磷脂酶A2在阿尔茨海默病中的作用机制及运动调控
2017-02-11崔莉莉李忠堂
崔莉莉,李忠堂
(牡丹江师范学院 体育科学学院,黑龙江 牡丹江 157011)
磷脂酶A2在阿尔茨海默病中的作用机制及运动调控
崔莉莉,李忠堂
(牡丹江师范学院 体育科学学院,黑龙江 牡丹江 157011)
磷脂酶A2(PLA2)是一类催化甘油磷脂sn-2酯键水解生成多不饱和脂肪酸的酶,与花生四烯酸(AA)一起在阿尔茨海默病(AD)的发病机制中起着重要作用。PLA2和AA与神经元活性有关,抑制PLA2和AA活性可抑制Aβ引起的神经毒性,缓解AD病症。运动对AD的防治已经不言而喻,运动通过抑制PLA2活性来抑制Aβ毒性,缓解AD学习记忆能力的机理仍需进一步研究。
阿尔茨海默病;磷脂酶A2;花生四烯酸;β淀粉样蛋白;运动训练
阿尔茨海默病(Alzheimer disease, AD)是一种进行性发展的致死性神经退行性疾病,临床表现为认知记忆功能不断恶化,日常生活能力进行性减退。调查发现,中国目前AD患者约为600万,且年龄每增加5岁,AD患病率将上升1倍,因此AD越来越受到研究人员的重视。AD发病机制异常复杂,其中Aβ诱导的神经毒性是其发病的最主要机制[1],近来研究发现,PLA2参与了Aβ的毒性作用,在AD的发生发展中起着极为重要的作用。
1 PLA2
PLA2存在于动物各种组织的细胞膜及线粒体膜上,是一类催化甘油磷脂sn-2酯键水解生成多不饱和脂肪酸的酶(其中包括AA)和溶血磷脂的超家族酶系。AA是一种必需脂肪酸,在神经元细胞、神经胶质细胞的信号传导中起重要作用。由于人体不能合成AA,必须从食物中摄取。AA在被运输进入大脑之前,先要在肝中合成脂蛋白,在外层附上一层磷脂质,进入大脑以后由PLA2分解出AA。AA在一系列酶的作用下可以转化成前列腺素、血栓烷、白三烯等。在正常细胞内,这些代谢产物含量很低,作为第二信使可调节信号转导、基因表达、细胞增殖等细胞功能,但当这些代谢产物大量积聚时又会产生神经毒性[2]。
PLA2的N-末端有一个Ca2+结合区域,类似于一些信号转导蛋白(如蛋白激酶C、GTP酶激活蛋白、磷脂酶C等)的Ca2+结合区域,是Ca2+作用的部位。PLA2有许多磷酸化部位,其中MAPK的Ser505是体内PLA2激活的最重要区域[3]。PLA2具有多个亚型,包括GIIA、GIVA、GVA、GVIA等,其中GIVA-PLA2存在于动物海马中,且含量相对较高,可以分解出较多的AA[4]。脑内游离的AA水平受到GIVA-PLA2活性的影响,而GIVA-PLA2的活性主要受细胞内Ca2+浓度以及磷酸化位点的调节[5]。
2 PLA2与神经元活性
研究发现,脑内的PLA2、AA在突触传递、LTP、学习记忆中起着重要的作用。对注射GVIA-PLA2抑制剂的小鼠进行Y迷宫测试时也发现其学习记忆能力下降[6]。提示,PLA2参与了脑学习记忆过程。进一步研究发现,磷脂质在PLA2的作用下产生AA、DHA等代谢物,AA通过增强神经元的兴奋性,增强海马的LTP功能,提高学习记忆能力[7],提示PLA2参与学习记忆过程可能与AA有关。对外源性PLA2注射大鼠海马切片时发现,其表面AMPA受体增多,而如果抑制PLA2的活性,表面AMPA受体水平降低[8]。提示,PLA2可通过调节AA表达,提高神经元细胞膜上AMPA受体水平,来增强神经元的活性,调节海马LTP以及学习记忆功能。但是又有学者提出疑问:PLA2与AA过多,是否会引发神经元毒性呢?大脑齿状回PLA2活性增强,AA释放增多,神经元活性增强,反过来神经元活性增强又可刺激谷氨酸盐产生Ca2+,更多的Ca2+透过NMDA受体与AMPA受体进入胞内,使得PLA2活性增强,AA水平提高。正是通过这种正反馈调节作用,PLA2活性增强使神经元活性出现了异常状态。
3 PLA2与Aβ介导的神经毒性
Aβ又称β-淀粉样蛋白,由39—43个氨基酸组成,是大脑皮质老年斑的主要成分。Aβ的可溶性二聚体可以有效削弱突触结构和功能,这种二聚体是最小的突触毒性物质,是引起AD的核心物质。在AD中,Aβ诱导IL-1β、IFN-γ、TNF-α等促炎性因子增加,过度激活PLA2,释放大量的AA,导致神经细胞的凋亡。Bate等[9]发现,大鼠皮质神经元内Aβ42增加,IFN-γ增加,PLA2活性增强,前列腺素增加,导致细胞凋亡。在AD中,Aβ引起ROS产生增多,ROS也会增强PLA2活性,增加AA释放,引起细胞凋亡。同时过量的ROS会导致氧化应激,引起神经元损伤。Kriem等[10]发现,大鼠星形胶质细胞中的H2O2刺激了PLA2的磷酸化,增加AA的释放。且Aβ还可以通过NADPH氧化酶与MAPK激活PLA2的活性,诱导线粒体损伤,导致活性氧增加,引起氧化应激损伤。Gottfried等[11]发现:AA以及ROS会抑制星形胶质细胞摄入谷氨酸盐,使得突触间隙内的谷氨酸盐水平升高,过度激活NMDA受体,增加Ca2+内流,激活PLA2的活性,同时激活对Ca2+敏感的各种酶类,产生活性氧,造成线粒体损伤,致使认知功能障碍。
在AD中,Aβ诱发的促炎性因子、ROS等物质可以激活PLA2活性,同时活性氧使得进入细胞内的Ca2+增多,激活MAPK酶,增强GIVA-PLA2活性,并将GIVA- PLA2转运到高尔基体、内质网、核膜上,使其接近磷脂质,促进AA的释放,AA通过AMPA受体与NMDA受体水平的升高增强神经元活性,过量的AA过度激活AMPA受体与NMDA受体,就会出现兴奋性中毒;且AA与其代谢产物的扩散也会使其周围区域血压升高,引起脑血管损伤;同时AA增多使脑内活性氧增加,从而引起氧化损伤[12];GIVA-PLA2的活性过强,致使过量的磷脂水解,膜上磷脂的迅速水解导致脂复合物来不及补充细胞膜的缺失,将直接导致神经元的丢失及损坏(图1)。
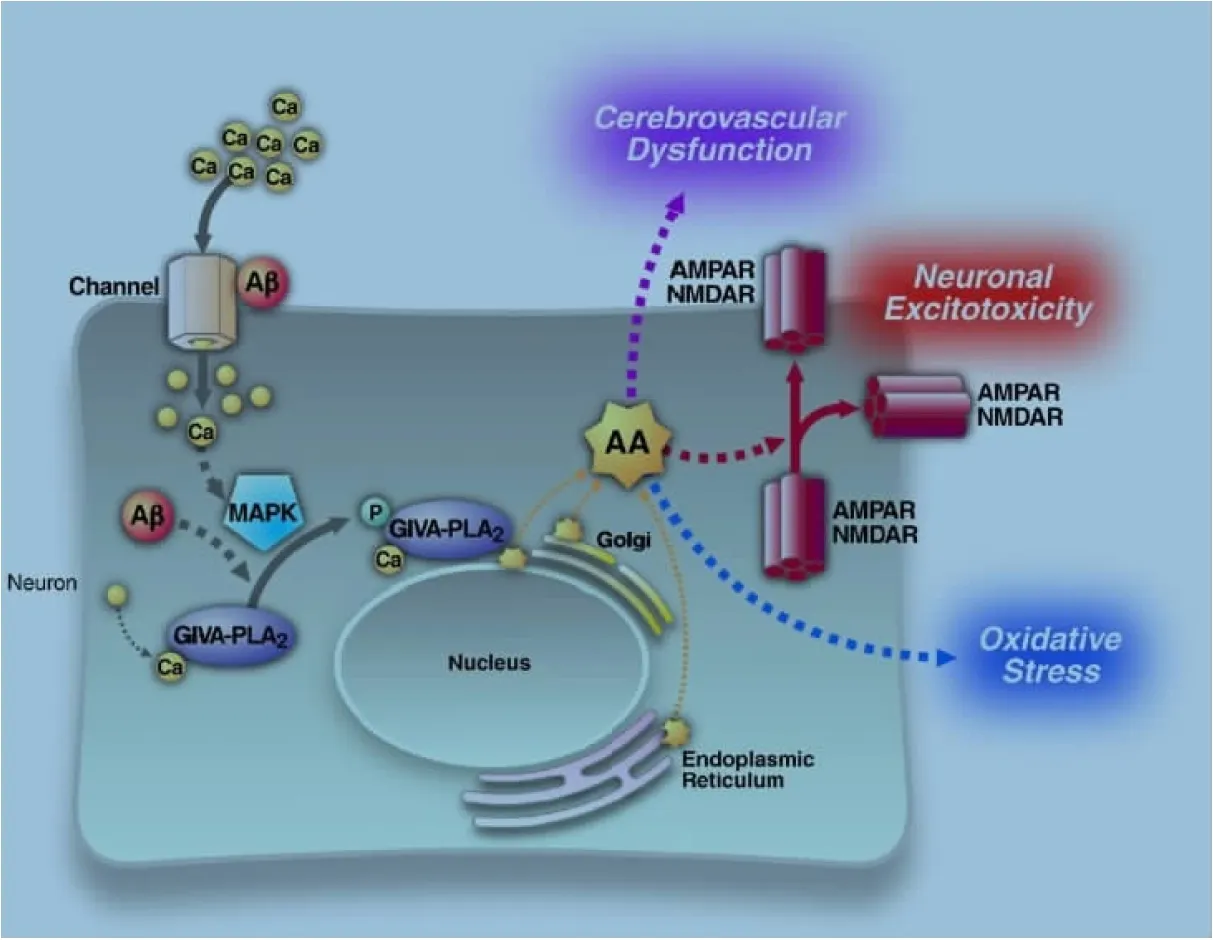
图1 PLA2在AD中的作用机制[13]
对APP转基因小鼠及同窝野生型小鼠进行液相色谱分析与质谱分析发现,AD小鼠海马中游离脂肪酸增多,脑内Aβ释放增加,GIVA-PLA2活性增强,AA含量增加,小鼠空间学习记忆能力下降,而敲除APP转基因小鼠的GIVA-PLA2基因,发现其空间学习记忆能力又有所缓解[4]。提示,Aβ可提高海马内GIVA-PLA2磷酸化水平,诱导神经毒性,而抑制PLA2活性可抑制Aβ毒性。此外,AD小鼠学习记忆能力下降也可能与钙调蛋白等突触活性蛋白下降有关。
4 运动、PLA2与AD
Aβ是导致AD的最重要致病因素,Aβ在脑内过度沉积可激活GIVA-PLA2活性,增加AA及其代谢产物释放,过度增强GIVA-PLA2活性,破坏神经元结构功能,导致神经元毒性。同时,AA可引起脑血管的损伤以及活性氧的过多释放,引起氧化损伤,导致脑内学习记忆能力下降,从而引发AD。
传统治疗AD的方法主要集中在药物疗法,该疗法毒副作用较强且无良好疗效。运动科学领域发现,积极的有氧运动可以有效延缓AD的发病。跑轮运动是一种可以规避强制性刺激带来的不利影响的自主运动方式,国外有关脑健康运动干预的研究多集中于该运动模式。Garcia等[14]对6月龄3XTg-AD转基因小鼠进行6个月自主跑轮运动干预,结果显示,长期自主跑轮运动降低了Aβ水平,抑制了AD转基因小鼠的认知功能下降。Rao等[15]研究了7个月跑轮运动对APP/PS1小鼠认知功能的影响,发现运动提高转基因小鼠认知功能与Aβ含量减少有关。提示,长期自主跑轮运动可以降低转基因AD小鼠的Aβ蛋白表达水平,抑制其认知功能的下降。研究人员也研究了短期自主运动对AD的影响,但其结果仍存在争议,如3周的自主跑轮运动可以抑制Tg2576AD小鼠脑内可溶性Aβ40表达[16],但是有研究[17]却发现,3周跑轮运动仅仅提高了该小鼠的认知功能,并没有改变Aβ40或Aβ42的表达。随后,研究人员比较了1个月和6个月自主运动对AD小鼠Aβ表达的影响,发现短期和长期运动均可以抑制Aβ40表达,且长期运动效果更好[18],提示自主跑轮运动强度较低,长期运动效果好于短期运动。针对于自主跑轮运动,跑台运动可以很好地模拟人体训练,但是该运动模式是一种强制性运动,结果易受强制性刺激的干扰。研究发现,4个月中等强度跑台运动可以降低13月龄APP转基因AD小鼠的Aβ42水平,提高认知功能[19]。Lin等[20]发现10周的跑台运动可以通过促进Aβ清除减少Aβ含量来提高4月龄APP/PS1小鼠空间学习记忆能力。以上研究表明,长期中等强度运动可以降低脑内Aβ的蛋白表达水平,提高认知功能。但是有研究将17月龄的APP/PS1转基因AD小鼠作为研究对象,对其进行5个月的跑台运动,发现运动可以减少海马内可溶性Aβ含量,增强突触可塑性,提高小鼠空间学习记忆能力,但是运动并没有改变海马内Aβ斑块的数量[21]。提示运动对AD的防治效果可能与年龄有关,推测在AD初期进行运动干预可以有效延缓AD的发病,但是随着年龄的增加,AD病情加重,运动的缓解作用减弱。以上研究结果提示,自主运动和被动运动均对AD转基因小鼠具有积极效应,且干预效果与年龄、运动的强度、时间等有关。
运动可以预防及延缓AD的发病,但是其机制并不清楚。随着研究的深入,发现PLA2在Aβ的神经毒性中起了重要作用。但是目前为止,研究人员并没有检测AD小鼠脑内的PLA2的表达水平。仅Wu等[22]研究了运动及DHA联合干预对脑外伤大鼠PLA2的影响,发现脑外伤后脑内PLA2减少,膜脂质减少,突触可塑性被抑制,认知功能下降,而运动和DHA补充均可以抑制PLA2的减少。该小鼠模型导致了PLA2表达的下降,而运动可以抑制脑外伤小鼠脑内PLA2的下降,维持膜脂质代谢。而AD模型脑内Aβ含量急剧增加,过多的Aβ增加了PLA2的活性,促进AA的释放,AA通过AMPA受体与NMDA受体水平的升高增强神经元活性,过量的AA过度激活AMPA受体与NMDA受体,引起了兴奋性中毒,因此并不清楚运动是否会抑制AD脑内PLA2的活性的增高。推测,运动可以通过减少Aβ含量将GIVA-PLA2活性、AA及其代谢产物维持在正常水平,从而提高学习记忆能力。但是,目前并没有通过AD动物模型来探讨运动对PLA2活性影响的相关研究,因此,其确切机制仍需进一步研究。
5 结论
我国已开始进入老龄化社会,AD的发病率正在逐年增加,AD越来越受到人们的重视,目前研究发现Aβ是导致AD的最主要原因,Aβ可通过过度激活PLA2来诱导神经元毒性,引起学习记忆损伤。运动作为一种积极有效的干预方式在预防及缓解AD中起了重要作用,但是运动缓解AD的分子机制不甚清楚,其确切机制仍需进一步研究。
[1] Querfurth H W, LaFerla F M.Alzheimer’s disease[J]. N Engl J Med, 2010, 362 (4) : 329-344.
[2] Farooqui A A, Horrocks L A. Phospholipase A2generated lipid mediators in the brain: the good, the bad, and the ugly[J].Neuroscientist, 2006, 12( 3) : 245-260.
[3] Sitaraman R, Israel D A, Romerogallo J, et al. Cell-associated hemolysis induced by Helicobacter pylori is mediated by phospholipases with mitogen-activated protein kinase-activating properties[J]. J Clin Microbiol, 2012, 50(3):75-78.
[4] Sanchez M O, Newman J W, Toh S, et al. Phospholipase A2reduction ameliorates cognitive deficits in a mouse model of Alzheimer’s disease[J]. Nat Neurosci,2008,11(11):1311-1318.
[5] Ahn K, McKinney M K, Cravatt B F. Enzymatic pathways that regulate endocannabinoid signaling in the nervous system[J]. Chem Rev,2008,108(5):1687-1707.
[6] Fujita S, Ikegaya Y, Nishiyama N, et al. Ca2+-independent phospholipase A2inhibitor impairs spatial memory of mice[J]. Jpn J Pharmacol,2000,83(3):277-278.
[7] Nomura T, Nishizaki T, Enomoto T, et al. A long-lasting facilitation of hippocampal neurotransmission via a phospholipase A2signaling pathway [J].Life Sci, 2001, 68(25):2885-2891.
[8] Massicotte G. Modification of glutamate receptors by phospholipase A2: its role in adaptive neural plasticity[J]. Cell Mol Life Sci, 2000, 57(11):1542-1550.
[9] Bate C, Kempster S, Last V, et al. Interferon-gamma increases neuronal death in response to amyloid-beta1-42[J]. J Neuroinflammation,2006,3(3):23-28.
[10] Kriem B, Sponne I, Fifre A, et al. Cytosolic phospholipase A2mediates neuronal apoptosis induced by soluble oligomers of the amyloid-beta peptide[J]. FASEB J,2005,19(1):85-87.
[11] Gottfried C, Tramontina F, Gonçalves D, et al. Glutamate uptake in cultured astrocytes depends on age: a study about the effect of guanosine and the sensitivity to oxidative stress induced by H2O2[J]. Mech Ageing Dev, 2002,123(10):1333-1340.
[12] Palop J J, Chin J, Mucke L. A network dysfunction perspective on neurodegenerative diseases [J]. Nature, 2006,443(7113):768-773.
[13] Sanchez-Mejia R O, Mucke L. Phospholipase A2and arachidonic acid in Alzheimer’s disease[J]. Biochim Biophys Acta, 2010,1801(8):784-790.
[14] Garcia-Mesa Y, Gimenez-Llort L, Lopez L C, et al. Melatonin plus physical exercise are highly neuroprotective in the 3xTg-AD mouse[J]. Neurobiol Aging, 2012,33(6):1113-1124.
[15] Rao S K, Ross J M, Harrison F E, et al. Differential proteomic and behavioral effects of long-term voluntary exercise in wild-type and APP-overexpressing transgenics[J]. Neurobiol Dis. 2015, 78:45-55.
[16] Nichol K E, Poon W W, Parachikova A I, et al. Exercise alters the immune profile in Tg2576 Alzheimer mice toward a response coincident with improved cognitive performance and decreased amyloid[J]. J Neuroinflammation, 2008,5(1):13.
[17] Parachikova A, Nichol K E, Cotman C W. Short-term exercise in aged Tg2576 mice alters neuroinflammation and improves cognition[J]. Neurobiol Dis, 2008,30(1):121-129.
[18] Garcia-Mesa Y, Lopez-Ramos J C, Gimenez-Llort L, et al. Physical exercise protects against Alzheimer’s disease in 3xTg-AD mice[J]. J Alzheimers Dis, 2011,24(3):421-454.
[19] Um H S, Kang E B, Leem Y H, et al. Exercise training acts as a therapeutic strategy for reduction of the pathogenic phenotypes for Alzheimer’s disease in an NSE/APPsw-transgenic model[J]. Int J Mol Med, 2008,22(4):529-539.
[20] Lin T W, Shih Y H, Chen S J, et al. Running exercise delays neurodegeneration in amygdala and hippocampus of Alzheimer’s disease (APP/PS1) transgenic mice[J]. Neurobiol Learn Mem, 2015, 118:189-197.
[21] Zhao G, Liu H L, Zhang H, et al. Treadmill exercise enhances synaptic plasticity, but does not alter β-amyloid deposition in hippocampi of aged APP/PS1 transgenic mice[J]. Neuroscience,2015, 298(9):357-366.
[22] Wu A, Ying Z, Gomez-Pinilla F.Exercise facilitates the action of dietary DHA on functional recovery after brain trauma[J]. Neuroscience, 2013, 248(38):655-663.
Functional Mechanisms of Phospholipase A2in Alzheimer Disease and Sport Exercise Regulation
CUI Li-li, LI Zhong-tang
(School of Sport Sciences, Mudanjiang Normal University, Mudanjiang 157011, China)
Phospholipase A2(PLA2) is a kind of enzyme catalyzing hydrolysis of glycerol phospholipid sn-2 ester bonds to produce polyunsaturated fatty acids. Together with arachidonic acid (AA), it plays an important role in the pathogenesis of Alzheimer disease. PLA2and AA are associated with neuronal activity. Inhibition of PLA2and AA activity could inhibit Aβ-induced neurotoxicity and alleviate AD. The function of sport exercise on AD prevention and cure has been self-evident. Sport exercise inhibits PLA2activity by inhibiting Aβ toxicity, while the mechanisms to alleviate AD learning and memory capacity still need further study.
Alzheimer disease; Phospholipase A2; Arachidonic acid; β-amyloid; sport training
2016-08-25
崔莉莉(1985-),女,山东齐河人,硕士,研究方向为运动与健康促进。
G80-05 < class="emphasis_bold">文献标志码:A
A
1008-3596(2017)01-0067-05
