中国弄蝶科昆虫分布格局的简约性分析
2017-02-09黄启通李劲松庄海玲黄珍富范骁凌
黄启通,李劲松,庄海玲,黄珍富,汤 静,王 敏,范骁凌
(华南农业大学 农学院,广东 广州 510642)
中国弄蝶科昆虫分布格局的简约性分析
黄启通,李劲松,庄海玲,黄珍富,汤 静,王 敏,范骁凌
(华南农业大学 农学院,广东 广州 510642)
【目的】根据中国弄蝶的分布特点划分生物地理区并推断弄蝶现代分布格局的成因。【方法】统计得到中国328种弄蝶及其分布;将中国地图划分成2°×2°的经纬网格,总计294个,依据地形、植被和弄蝶分布的特点将294个网格进一步汇总为28个大小和形状不一的区域,采用特有性简约分析方法进行分析。【结果】中国弄蝶的分布包括10个主要区,即中部地区(B1)、南部地区(B2)、台湾地区(B3)、中北部地区(C2)、西部地区(C3)、北部地区(D2)、秦巴山区(D3)、东北地区(D4)、西南地区(E1)和西藏地区(E2),其中台湾和西藏分别自成一支。通过比对地质板块与弄蝶的分布格局,表明弄蝶主要分布在秦巴山区、藏东南地区、喜马拉雅山东段南坡、黄河和长江中下游地区以及南部地区。中国弄蝶的分布区与中国动物地理区划的对比结果显示,两者有5个区基本相符,分别为中部地区(B1)、南部地区(B2)、西部地区(C3)、北部地区(D2)和东北地区(D4)。【结论】中国弄蝶的分布区和中国地质省区的划分基本吻合。
弄蝶; 生物地理区; 中国; 分布格局; 地质省区; 特有性简约分析
历史生物地理学在揭示地质历史变迁、解释生物多样性的演化历程方面起着重要的作用。生物演化以地质历史的变迁为客观条件,而地质历史的变迁又以生物分布规律及其演化进程为其佐证。随着板块的分裂、移动或连接,新的山脉随之隆起、海洋出现或消失、气候转变,生物物种分化、灭绝和扩散[1]。我国地处亚洲大陆东南边缘,晚古生代以来泛大陆(Pangaea)的解体和重组,特别是印度板块的北移与青藏高原的急剧隆起,对我国现生生物的分布格局产生重大影响。Hsü等[2]基于大陆内部的地缝合线将中国地壳划分为华北板块、塔里木板块、扬子板块、华夏板块、西伯利亚板块和冈瓦纳板块的一部分。受地壳运动的影响,至更新世,这六大板块与古西伯利亚和欧洲板块连为一体[2-3]。受地壳运动的影响,中国的地形地势西高东低,以青藏高原最高,向东逐渐下降呈阶梯状分布。
中国复杂的地质演化历史(喜马拉雅造山运动和青藏高原隆起)、错综紊乱的地形(青藏高原,长江、黄河中下游平原等)及其历史演替过程和气候类型(从最北部的寒带至最南部的热带)的多样性不仅对现存生物的地理分布格局产生重大影响,而且也使得中国生物地理学的研究结果备受国际学术界的重视。张荣祖[4]根据对现代陆栖脊椎动物的研究,将我国大陆动物区系划分为古北界和东洋界;古北界包括西北、东北和华北的大部分地区;东洋界包括以西南、华中区南部以及华南区为主的中国南部的省区。中国作为东亚地区的主体,是唯一横跨两大动物界的国家,其各生物类群的研究对于生物地理学的研究具有重要意义。近年来,依据两栖类、鸟类、陆生哺乳类和高等植物的分布进行生物地理学研究的报道越来越多[5-7]。然而,基于昆虫类群进行分布格局的研究并不多见。
蝴蝶是自然生态系统的重要组成部分之一,与被子植物在长期的协同进化过程中形成了密切的关系。蝴蝶数量是衡量一个地区生物多样性丰富程度的重要指标。本文选用地理分布数据较为全面的弄蝶科Hesperiidae作为研究类群,采用特有性简约分析方法对其分布进行聚类分析,在此基础上对中国弄蝶的分布进行分区,推测弄蝶现有分布的成因,为中国的地质历史演化提供生物学方面的证据。
1 材料与方法
1.1 研究区域
本文基于Meng等[8]的方法,依据中国的地形、植被、弄蝶科昆虫的分布特点和已有的其他研究结果[5],对弄蝶的28个地理区域进行了稍微的调整,譬如:将四川分为川西高原(A15)和四川盆地(A13),将云南分为滇南山地(A08)和云南高原(A27),采纳冀晋山地(A07)和秦巴山区(A06)单独分区的观点,结果见图1。
1.2 方法
1.2.1 构建分布矩阵 查阅《中国动物志》及其他相关文献资料,结合华南农业大学鳞翅目昆虫研究室多年来的野外调查实物标本,统计中国已记录的弄蝶种类及分布,得到共计 428 种(包括亚种)的分布记录。将分布区有某物种存在编码为“1”,反之,编码为“0”,将不符合特有性简约分析原理的物种(分布区过于狭窄的物种)排除在外,最终用于分析的弄蝶为 328种。以划定的 28 个区为内群,假设 1 个没有任何分布的 A0 为外群[9-11],构建 1 个29×328 的矩阵。
1.2.2 特有性简约分析 特有性简约分析(Parsimony analysis of endemicity, PAE)是生物地理学研究中普遍采用的方法,用于生物地理格局及其特有性分析。该方法对划定区域的大小和形状没有特殊的要求,且区域的大小和外形不会影响其分析结果[12]。本研究根据28个区域(图1)中的物种数,采用特有性简约分析方法对其分布格局进行分析。利用PAUP4.0[13]构建简约树(Maximum parsimony, MP),进行自展检验[14],并以重复 1 000 次抽样的自展检验值(Bootstrap proportion, BP)来评估其分支的稳定性。

A01:淮北平原;A02:洞庭湖平原;A03:江汉平原;A04:浙闽丘陵北段;A05:西部荒漠;A06:秦巴山区;A07:河北平原;A08:滇南丘陵;A09:鄱阳湖平原;A10:内蒙古高原;A11:青藏高原东北部;A12:山西高原;A13:四川盆地;A14:长白山区;A15:川西高原;A16:贵州高原;A17:冀北辽西山地;A18:东北北部平原;A19:西北荒漠、高原;A20:山东丘陵;A21:西藏高原;A22:浙闽丘陵南段;A23:台湾地区;A24:广东沿海丘陵;A25:广西盆地;A26:海南地区;A27:西南山地;A28:陕晋高原。
图1 基于中国地形特点划分的28个弄蝶地理区
Fig.1 The 28 geographic areas of skippers in China based on topographical characteristics
2 结果与分析
构建基于弄蝶分布特点的MP树(图2),步长=1 135,一致性指数(Consistency index,CI)=0.290 7,存留指数(Retention index,RI)=0.448 3。中国弄蝶分布区聚成12支:B1[(A01+A03+A04+A09)+(A02+A13)],B2[(A08+A26+A22+A24+A25)+(A16)],B3(A23),C1(A11),C2(A05+A28),C3(A10+A19),D1(A20),D2(A07+A12),D3(A06),D4[(A14+A18)+(A17)],E1(A15+A27)和E2(A21);而且每一支对应一种地形。其中B1包括 2 个亚支:“A01+A03+A04+A09”和“A02+A13”,并得到一定的支持(BP=34),它们分别与中国动物地理区划中的东部丘陵平原亚区和西部山地高原亚区相对应。B2由“A08+A26+A22+A24+A25”和“A16”2 个亚支组成,分别对应于滇南山地亚区、海南岛亚区、闽广沿海亚区和贵州高原南部地区,且得到一定的支持(BP=39)。D2由A07和A12聚成,以冀晋山区为主。D4对应于东北三省,包括长白山山区和小兴安岭山区,且其支持值(BP=50)高于D2的(BP=47)。C2包括A05和A28,以黄土高原为主。C3由A10和A19组成,包括中国西北荒漠草原。E1分为A15和A27亚支,依次对应于川西高原和云南高原,且支持值(BP=43)低于C2(BP=52)和C3(BP=55)。B3(A23)、C1(A11)、D1(A20)、D3(A06)和E2(A21)分别独立成支,依次对应于台湾地区、青海地区、黄淮海平原地区、秦巴山区和西藏高原地区。
根据简约性分析结果和弄蝶物种的分布特点,将中国版图划分成10个主要区(图3):中部地区(B1);南部地区(B2);台湾地区(B3);中北部地区(C2);西部地区(C3)包括内蒙古和新疆;北部地区(D2);秦巴山区(D3);东北地区(D4);西南地区(E1);西藏地区(E2)。
中部地区(B1)包括四川盆地、长江及其支流。气候属于亚热带暖湿类型,雨热同期。总体地貌特征属长江水系,河网密布,山地、丘陵、平原交错,地形复杂,弄蝶生物多样性较丰富(184种),其中有1个特有种。南部地区(B2)包括广东、广西和云南南部、福建东南沿海、海南、贵州,总的地势由西北向南、东南或西南倾斜,境内山地、丘陵、平原、河川纵横交错,地形复杂,气候类型众多,属南亚热带及热带,降水充沛,植被茂密,弄蝶生物多样性最为丰富,由于复杂的地质历史演化,该区特有种十分丰富,共计38种。台湾地区(B3)因欧亚大陆板块、菲律宾海洋板块挤压而隆起,全岛地形东高西低,具有丰富的低地、低山及高山环境。高山环境为动物提供了合适的居所,有利于接纳不同水平地带的种类,故该区弄蝶生物物种相对丰富,共计61种。中北部地区(C2)由黄土高原及河谷地带组成,受人为因素的影响,原有的天然森林和植被自然景观已被农田和零星分散的林地所替代,剩余的一点天然植被仅在黄土塬上的山地有分布,生物赖以生存的栖息环境的消失使得该区弄蝶种类少(54种)。西部地区(C3)涵盖新疆和内蒙古,主要以草原和荒漠为主,气候属于半干燥和干燥型,东西部差异显著,西部受西来湿润气候的影响,形成草原。干旱的气候、荒漠和草原为主的植被条件使得弄蝶资源颇为贫乏(24种),其中有1个特有种。北部地区(D2)包括北京市、天津市、山西省、河北省及陕西东部的一部分,属于暖温带半湿润大陆性气候,四季分明,光照充足;冬季寒冷干燥且较长,夏季高温降水相对较多,春秋季较短,受人类活动的干扰较大,弄蝶种类和数量较少(42种)。秦巴山区(D3):秦岭、巴山是我国中西部的横断山系,自西向东,横亘甘肃南部、陕西南部、河南西部的广大地区,是世界动物地理区划东洋界与古北界的交汇处,属于亚热带和温带过渡地带,森林茂盛,弄蝶种类较多(97种)。东北地区(D4)由大、小兴安岭,张广才岭,老爷岭,长白山以及松花江和辽河平原组成,属于温带湿润、半湿润大陆性季风气候。该区弄蝶少(34种),无特有种。西南地区(E1)包括四川西部、云南中北部,境内三江并流,南北走向的高山、峡谷纵横交错,气候复杂多变,干湿季明显。西南山地复杂的地形与有利的湿润条件使其生物多样性非常丰富,拥有大量的特有动植物种类,该区弄蝶种类十分丰富(180种),占最终用于分析弄蝶种类的55%,含8个特有种。西藏地区(E2)位于我国西南部,平均海拔 4 000 m以上,属典型的大陆性气候,境内地理位置高低悬殊,生态环境极其复杂多样,由于地势高,各地平均气温都较低,且随着地形地势的变化气温差异较大,自东南至西北,从温、热带逐渐过渡到寒冷地区,本区记录了65种弄蝶。
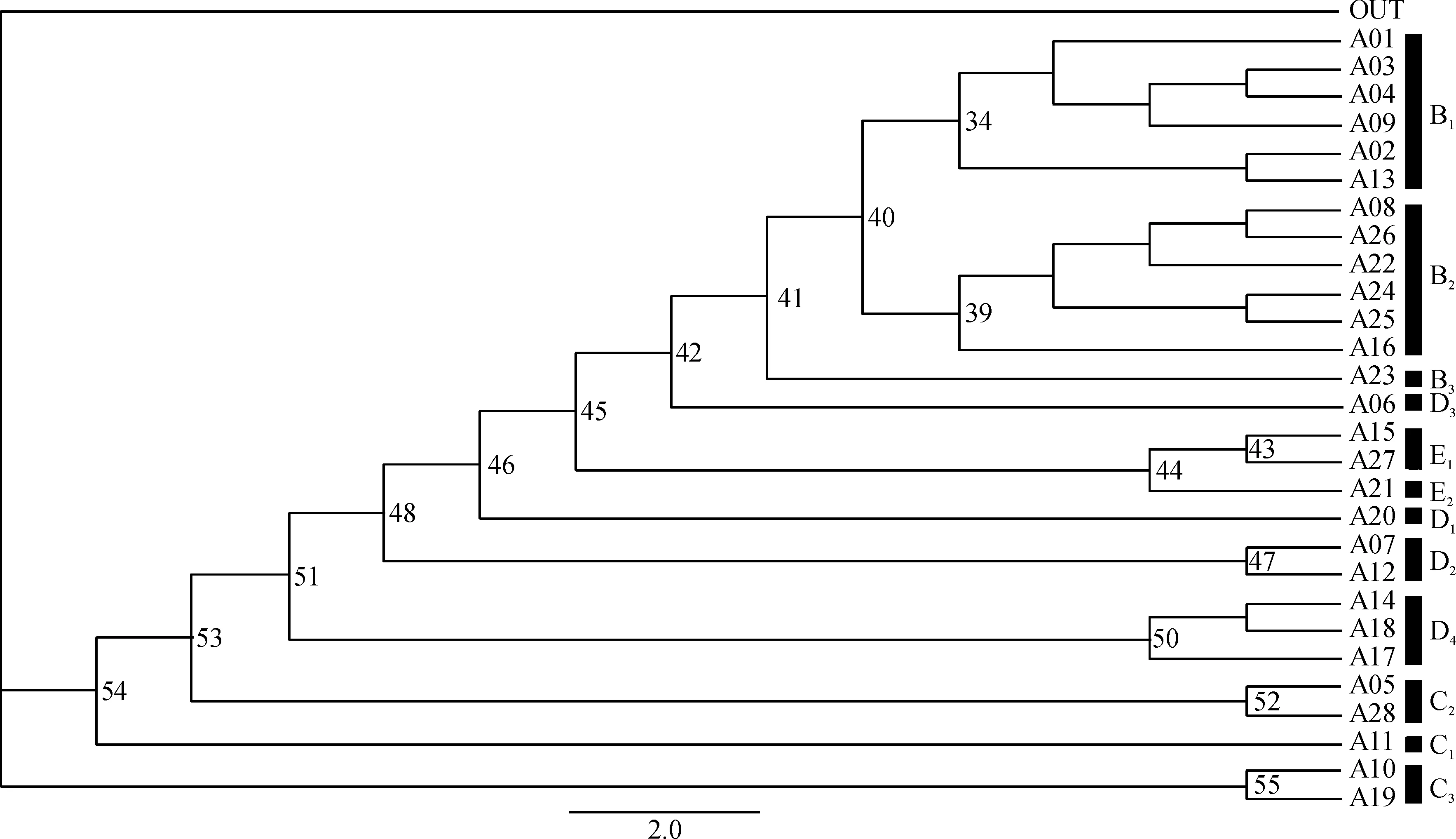
图中的数字为 1 000 次自展置信检验值;A01:淮北平原;A02:洞庭湖平原;A03:江汉平原;A04:浙闽丘陵北段;A05:西部荒漠;A06:秦巴山区;A07:河北平原;A08:滇南丘陵;A09:鄱阳湖平原;A10:内蒙古高原;A11:青藏高原东北部;A12:山西高原;A13:四川盆地;A14:长白山区;A15:川西高原;A16:贵州高原;A17:冀北辽西山地;A18:东北北部平原;A19:西北荒漠、高原;A20:山东丘陵;A21:西藏高原;A22:浙闽丘陵南段;A23:台湾地区;A24:广东沿海丘陵;A25:广西盆地;A26:海南地区;A27:西南山地;A28:陕晋高原;B1:中部地区;B2:南部地区;B3:台湾地区;C2:中北部地区; C3:西部地区,包括内蒙古和新疆;D2:北部地区;D3:秦巴山区;D4:东北地区;E1:西南地区;E2:西藏地区。
图2 特有性简约性分析所得的严格合意支序图
Fig.2 The strict consensus cladogram based on parsimony analysis of endemicity

A01:淮北平原;A02:洞庭湖平原;A03:江汉平原;A04:浙闽丘陵北段;A05:西部荒漠;A06:秦巴山区;A07:河北平原;A08:滇南丘陵;A09:鄱阳湖平原;A10:内蒙古高原;A11:青藏高原东北部;A12:山西高原;A13:四川盆地;A14:长白山区;A15:川西高原;A16:贵州高原;A17:冀北辽西山地;A18:东北北部平原;A19:西北荒漠、高原;A20:山东丘陵;A21:西藏高原;A22:浙闽丘陵南段;A23:台湾地区;A24:广东沿海丘陵;A25:广西盆地;A26:海南地区;A27:西南山地;A28:陕晋高原; B1:中部地区;B2:南部地区;B3:台湾地区;C2:中北部地区; C3:西部地区,包括内蒙古和新疆;D2:北部地区;D3:秦巴山区;D4:东北地区;E1:西南地区;E2:西藏地区。
图3 基于特有性简约分析的中国弄蝶分布格局
Fig.3 Distribution patterns of skippers in China based on parsimony analysis of endemicity
3 讨论与结论
Hsü等[2]根据造山运动群岛模型将中国版图划分为六大地质省区:劳伦/华夏南部和西南边缘省区、西北省区、华中省区、华南省区、西藏省区和环太平洋省区。中国弄蝶的分布格局与这六大地质省区大致重叠。如:南部地区(B2)与华南地质省区基本重合;中部地区(B1)与华中地质省区大致相同;西部地区(C3)(包括内蒙古和新疆)及华北(C2和D2)西南部区域与西北地质省区基本重叠;东北地区(D4)与环太平洋北部省区基本一致;西藏地区(E2)与西藏省区相吻合;西南地区(E1)与劳伦/华夏南部和西南边缘省区基本重叠。
通过比对地质板块分布格局与弄蝶的分布格局可知:弄蝶主要分布在秦岭山脉的东部、藏东南地区、黄河和长江中下游以及南部地区。这些地区自更新世以来,伴随着青藏高原的隆起,经历了深刻的气候变化(东季风的形成,冰期和间冰期的频繁交替)以及河流系统的演变[15-16]。最近的研究表明,在冰期期间,复杂的地形和山区可作为合适的潜在栖息地为温带物种提供相对稳定的微生态系统[17]。中国中部、东部、南部于更新世冰川期间成为动植物的避难所,为生物提供适宜的栖息环境[18-25]。避难所的存在可能对弄蝶的分布格局具有一定的作用,也可能是促进弄蝶种间和种内进一步分化的原因。
不同类群受同一地质历史的影响可能具有相同的分布格局。为检验这一假设,我们将本研究结果与其他类群进行了比较[4-5, 7],结果显示有 5 个区基本一致:中部地区(B1)、南部地区(B2)、西部地区(C3)、北部地区(D2)、东北地区(D4)。同时,不同类群其分布格局亦有差异。张荣祖[4]根据我国已有陆栖脊椎动物的研究,将云南南部山地作为滇南亚区处理,隶属于华南区。Xing等[7]以动物区系内物种数为研究对象,对中国鸟类和哺乳类动物地理区划的19个亚区进行聚类分析,结果显示云南南部山地亚区与其他分支明显分开,独成一支,在此基础上,结合亚区的气候条件、植被类型和地质演化历程,将云南南部山地亚区提升为区,即滇南山区。本研究根据中国弄蝶的分布特点,利用特有性简约分析方法进行聚类,结果显示滇南山地亚区、闽广沿海亚区和海南亚区聚为一支,说明将滇南山地亚区放在华南区是非常合适的,故本研究支持张荣祖[4]提出的视云南南部山地为 1 个亚区的观点。这种差异可能是因研究类群的不同所导致的,也可能与调查不全面有关。
在传统的中国动物地理区划中,台湾一直被视为 1 个亚区即台湾亚区,归属于华南区[4]。在中国生物地理区划研究中,Xie等[5]将台湾及其附近各小岛作为一个整体视为 1 个亚区。在对中国 40 000 种昆虫蜘蛛区系的多元相似性聚类分析研究中,申效诚等[26]提出台湾单独为 1 个分布区。本研究结果显示台湾(A23)单独聚为一支,显示了其物种分布格局的特殊性。台湾是我国东部最大的陆地性岛屿,因印度大陆北缘与亚洲大陆南缘挤压而隆起,其虽为岛屿性陆地,但有丰富的低地、低山及高山环境。小生境气候条件和高山环境为动物提供了合适的居所,因此也被赋予了“避难所”的称号[27-29]。此外,台湾岛属于西太平洋岛弧系的一部分,具备独特的地质和地球物理特征。因此,本研究认为台湾单独成为 1 个分布区比较合理,这一观点与申效诚等[26]的研究结果一致。
西藏在我国动物地理区划中,存在较大分歧。陈宜瑜等[30]从历史时空的角度对青藏高原地区鱼类区系的历史演化过程进行分析,认为高原地区在动物地理区划上应作为独立的区划单元,即为青藏区。王保海等[31]根据西藏昆虫特有种和高山成分以及昆虫区系的垂直带谱,提出青藏高原区为单独的动物地理区,与古北区、东洋区等其他动物地理区并列。Xiang等[6]依据中国及周边国家577种陆生哺乳动物的定量分析研究,提出青藏地区为 1 个亚区。本研究结果显示西藏(A21)独立为一支,与其他分支明显分开,故本研究暂且将西藏区作为单独的 1 个区。由于对西藏地区的划分,仅考虑了物种分布上的特点,建议今后的研究充分考虑当地的自然气候条件、植被类型、不同海拔高度和地质历史的演变历程,结合已有的动植物区划体系多角度、多方位对西藏在动物地理区划中的划分作进一步的探讨。
致谢:感谢华南农业大学农学院昆虫学系王厚帅博士、卢锦明博士研究生给予的支持和帮助!
[1] DINIZ-FILHO J A F. A nice step towards the final frontier[J]. J Biogeogr, 2005, 32(7): 1287-1288.
[2] HSÜ K J, CHEN H. Geologic atlas of China: An application of the tectonic facies concept to the geology of China[M]. Amsterdam :Elsevier, 1999.
[3] 黄汲清,任纪舜,姜春发,等.中国大地构造基本轮廓[J].地质学报,1997, 51(2): 117-135.
[4] 张荣祖. 中国动物地理[M]. 北京: 科学出版社, 1999.
[5] XIE Y, MACKINNON J, LI D. Study on biogeographical divisions of China[J]. Biodivers Conserv, 2004, 13(7): 1391-1417.
[6] XIANG Z, LIANG X, HUO S, et al. Quantitative analysis of land mammal zoogeographical regions in China and adjacent regions[J]. Zool Stud, 2004, 43(1): 142-160.
[7] XING Y, ZHOU L, ZHANG Y, et al. Geographical patterns based on faunal types of breeding birds and mammals in China[J]. Integr Zool, 2008, 3(4): 280-289.
[8] MENG K, LI S, MURPHY R W. Biogeographical patterns of Chinese spiders (Arachnida: Araneae) based on a parsimony analysis of endemicity[J]. J Biogeogr, 2008, 35(7): 1241-1249.
[9] MORRONE J J, ESCALANTE T. Parsimony analysis of endemicity (PAE) of Mexican terrestrial mammals at different area units: When size matters[J]. J Biogeogr, 2002, 29(8): 1095-1104.
[10]BROOKS D R, VAN VELLER M G. Critique of parsimony analysis of endemicity as a method of historical biogeography[J]. J Biogeogr, 2003, 30(6): 819-825.
[11]PORZECANSKI A L, CRACRAFT J. Cladistic analysis of distributions and endemism (CADE): Using raw distributions of birds to unravel the biogeography of the South American aridlands[J]. J Biogeogr, 2005, 32(2): 261-275.
[12]POSADAS P. Distributional patterns of vascular plants in Tierra del Fuego: A study applying parsimony analysis of endemicity (PAE)[J]. Biogeographica, 1996, 72(4): 161-177.
[13]PAUP D L S. Phylogenetic analysis using parsimony (and other methods)[M]. Sunderland: Sinauer Associates,2005.
[14]FELSENSTEIN J. Confidence limits on phylogenies: An approach using the bootstrap[J]. Evolution, 1985,39(4): 783-791.
[15]李吉均. 青藏高原的地貌演化与亚洲季风[J]. 海洋地质与第四纪地质,1999, 19(1): 1-11.
[16]WU Y, CUI Z, LIU G, et al. Quaternary geomorphological evolution of the Kunlun Pass area and uplift of the Qinghai-Xizang (Tibet) Plateau[J]. Geomorphology, 2001, 36(3): 203-216.
[17]QIAN H, RICKLEFS R E. Large-scale processes and the Asian bias in species diversity of temperate plants[J]. Nature, 2000, 407(6801): 180-182.
[18]GAO L M, MÖLLER M, ZHANG X M, et al. High variation and strong phylogeographic pattern among cpDNA haplotypes inTaxuswallichiana(Taxaceae) in China and North Vietnam[J]. Mol Ecol, 2007, 16(22): 4684-4698.
[19]LI S H, YEUNG C K L, FEINSTEIN J, et al. Sailing through the Late Pleistocene: Unusual historical demography of an East Asian endemic, the Chinese Hwamei (Leucodioptroncanorumcanorum), during the last glacial period[J]. Mol Ecol, 2009, 18(4): 622-633.
[20]QIU Y, GUAN B, FU C, et al. Did glacials and/or interglacials promote allopatric incipient speciation in East Asian temperate plants? Phylogeographic and coalescent analyses on refugial isolation and divergence inDysosmaversipellis[J]. Mol Phylogenet Evol, 2009, 51(2): 281-293.
[21]SONG G, QU Y, YIN Z, et al. Phylogeography of theAlcippemorrisonia(Aves: Timaliidae): Long population history beyond late Pleistocene glaciations[J]. BMC Evol Biol, 2009, 9(1): 143.
[22]TIAN B, LIU R, WANG L, et al. Phylogeographic analyses suggest that a deciduous species (OstryopsisdavidianaDecne., Betulaceae) survived in northern China during the Last Glacial Maximum[J]. J Biogeogr, 2009, 36(11): 2148-2155.
[23]FLANDERS J, WEI L, ROSSITER S J, et al. Identifying the effects of the Pleistocene on the greater horseshoe bat,Rhinolophusferrumequinum, in East Asia using ecological niche modelling and phylogenetic analyses[J]. J Biogeogr, 2010, 38(3): 439-452.
[24]HUANG Z, LIU N, LIANG W, et al. Phylogeography of Chinese bamboo partridge,Bambusicolathoracicathoracica(Aves: Galliformes) in south China: Inference from mitochondrial DNA control-region sequences[J]. Mol Phylogenet Evol, 2010, 56(1): 273-280.
[26]申效诚,孙浩,马晓静. 中国 40 000 种昆虫蜘蛛区系的多元相似性聚类分析[J]. 生命科学, 2010, 4(2): 35-40.
[27]SHEN L, CHEN X Y, ZHANG X, et al. Genetic variation ofGinkgobilobaL.(Ginkgoaceae) based on cpDNA PCR-RFLPs: Inference of glacial refugia[J]. Heredity, 2005, 94(4): 396-401.
[28]WU S H, HWANG C Y, LIN T P, et al. Contrasting phylogeographical patterns of two closely related species,MachilusthunbergiiandMachiluskusanoi(Lauraceae), in Taiwan[J]. J Biogeogr, 2006, 33(5): 936-947.
[29]MANCHESTER S R, CHEN Z D, LU A M, et al. Eastern Asian endemic seed plant genera and their paleogeographic history throughout the Northern Hemisphere[J]. J Syst Evol, 2009, 47(1): 1-42.
[30]陈宜瑜,陈毅峰,刘焕章.青藏高原动物地理区的地位和东部界线问题[J]. 水生生物学报,1996, 20(2):97-103.
[31]王保海,王成明,戴万安,等. 西藏昆虫研究综述[J]. 西藏农业科技,1997(4): 4-18.
【责任编辑 霍 欢】
Distribution patterns of skippers (Lepidoptera:Hesperiidae) in China based on parsimony analysis of endemicity
HUANG Qitong, LI Jinsong, ZHUANG Hailing, HUANG Zhenfu, TANG Jing, WANG Min, FAN Xiaoling
(College of Agriculture,South China Agricultural University, Guangzhou 510642,China)
【Objective】 To determine the biogeographical regions of skippers in China based on their distribution, and to infer the causes of the distribution pattern.【Method】 The distribution of 328 species of skippers in China were summarized. China was initially divided into 294 quadrats of 2° latitude×2° longitude. Subsequently, these quadrats were further pooled into 28 areas based on topographical characteristics, vegetation and distribution patterns of all known skippers. Parsimony analysis of endemicity(PAE) was used to classify the 28 areas according to the shared distributional patterns of skippers.【Result】China was found to have ten major biogeographical regions based on the distributional patterns of skippers: central region (B1), southern region (B2), Taiwan region (B3), central northern region (C2), western region (C3), northern region (D2),Qingling-Daba mountains region (D3), eastern northern region (D4), western southern region (E1) and Tibet region (E2). Among them, Taiwan region and Tibet region formed a independent clade respectively. A comparison of the distributional patterns of skippers and geological provinces revealed that the skippers were mainly distributed in Qinba mountains, eastern southern Tibet, southeast Himalayan slope, the middle and lower reaches of the Yellow and Yangtze Rivers, and south China. Compared with the distributional patterns of existing organisms,five distributional regions of skippers were found to be matched: central region (B1), southern region (B2), western region (C3), northern region (D2) and eastern northern region (D4). 【Conclusion】The distributional patterns of skippers in China correspond broadly to geological provinces.
skipper; biogeographical region; China; distribution pattern; geological province; parsimony analysis of endemicity
2016- 04- 19优先出版时间:2016-12-28
黄启通(1990—),男,硕士研究生,E-mail:1508030013@qq.com;通信作者:范骁凌(1966—),女,副教授,博士,E-mail: fanxiaol66@scau.edu.cn
国家自然科学基金(31172136,31471984,40971037)
S511;S502
A
1001- 411X(2017)01- 0069- 07
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20161228.0922.002.html
黄启通,李劲松,庄海玲,等.中国弄蝶科昆虫分布格局的简约性分析[J].华南农业大学学报,2017,38(1):69- 75.
