2016ZX310033);北京市优秀人才培养项目(2015000020124G073)。
韦荣飞(1990-),女,助理研究员,研究方向:免疫与肿瘤。E-mail: weirongfei2010@163.com
高苒(1980-),女,副研究员,研究方向:肿瘤学、免疫学。E-mail: E-mail: gaoran26@hotmail.com
研究报告
R-332
A
1671-7856(2017) 01-0011-05
10.3969.j.issn.1671-7856.2017.01.003
2016-06-14
