红椿不同种源的苗期生长节律研究
2017-02-09阙青敏吴林瑛陈晓阳
李 培,阙青敏,吴林瑛,朱 芹,3,陈晓阳
(1 华南农业大学 林学与风景园林学院/广东省森林植物种质创新与利用重点实验室,广东 广州 510642; 2 广东省农业科学院 动物科学研究所,广东 广州 510640;3 嘉应学院,广东 梅州 514015)
红椿不同种源的苗期生长节律研究
李 培1,阙青敏1,吴林瑛2,朱 芹1,3,陈晓阳1
(1 华南农业大学 林学与风景园林学院/广东省森林植物种质创新与利用重点实验室,广东 广州 510642; 2 广东省农业科学院 动物科学研究所,广东 广州 510640;3 嘉应学院,广东 梅州 514015)
【目的】揭示红椿Toonaciliata18个种源的苗期生长规律,为培育优质红椿苗木和选择优良种源提供科学依据。【方法】对红椿18个种源一年生苗木的苗高和地径进行定期观测,利用SPSS进行方差分析,获得遗传变异系数,并利用Logistic方程对苗高和地径进行曲线拟合,估算生长参数,分析苗高、地径与地理及生态因子的相关性。【结果】红椿苗期苗高和地径遗传变异系数分别为43.31%和36.25%;参试种源苗高和地径生长的地理变异以经度控制为主。苗高和地径的生长均呈现“慢-快-慢”的“S”型生长曲线模式;在观测期间,苗高与地径生长都出现2次高峰;不同种源红椿生长性状的Logistic方程拟合决定系数为0.971~0.998,均达到了显著水平;可将红椿苗高和地径的生长分为生长前期、速生期及生长后期3个阶段。参试红椿种源在各个时期的起始及持续时间存在差异。【结论】 红椿苗高和地径遗传潜力大。不同种源生长性状的地理变异趋势明显,采种点由东到西苗高及地径生长变快。苗期生长节律存在显著差异。在速生期中,华中及华东地区种源苗高和地径快速生长持续时间较长,但生长量较小,并且年总生长量较低。
红椿; 种源; 苗期; 生长节律; 遗传变异; 地理变异
红椿Toonaciliata是楝科Meliaceae香椿属Toona植物,落叶大乔木,在我国主要分布于华南、华中、华东及西南地区,垂直分布跨度为海拔300~2 600 m[1-7],在印度、老挝、缅甸、巴基斯坦、澳大利亚也有分布[8]。红椿早期速生明显,主干通直,树姿挺秀,木材纹理优美,质地坚韧,结构细致,其材质呈现红色,素有“中国桃花心木”之称,常被作为高档家具及装饰用材出口,经济价值极高,由于生存环境的破坏,其数量不断减少,已被国家林业局列为国家Ⅱ级重点保护濒危种。目前国内外对红椿的研究主要集中在生态学、生理生化、引种和繁育等方面[9-11]。本研究选取红椿在中国全分布区内的18个种源进行育苗,定期观测苗期生长情况,分析不同种源苗高和地径生长及生长节律的差异,并利用Logistic方程对苗高和地径进行曲线拟合,为培育红椿优质壮苗和选择红椿优良种源提供依据。
1 材料与方法
1.1 试验地概况
试验地设置在华南农业大学教学科研苗圃,该地位于广州市天河区,23°09′50″N,113°21′60″E,海拔42.3 m,属南亚热带季风海洋气候,全年平均气温20~22 ℃,最冷月平均气温11 ℃,最热月平均气温28.7 ℃,极端低温-2.6 ℃,极端高温39.3 ℃。年降雨量约为1 696.5 mm,平均相对湿度68%,温暖多雨、光热充足、夏季较长。
1.2 试验材料
在我国红椿分布区中均匀布置采种点,共18个(表1)。根据果实成熟时间,分别于2013年5—11月采种。 包括西南(贵州和云南省)、华南(广西和广东省)、华中(湖北和湖南省)和华东(安徽、江西和福建省)种源。在各采种林分随机选取采种母树,要求母树无病虫害、生长正常,采种母树株距100 m以上。2013年12月进行催芽处理,方法是:种子在45 ℃温水中浸泡3~4 h,捞出后,均匀平铺于带有滤纸的育苗盘中,定期喷水,并覆盖保鲜膜,室温控制在27 ℃左右。种子咧嘴发芽后,移至混有轻基质的穴盘中生长,生根后移植到底部有透水孔的长宽为8 cm×12 cm 软塑料容器袋中,容器基质为V(黄心土)∶V(泥炭土)∶V(珍珠岩)=3∶3∶1。
表1 红椿18个种源的地理分布与生态因子
Tab.1 Locations and ecological factors of 18 provenances ofToonaciliata
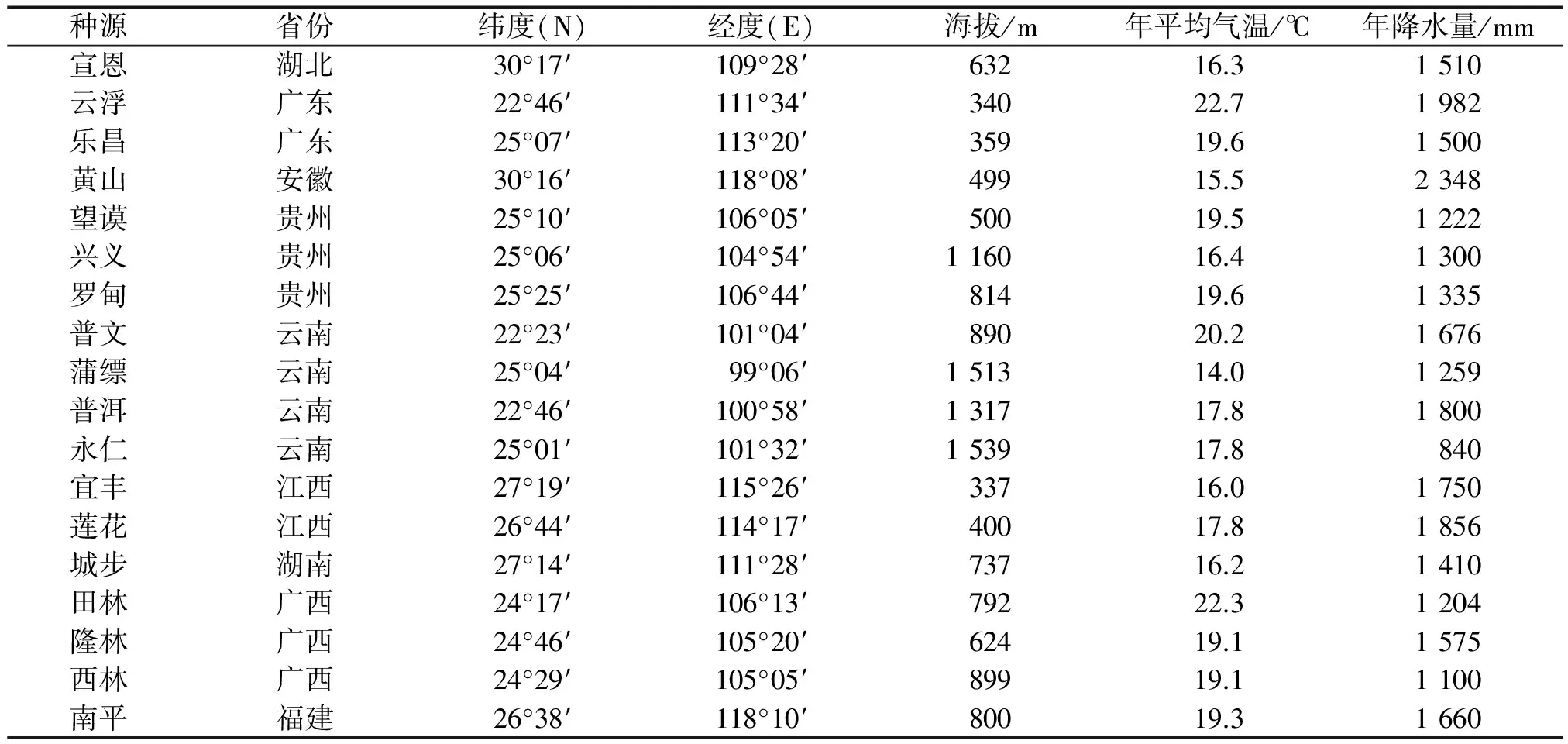
种源省份纬度(N)经度(E)海拔/m年平均气温/℃年降水量/mm宣恩湖北30°17′109°28′63216.31510云浮广东22°46′111°34′34022.71982乐昌广东25°07′113°20′35919.61500黄山安徽30°16′118°08′49915.52348望谟贵州25°10′106°05′50019.51222兴义贵州25°06′104°54′116016.41300罗甸贵州25°25′106°44′81419.61335普文云南22°23′101°04′89020.21676蒲缥云南25°04′ 99°06′151314.01259普洱云南22°46′100°58′131717.81800永仁云南25°01′101°32′153917.8840宜丰江西27°19′115°26′33716.01750莲花江西26°44′114°17′40017.81856城步湖南27°14′111°28′73716.21410田林广西24°17′106°13′79222.31204隆林广西24°46′105°20′62419.11575西林广西24°29′105°05′89919.11100南平福建26°38′118°10′80019.31660
1.3 田间试验设计
苗圃土质为酸性土红壤,地势平坦,种植穴规格为40 cm×40 cm,施鸡粪肥为基肥,每穴500 g。每个种源选取50株生长基本一致(苗高5 cm左右,地径 1 cm 左右)的植株种植于苗圃进行苗期试验。试验采取完全随机区组设计,5株小区,10个区组,株行距1 m×1 m。作为生长节律观测,每个种源每个区组固定测定3株,共30株(3株×10区组)苗木,18个种源共计540 株苗木。2014年3月23日种植,3月30日开始进行测量,每隔15 d测定苗高、地径。测量至2014年12月30日结束,共测量19次。试验期间定期除草,无追肥。2015年1月30日对18个种源所有植株(50株)苗高、地径最终测定,用以分析苗期地理变异及其规律。
1.4 数据处理
利用Excel 2007对红椿种源的苗高和地径生长量数据进行处理。利用Origin 9绘图。利用SPSS 20.0软件进行2个生长性状的方差分析、Duncan’s法多重比较(Dancan multiple range test, DMRT)及偏相关分析,分析拟合Logistic年生长曲线,探讨生长时间与苗高、地径生长量之间的关系。
按照下列公式计算2个生长性状的遗传变异系数(GCV):

(1)

Logistic曲线拟合方程[12]为:
(2)
式中,y为苗高(或地径)累积生长量,x为生长时间,k为特定条件下苗高(或地径)生长可能达到的极限值,a、b为待定系数。进行运算时,首先确定参数的初始值,达到正常的快速收敛。参数k为任意选择距离最大观测值不远的渐近线的值,之后运用最开始测定的2次观测值带入方程,求出a和b,作为初始赋值[13],得出红椿种源的苗高和地径的生长拟合曲线。
对式(2)进行多次求导,从而获得连日生长量变化速率最快的2个拐点[12]:
(3)
(4)
式中,x1、x2分别为萌动到快速生长、从快速生长转为缓慢生长的分界点,x1、x22个拐点之间即可定义为速生期。根据所得数据,结合生长量数据划分18个种源红椿苗高和地径的生长时期。
2 结果与分析
2.1 苗高与地径生长量的差异
对不同种源红椿1年生苗木的苗高与地径进行方差分析,结果(表2)表明:苗高与地径生长量在种源间及区组间的差异均达到极显著水平(P<0.01)。
表2 各种源红椿苗高与地径的方差分析
Tab.2 Analysis of variance on seedling height and ground diameter ofToonaciliatafrom different provenances
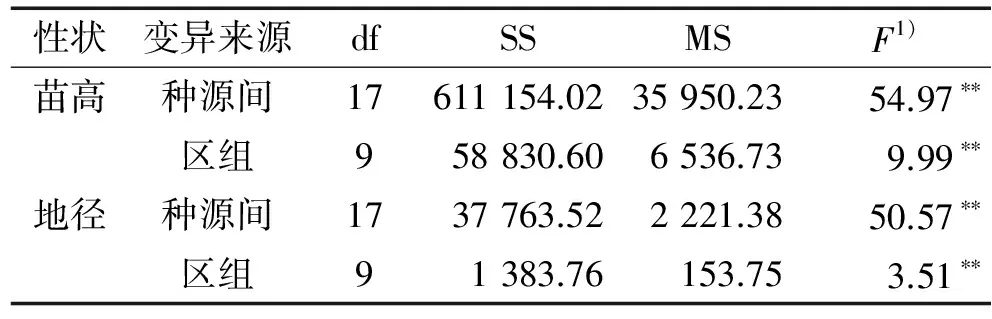
性状变异来源dfSSMSF1)苗高种源间17611154.0235950.2354.97∗∗区组958830.606536.739.99∗∗地径种源间1737763.522221.3850.57∗∗区组91383.76153.753.51∗∗
1)**表示在0.01水平上差异显著(ANOVA方差分析)。
不同种源1年生苗木的苗高和地径的生长量见表3。苗高生长表现最优的种源是云南永仁,其次是广西隆林和云南蒲缥,最差的是湖南城步;地径生长中,云南普文种源表现最优,其次是广西隆林和云南蒲缥,生长最差的为湖北宣恩种源。
表3 各种源一年生红椿苗高、地径多重比较1)
Tab.3 DMRT analysis of seedling height and round diameter of one-year-oldToonaciliatafrom different provenances

种源苗高/cm地径/mm永仁93.39±38.34a24.89±8.09a隆林93.13±32.15a26.10±7.10a蒲缥91.29±27.99a25.96±7.98a田林90.21±34.27a25.68±7.27a兴义90.11±30.14a24.71±7.77ab望谟89.31±28.44a24.96±7.34a普文88.43±34.58a26.20±7.44a普洱85.41±34.41ab24.96±7.94a西林84.96±29.57ab25.15±6.28a罗甸75.11±29.51b23.08±8.45ab云浮61.57±19.43c20.68±6.03b南平47.93±23.57d13.86±6.54c乐昌38.52±17.17de12.91±7.25c黄山28.61±9.17ef9.35±3.02de莲花28.20±10.21ef11.07±3.72cd宜丰26.31±10.39f10.93±3.36cd宣恩21.24±14.19f7.57±5.32e城步16.82±4.78f7.79±2.98e平均值67.62±38.2420.15±9.53
1)同列数据后凡是有一个相同字母者,表示种源间无显著差异(P>0.05 ,Duncan’s法)。
1年生红椿苗高和地径的遗传变异系数分别为43.31%和36.25%。
2.2 生长性状地理变异及气候生态学分析
对不同种源红椿的苗高、地径与地理因子、生态因子分别进行偏相关性分析。结果(表4)表明:种源苗木苗高与采种点经度呈极显著负相关;地径与采种点纬度呈显著负相关,与经度呈极显著负相关。海拔与2个生长性状的相关性均不显著。由此可以看出,不同种源红椿苗期生长性状的地理变异受纬度和经度双重控制,但以经度控制为主。变异趋势为采种点由东到西苗高及地径生长变快。种源苗木生长性状与采种点年降水量呈负相关,相关性达到显著水平;与年平均温度呈正相关,但相关性没有达到显著水平。
表4 红椿种源生长性状与地理及生态因子的偏相关分析1)
Tab.4 The partial correlation coefficients between growth traits and geographical, ecological factors forToonaciliatafrom different provenances

性状纬度经度海拔年均温年降水量苗高-0.411-0.671∗∗0.0190.369-0.574∗地径-0.503∗-0.657∗∗-0.0940.337-0.558∗
1)**表示在0.01水平上显著相关,“*”表示在0.05水平上显著相关(双侧检验)。
2.3 苗木生长节律的观测
从图1及图2中可以看出,各种源红椿的苗高及地径生长情况均呈现“慢-快-慢”的生长趋势,生长曲线为典型的“S”曲线。苗高生长的速生阶段主要集中在7—8月,9月下旬—10月上旬各种源的苗高生长趋于缓和,生长量并不明显;而地径生长经过4—5月的缓慢生长后,6月初开始地径生长速率明显加快,并持续到10月中下旬。从图1及图2中也可看出,西南和华南地区种源的苗高及地径生长速度在5月底以后明显比华东与华中种源快。
从不同种源的苗高净生长量曲线(图3)可以看出,各个种源每15天的净生长量差异较大,主要存在2次高峰。第1次高峰期出现在8月1日前后,之后生长速度减慢,在9月中下旬前后又出现1次生长小高峰。此种表现并不是完全遵循直线上升趋势,而是会出现1~2次生长暂缓,之后出现第2次生长高峰,通常被称为全期生长类型[14]。
对比各个种源地径的净生长量曲线(图4)可以看出,虽然不同种源的地径生长量差异较大,但整体来看,地径生长在1年中也普遍存在2个高峰期:第1个高峰期出现在8月底—9月初,达到净生长量的最大值;在之后的1个月内,生长减缓,在10月1日左右,又出现1个高峰期,虽然此次的地径生长量比第1个高峰期净生长量相对较小,但在后期的生长中也具有突出表现。2次地径生长的高峰期都晚于苗高生长的高峰期。

图1 各种源红椿苗高累积生长曲线Fig.1 Accumulated growth curves of seedling heights of Toona ciliata from different provenances
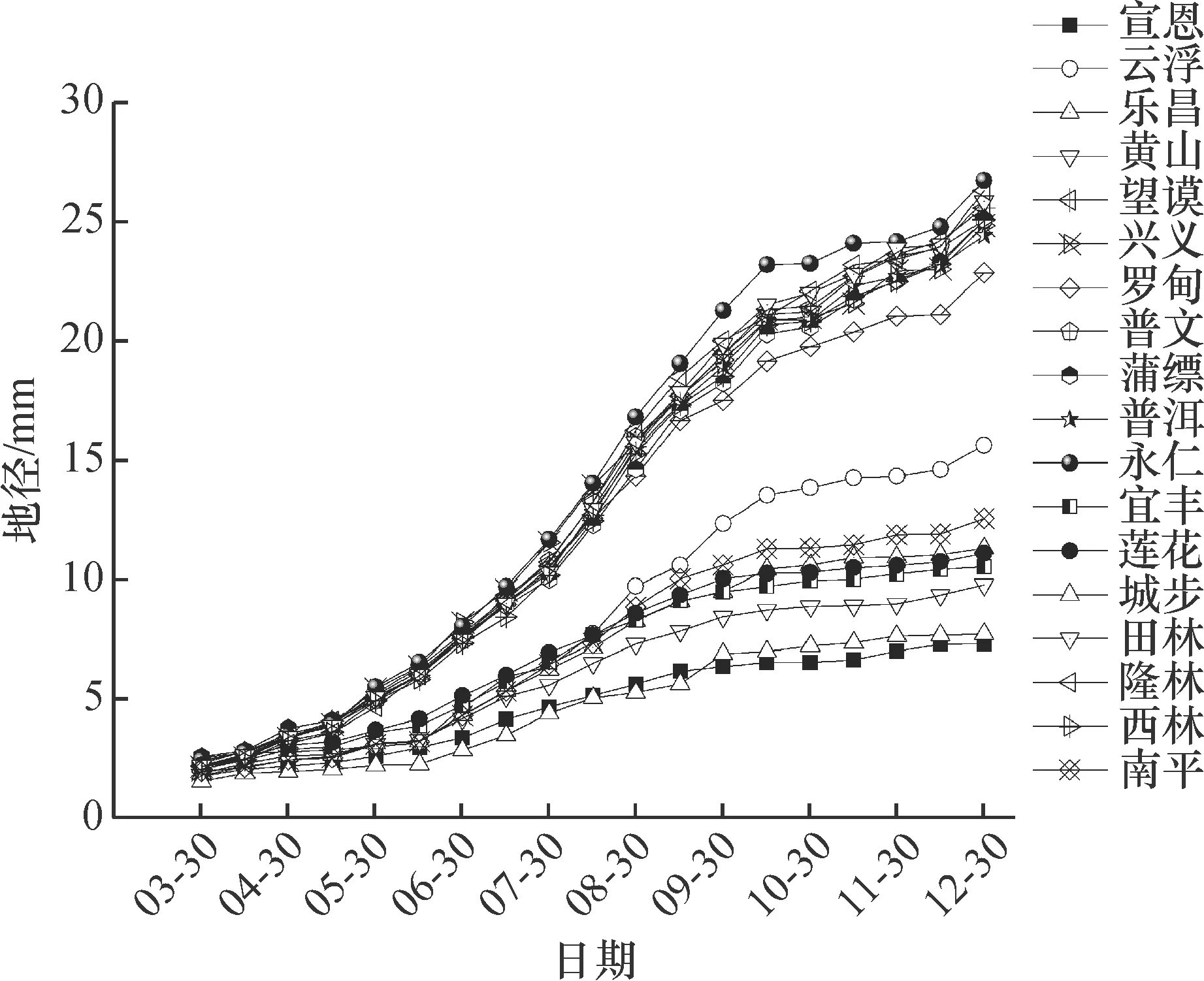
图2 各种源红椿地径累积生长曲线
Fig.2 Accumulated growth curves of ground diameters ofToonaciliatafrom different provenances
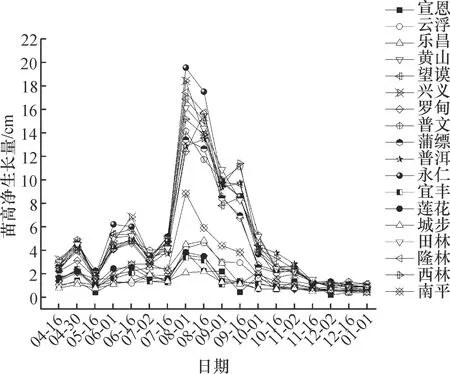
图3 各种源红椿苗高净生长曲线
Fig.3 Net growth curves of seedling heights ofToonaciliatafrom different provenances

图4 各种源红椿地径净生长曲线
Fig.4 Net growth curves of ground diameters ofToonaciliatafrom different provenances
2.4 苗木生长节律Logistic模型拟合分析
利用Logistic模型结合各种源的苗高和地径数据,对18个种源红椿生长曲线模型进行拟合(表5)。各种源苗高Logistic拟合方程的决定系数为0.971~0.992,地径的决定系数为0.982~0.998,均达到了极显著拟合水平。
采用式(3)和式(4)求出x1、x2,这2个拐点之间即可定义为速生期,之前为生长前期,之后为生长后期。各阶段起始时间、持续时间、生长量及所占总生长量的比率见表6。
18个红椿种源中,苗高生长最快进入速生期的种源为江西宜丰和莲花2个种源,生长开始15 d便进入了速生期;广东云浮、湖北宣恩、湖南城步和安徽黄山种源也较早进入快速生长期,而进入速生期最慢的是云南普洱和广西隆林种源,都经历了70 d。不同种源生长前期苗高净生长量占总生长量的比例存在差异,华中及华东地区的种源明显小于西南及华南地区种源,苗高生长量最大的是贵州兴义种源,净生长量达到24.48 cm,占该种源苗高总生长量的27.88%,是净生长量最小的湖南城步种源(0.65 cm)的37.66倍。
表5 各种源红椿苗高、地径的Logistic拟合方程
Tab.5 Logistic equations of seedling heights and ground diameters ofToonaciliatafrom different provenances

种源苗高地径Logistic方程1)R2Logistic方程1)R2宣恩y=32.124/(1+5.713e-0.016x)0.992y=7.773/(1+5.220e-0.016x)0.989云浮y=36.799/(1+4.677e-0.014x)0.971y=17.071/(1+14.643e-0.018x)0.984乐昌y=42.149/(1+6.177e-0.018x)0.989y=12.161/(1+9.885e-0.019x)0.990黄山y=30.734/(1+5.603e-0.021x)0.994y=10.125/(1+7.431e-0.018x)0.991望谟y=95.537/(1+19.336e-0.026x)0.986y=27.070/(1+17.137e-0.020x)0.997兴义y=100.421(1+21.028e-0.026x)0.987y=25.311/(1+15.343e-0.020x)0.998罗甸y=83.135/(1+19.124e-0.026x)0.982y=23.783/(1+13.601e-0.019x)0.997普文y=97.703/(1+16.727e-0.023x)0.985y=26.529/(1+16.252e-0.020x)0.998蒲缥y=88.839/(1+14.264e-0.024x)0.988y=26.974/(1+14.735e-0.018x)0.996普洱y=95.321/(1+19.859e-0.024x)0.987y=25.666/(1+17.094e-0.020x)0.997永仁y=102.881/(1+25.681e-0.028x)0.985y=27.407/(1+18.630e-0.021x)0.995宜丰y=28.011/(1+3.910e-0.017x)0.992y=11.114/(1+7.072e-0.019x)0.990莲花y=30.812/(1+4.310e-0.021x)0.986y=11.631/(1+6.774e-0.018x)0.991城步y=21.794/(1+5.626e-0.017x)0.988y=8.683/(1+8.356e-0.016x)0.982田林y=98.363/(1+20.618e-0.026x)0.985y=27.058/(1+18.47e-0.020x)0.996隆林y=96.754/(1+26.202e-0.028x)0.988y=26.642/(1+19.676e-0.021x)0.996西林y=93.426/(1+22.004e-0.026x)0.983y=25.993/(1+17.976e-0.020x)0.996南平y=48.987/(1+13.441e-0.023x)0.985y=13.086/(1+12.800e-0.020x)0.982
1)y代表苗高或地径累积生长量,x代表生长时间。
华中及华东地区种源在4月中下旬就已进入速生期,比华南及西南地区种源早近1个月,并且华中及华东地区种源在速生期持续的时间比华南及西南地区种源长50 d 左右,但苗高生长量较小,因而总生长量较低。在速生期内,广西隆林种源苗高净生长量达到62.81 cm,生长量最大,持续的时间最短。湖南城步种源在速生期中,苗高生长量最小,只有13.17 cm,占总生长量的73.71%。从9月初,各种源的速生期结束,苗高生长量逐渐减小,维持小幅度增长。广东云浮速生期持续到10月中旬,结束的时间最晚。
表6 各种源红椿苗高(地径)生长阶段划分及生长参数1)
Tab.6 Growth stage classifications and growth parameters for seedling heights(ground diameters)ofToonaciliatafrom different provenances

种源生长前期速生期生长后期持续时间/d净生长量/cm(mm)占总生长量比例/%起始日期(月-日)持续时间/d净生长量/cm(mm)占总生长量比例/%起始日期(月-日)持续时间/d净生长量/cm(mm)占总生长量比例/%宣恩27(31)2.40(0.29)10.28(5.10) 04-19(04-22) 165(165)16.44(4.33)70.52(77.51)09-30(10-03) 85(81)4.48(0.97)19.20(17.39)云浮16(76)1.06(0.63)4.31(4.78)04-08(06-05) 188(146) 20.23(10.75)82.26(81.46)10-12(10-28) 73(55)3.30(1.82)13.43(13.76)乐昌28(51)3.95(0.81)12.30(8.52) 04-20(05-11) 146(139)22.76(7.82)70.88(82.32)09-12(09-27)103(87)5.40(0.87)16.82(9.16)黄山19(38)1.67(0.66)6.72(8.24)04-10(04-30)125(146)18.18(6.05)73.24(75.03)08-13(09-24)133(93)4.98(1.35)20.04(16.73)望谟63(76)22.87(3.83)27.68(16.30)05-23(06-05)101(132)50.47(16.80)61.07(71.48)08-21(10-14)113(69)9.30(2.87)11.25(12.22)兴义66(71)24.48(4.23)27.88(18.55)05-26(05-31)101(132)53.45(14.7)60.88(64.44)08-24(10-08)110(74)9.87(3.88)11.24(17.01)罗甸63(68)18.17(2.86)25.81(13.85)05-23(05-28)101(139)44.60(15.31)63.34(74.06)08-21(10-13)113(70)7.64(2.50)10.85(12.09)普文65(74)22.26(3.97)27.17(17.39)05-25(06-03)115(132)54.46(16.42)66.48(71.95)09-16(10-12) 97(71)5.20(2.43)6.35(10.66)蒲缥56(76)18.68(3.53)24.95(15.59)05-16(06-05)110(146)48.03(16.54)64.13(73.07)09-13(10-28)111(55)8.18(2.57)10.92(11.34)普洱70(76)20.85(3.81)25.56(17.07)05-30(06-05)110(132)52.93(16.36)64.86(73.29)09-17(10-14) 97(69)7.82(2.15)9.58(9.64)永仁69(77)24.07(4.14)26.81(16.93)05-29(06-06) 94(125)58.51(16.80)65.17(68.78)08-30(10-07)114(75)7.20(3.49)8.02(14.29)宜丰15(34)1.37(0.74)6.60(8.72)04-07(04-26)155(139)15.36(6.27)74.12(74.43)09-08(09-11) 107(104)3.99(1.42)19.27(16.85)莲花15(33)1.68(0.84)7.30(9.44)04-07(04-25)155(146)14.84(6.34)64.61(71.01)09-09(09-17)107(98)6.45(1.75)28.09(19.56)城步24(50)0.65(0.50)3.65(8.06)04-16(05-10)155(165)13.17(5.35)73.71(85.99)09-17(10-21) 98(62)4.05(0.37)22.64(5.95)田林66(80)14.32(3.84)16.87(16.16)05-26(06-09)101(132)61.01(16.88)71.91(70.97)09-03(10-20)110(65)9.51(3.06)11.21(12.88)隆林70(79)13.19(3.86)15.18(15.89)05-30(06-08) 94(125)62.81(17.29)72.28(71.18)09-01(10-11)113(73)10.90(3.14)12.54(12.93)西林68(79)12.56(3.79)15.64(16.38)05-28(06-08)101(132)59.37(16.16)73.94(69.75)09-05(10-19)108(66)8.37(3.22)10.42(13.88)南平56(62)4.49(1.15)11.05(10.76)05-16(05-22)115(132)31.13(8.28)76.66(77.54)09-07(09-30)106(63)4.99(1.25)12.30(11.70)
1)表中数据括号外为苗高,括号内为地径。
在生长后期,各种源的苗高生长量占总生长量的比例存在较大差异。在广州的生长环境条件下,华中及华东地区种源生长后期苗高生长量所占总生长量的比例较生长前期占比大,而华南及西南地区种源生长前期生长量所占比例大于生长后期,这表明华中及华东地区的种源生长持续性较好。
在地径的生长过程中,各个种源进入速生期的时间普遍延后于苗高的生长时间。在生长前期地径净生长量最大的是贵州兴义种源(4.23 mm),最小的是湖北宣恩种源(仅有0.29 mm),这可能与该种源最早进入速生期有关。5—6月,地径逐渐进入速生期,平均持续时间为139 d。维持时间最长的是湖南城步和湖北宣恩种源,可达到165 d;持续最短的是广西隆林和云南永仁种源,只有125 d。地径生长量最大的是广西隆林种源,在125 d的速生期内加粗17.29 mm,占总生长量的71.18%。地径生长量最小的是湖北宣恩种源,仅有4.33 mm。进入9月末,各种源陆续结束地径速生生长,进入到生长后期。最晚结束地径速生期的是广东云浮和云南蒲缥种源,在10月末结束快速生长。在生长后期,18个种源红椿的地径生长量明显减弱,华中及华东地区种源地径生长量也明显小于华南及西南地区种源。
3 讨论与结论
红椿生长快,材质好,属濒危树种,近年来受到国内外广泛关注。本研究结果表明,不同种源间在苗高与地径生长量上的差异均达到显著水平。不同种源红椿苗期生长性状的地理变异受纬度和经度双重控制,且以经度控制为主。变异趋势为采种点由东到西苗高及地径生长变快。种源苗木生长性状与采种点年降水量呈负相关,相关性达到显著水平,与年平均温度呈正相关,但相关性没有达到显著水平。
遗传变异系数是衡量相关生长性状遗传变异潜力的有效指标,若某一性状该数值越大,则根据该性状选择出优良遗传型的潜力越大[15-16]。红椿苗期苗高和地径的遗传变异系数分别为43.31%和36.25%,其值较高,说明红椿1年生苗木苗高和地径具有较大选择潜力,可作为早期选择依据之一。
“S”型生长曲线可以用于描述某一种群受空间约束的生长过程,其中表现出的各种参数在林木培育过程中具有重要参考价值[17]。各个红椿种源苗高和地径生长在一年中都呈现“慢-快-慢”的生长规律,并符合“S”型生长模型。利用Logistic模型拟合红椿各种源的苗高、地径数据,各种源苗高Logistic拟合方程的决定系数为0.971~0.992,地径的决定系数为0.982~0.998,均达到了极显著拟合水平,表明用Logistic方程拟合红椿苗高、地径生长节律精确度较高。
红椿苗高在7—9月生长最快,与其变种毛红椿T.ciliatavar.pubescens生长高峰期类似[18],地径生长的高峰期出现在8—10月。10月后,苗高和地径的生长缓慢。苗高和地径在生长过程中都存在2个不连续的高峰期,与同属植物香椿T.sinensis及金华香椿T.jinhuaensis生长节律相似[19],并与苦楝MeliaAzedarach、红榉Zelkovaschneideriana及美丽异木棉Ceibaspeciosa呈现相同趋势[14, 20-21]。
根据Logistic方程求导所得的拐点进行苗期生长时期的划分,苗高和地径都可以划分为生长前期、速生期和生长后期3个阶段。不同种源各个时期的划分起始时间与持续时间存在明显差异。总体而言,华中和华东种源速生期早于西南和华南种源,持续的时间较长。原因可能是华中和华东地区的气温比西南和华南低,则华中和华东种源生长所需温度比西南和华南种源低,当华中和华东种源引种到广东,自然比西南和华南种源萌芽早,停止生长晚。18个红椿种源在苗高与地径生长上存在极显著差异,说明种源选择对红椿遗传改良是很有意义的。以速生性改良为目标时,速生期的生长量可以作为速生选择的基本参考标准[12]。广西隆林种源虽然速生期持续时间最短,但在速生期苗高及地径净生长量最大,因而该种源的生长表现最优。此外,云南永仁和广西田林种源在速生期生长量也较大。在广东开展红椿造林可以重点考虑这3个种源,但是否为优良种源,仍需进行造林试验才能最终确定。
揭示和总结红椿种苗不同时期的生长特性,对红椿种源的苗期育苗具有重要的指导意义。速生期是树木生长的关键时期,应当通过施肥等抚育管理措施,促进苗木的快速生长,生长后期应注意提高苗木木质化的程度,以提高其抵御冬季寒害的能力。
[1] 张亚东,钟艺,周国清,等. 湖北恩施种源红椿不同家系育苗试验初报[J]. 湖北林业科技, 2013,42(3): 17-20.
[2] 陈昌笃,李迪华. 湖南省武陵源地区的生物多样性和生态完整性[J]. 生态学报, 2003, 23(11): 2414-2423.
[3] 龙汉利,冯毅,向青,等. 四川盆周山地红椿生长特性研究[J]. 四川林业科技, 2011, 32(3): 37-41.
[4] 冯立新,陈荣,朱昌叁,等. 桂西北红椿种群年龄结构与空间分布格局研究[J]. 西北林学院学报, 2015, 30(1): 46 50.
[5] 王达明,杨绍增,张懋嵩,等. 云南珍贵用材树种的产材类别品性及分布特征[J]. 西部林业科学, 2012, 41(1): 7-16.
[6] 邹军. 红椿发芽试验与育苗技术[J]. 云南农业科技, 2012(6): 47-48.
[7] 薛克娜,何育桥,李镇魁,等. 广东省珍稀濒危植物现状调查[J]. 中国城市林业, 2010, 8(3): 53-56, 33.
[8] BOLAND D J.ToonaciliataM. Roemer[M]. Wallingford: CAB International, 2000: 6472-6485.
[9] MALAIRAJAN P, GOPALAKRISHNAN G, NARASIMHAN S, et al. Anti-ulcer activity of crude alcoholic extract ofToonaciliataRoemer (heart wood)[J]. J Ethnopharmacol, 2007, 110(2): 348-351.
[10]冯立新,陈荣,朱昌叁,等. 红椿天然林优树选择[J]. 福建林业科技, 2015, 42(1): 111-114.
[11]吴际友,程勇,王旭军,等. 红椿无性系嫩枝扦插繁殖试验[J]. 湖南林业科技, 2011, 38(4): 5-7.
[12]邝雷,邓小梅,陈思,等. 4个任豆种源苗期生长节律的研究[J]. 华南农业大学学报, 2014, 35(5): 98-101.
[13]董江水. 应用SPSS软件拟合Logistic曲线研究[J]. 金陵科技学院学报, 2007, 23(1): 21-24.
[14]廖美兰,王华新,汪艳云,等. 美丽异木棉苗期生长节律[J]. 广西林业科学, 2014, 43(1): 101-104.
[15]王明庥,黄敏仁,吕士行,等. 黑杨派新无性系研究[J]. 南京林业大学学报,1987,11(2): 1-12.
[16]宋德凤,吴高岭. 玉米种质资源遗传力和遗传变异系数的研究[J]. 湖北农学院学报, 1999, 19(3): 212-214.
[17]杨志玲,杨旭,谭梓峰,等. 厚朴不同种源苗期生长模型的拟合[J]. 西北农林科技大学学报(自然科学版), 2011, 39(4): 60-68.
[18]周诚,廖海红,王丽艳,等. 毛红椿播种苗苗期生长规律分析[J]. 林业科技开发, 2013, 27(1): 18-21.
[19]周翔宇. 中国香椿属的研究[D]. 南京: 南京林业大学, 2005.
[20]易文成,王旭军,张连金,等. 大叶榉不同种源早期生长比较[J]. 湖南林业科技, 2014, 41(5): 22-24.
[21]陈丽君,刘明骞,廖柏勇,等. 苦楝不同种源苗期生长性状和生长节律研究[J]. 西南林业大学学报, 2014, 34(4): 1-7.
【责任编辑 李晓卉】
Growth rhythms of Toona ciliata seedlings from different provenances
LI Pei1, QUE Qingmin1, WU Linying2, ZHU Qin1,3, CHEN Xiaoyang1
(1 College of Forestry and Landscape Architecture, South China Agricultural University/Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, Guangzhou 510642, China; 2 Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China; 3 Jiaying University, Meizhou 514015, China)
【Objective】 To study the growth rhythms ofToonaciliataseedlings from 18 provenances, to provide a scientific basis for breeding and screening high-qualityT. ciliataprovenance. 【Method】Heights and ground diameters of one-year-oldT. ciliataseedlings from 18 provenances were regularly observed. Analysis of variance was performed using SPSS, and the genetic coefficients of variation (GCV) were obtained. The Logistic equations were used for fitting the growth curves of seedling heights and ground diameters, and for estimation of the growth parameters. The correlations between seeding height, ground diameter and geographical, ecological factors were investigated. 【Result】The GCV of seeding height and ground diameter ofT. ciliatawere 43.31% and 36.25% respectively. The geographic variations in two traits were mainly influence by longitude. The growths of the two traits were both in a “slow-fast-slow” pattern showing a similar “S” curve. Two growth peaks of each trait were observed. The determination coefficients of Logistic equations varied from 0.971 to 0.998 forT.ciliatafrom different provenances, and the fitting degrees reached significant levels. The growth ofT.ciliataseedlings could be divided into three stages, including the early, fast and late growth stages.T.ciliatafrom different provenances had different starting and ending time for the three stages.【Conclusion】There are great heredity potentials in seeding height and ground diameter ofT.ciliata. Geographic variation trends of the two traits are obvious among different provenances. The seeding height and ground diameter grew faster forT.ciliatafrom western provenances compared to eastern provenances. There were significant differences in seedling growth rhythms. During the fast growth stage, the growths in seedling height and ground diameter forT.ciliatafrom eastern provenances lasted longer compared to western provenances, while the extent of growth of this stage and yearly growth were both smaller for the provenances from central China and east China.
Toonaciliata; provenance; seedling; growth rhythm; genetic variation; geographic variation
2016- 03- 18优先出版时间:2016-12-28
李 培(1985—),女,博士,E-mail:lipei-meinv@163.com;通信作者:陈晓阳(1958—),男,教授,博士,E-mail:xychen@scau.edu.cn
“十三五”国家重点研发计划林业资源培育及高效利用技术创新重点专项(2016YFD0600606); 国家林业局林业公益性行业科研专项(201004020)
S722.3
A
1001- 411X(2017)01- 0096- 07
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20161228.0922.010.html
李 培,阙青敏,吴林瑛,等.红椿不同种源的苗期生长节律研究[J].华南农业大学学报,2017,38(1):96- 102.
