4个茶树品种自交后代群体遗传结构分析
2017-01-13王让剑孔祥瑞郑国华
杨 军,王让剑,孔祥瑞,郑国华
(福建省农业科学院茶叶研究所/国家茶树改良中心福建分中心,福建 福安 355015)
4个茶树品种自交后代群体遗传结构分析
杨 军,王让剑,孔祥瑞,郑国华
(福建省农业科学院茶叶研究所/国家茶树改良中心福建分中心,福建 福安 355015)
利用EST-SSR对茶树品种紫玫瑰、金牡丹、茗科1号 (金观音)、悦茗香及其自然杂交后代,进行亲缘关系、遗传多样性、群体结构分析。16对EST-SSR标记共检测到等位位点125个,反映位点多态性水平的PIC值平均值为0.617,变幅为0.159~0.840。参试材料的观测杂合度、期望杂合度、Nei′s基因多样性、Shannon信息多样性指数的平均值分别为0.579、0.639、0.632、1.281。结果表明,参试自然杂交后代有较强母本遗传效应,4个群体具有明显的群体遗传组成特征,Structure软件适宜茶树品种群体遗传结构分析。
茶树;自然杂交;EST-SSR;亲缘关系;群体结构
茶树是山茶科山茶属茶组的多年生常绿木本植物[1],且为常异花授粉植物,具有高度杂合性,其较为复杂的群体遗传结构给育种工作造成了较大难度。EST-SSR来自于功能基因,具有多态性高、共显性、技术简单、重复性好等特点,在茶树遗传多样性、亲缘关系分析、群体结构分析、关联分析、种质资源鉴定筛选中广泛应用[2-4]。对材料的遗传多样性与群体结构的解析是物种进化、等位基因发掘、复杂性状关联分析的重要基础[5-7]。刘文等[8]对大白菜与白菜群体的遗传结构分析,表明白菜与大白菜群体间存在基因交流,且群体与材料的来源地、抗性有明显的对应关系。茶树群体结构分析也有大量的研究。王丽鸳等[9]认为龙井群体的遗传分化程度较低。姚明哲等[10]认为江北茶区主要省份间茶树种质的遗传分化程度低。乔婷婷等[11]认为茶树地方品种、选育品种 (系)具有相对独立的群体结构,选育品种 (系)根据亲缘关系的不同形成不同的类群。吴晓梅等[12]认为乌龙茶品种与绿茶品种间的遗传结构存在差异。本试验对高香乌龙茶品种紫玫瑰、茗科1号 (金观音)、金牡丹与悦茗香及其自然杂交后代进行遗传多样性与群体结构分析,以期为杂交后代遗传多样性与标记性状关联分析提供参考价值。
1 材料与方法
1.1 试验材料
参试材料来源于福建省农业科学院茶叶研究所,紫玫瑰、金观音、金牡丹、悦茗香及其自然杂交后代共52个 (每个母本与自然杂交后代数量均为13个),采集各个材料1芽2叶鲜叶后用液氮迅速冷冻处理,保存在-70℃冰箱中备用。
1.2 试验方法
1.2.1 基因组DNA的提取 采用改进的CTAB法[13]提取茶树基因组DNA。用1.0%的琼脂糖凝胶电泳进行茶树基因组分子量大小检测,用756-MC型紫外分光光度计测定茶树基因组DNA的纯度(OD260/280比值)。
1.2.2 引物合成 参照文献 [14]引物序列,序列由上海Sangon公司合成。
1.2.3 PCR扩增和产物鉴定 PCR反应体系为:ddH2O 18.8μL,10×Buffer 2.5μL(Mg2+),dNTP(10mmol·L-1)0.5μL,上、下游primer (10μmol·L-1)各0.5μL,Taq polymerase 0.2 μL(0.5U),模板DNA 1μL。PCR反应于美国ABI-9600型扩增仪上进行,热循环程序为:94℃预变性4min,使模板DNA充分变性,然后进入下列温度循环:94℃变性45s,不同温度条件退火60s,72℃延伸75s,重复35个热循环;72℃延伸10min,最后4℃保存。
引物统一在反向引物 (R)的5′段标记荧光(FAM/TAMRA),由上海百力格生物科技有限公司合成。扩增产物0.5μL,GeneScanTM500 ROXTM0.5μL,HiDi 9μL,混匀后使用ABI公司3730XL进行毛细管电泳。
1.2.4 群体结构分析 使用Struture 2.3.4[15]进行群体结构分析,先计算各材料归属第k亚群的概率Q值。使用混合模型K值从3设置到10模拟群体结构,每一个K值重复5次。将MCMC迭代设为100000次,初始burn-in次数为100000。
1.2.5 数据处理 采用ABI公司的GeneMapper 4.0软件,选择GeneScanTM500ROXTM作为分子量标准,对每一个扩增出的条带记录大小。运用PopGen3.2软件计算Shannon信息指数(I)和Nei′s基因多样性指数(H)。运用NTSYSpc 2.1中的Tree plot程序进行聚类分析。方差分析采用SAS。
2 结果与分析
2.1 引物多态性与等位位点分析
从表1中看出,16对EST-SSR标记在参试材料中共检测到等位位点125个,平均每对引物检测到7.812个,其中最多为TM345为13个,多态性十分丰富;16对引物的PIC值平均为0.617,高于0.5,筛选出来的引物具有较强鉴定能力;根据Nei(1978)提供的公式计算观测杂合度与期望杂合度平均值分别为0.581与0.662,数值比较接近,说明筛选的引物检测到的等位基因在4个群体中分布相对均匀。
2.2 自然杂交后代与母本遗传分析
在相似系数0.76处,52份材料被分为3组,第I组主要为金观音与金牡丹及其自然杂交后代,且金牡丹及其自然杂交后代主要集中在第i组,第II组主要为紫玫瑰及其自然杂交后代,第III组主要为悦茗香及其自然杂交后代 (图1)。参试自然杂交后代都与母本亲缘关系近,且方差分析结果表明金牡丹、悦茗香与其自然杂交后代的相似系数显著高于金观音与紫玫瑰 (表2),说明参试的自然杂交后代具有较强母本遗传效应,且参试金牡丹、悦茗香的自然杂交后代母本遗传效应强于金观音与紫玫瑰。
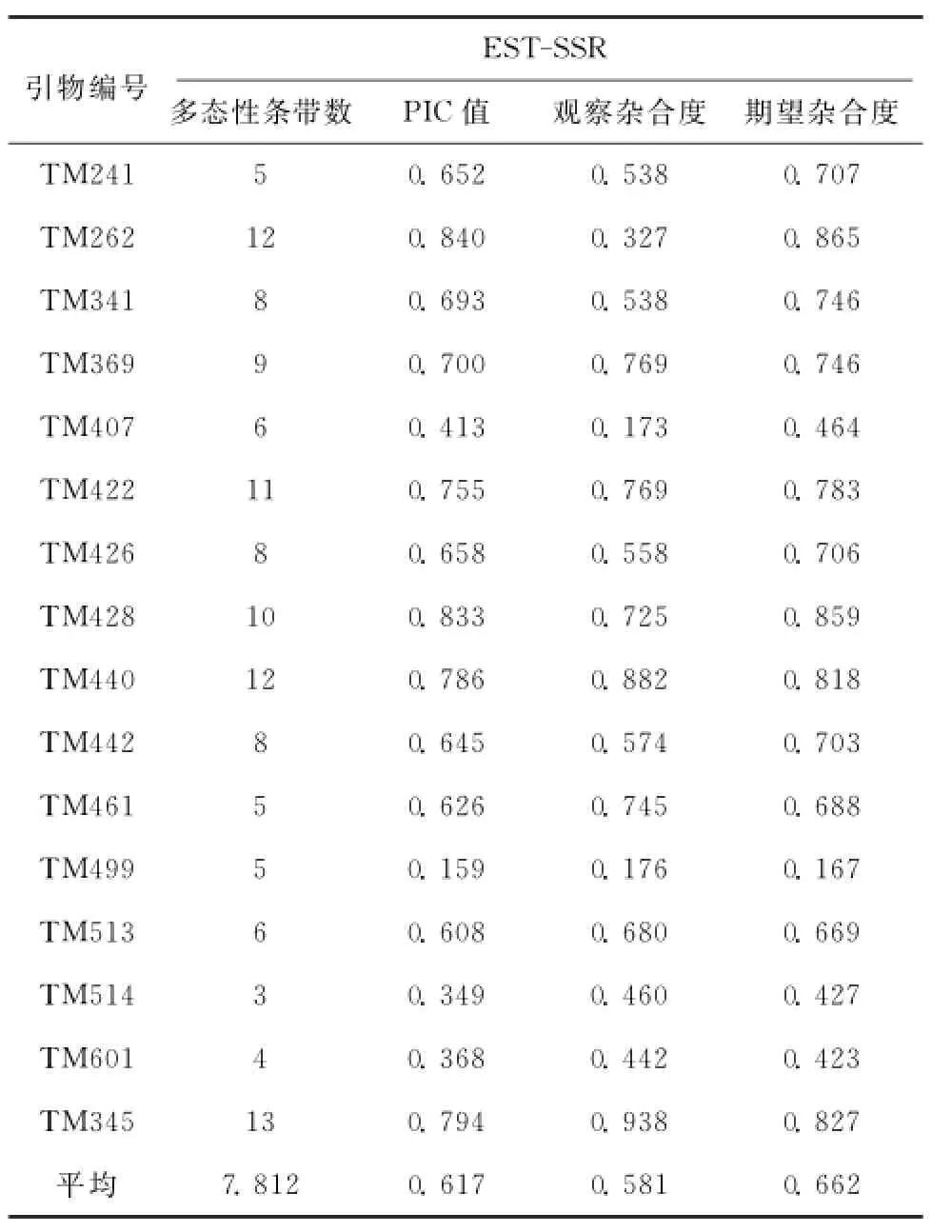
表1 参试材料的扩增结果Table 1 Amplified results of tested materials
2.3 群体间遗传结构分析
利用Structure软件对参试材料群体结构进行了系统解析 (图2)。当类群数 (K值)在2~7变化时,△K值在K=4时达到最大值,表明52份参试材料可以划分为4个类群。当以类群属性比率(研究材料的遗传组分在各类群上的分布百分比,≥50%的遗传组分称为类群属性比率,<50%的遗传组分则称为非类群属性比率[6])作为划分依据时,52份材料中,有49份(94.3%)被划分到4个类群中的一个,而剩余的3份 (5.8%)则没有明确的类群归属特性,形成了一个混合类群P(表3)。依据各类群所包含材料的系谱等信息,这4个类群依次确定为S1类群(25.0%)、S2类群(19.2%)、S3类群(26.9%)与S4类群(23.1%)。
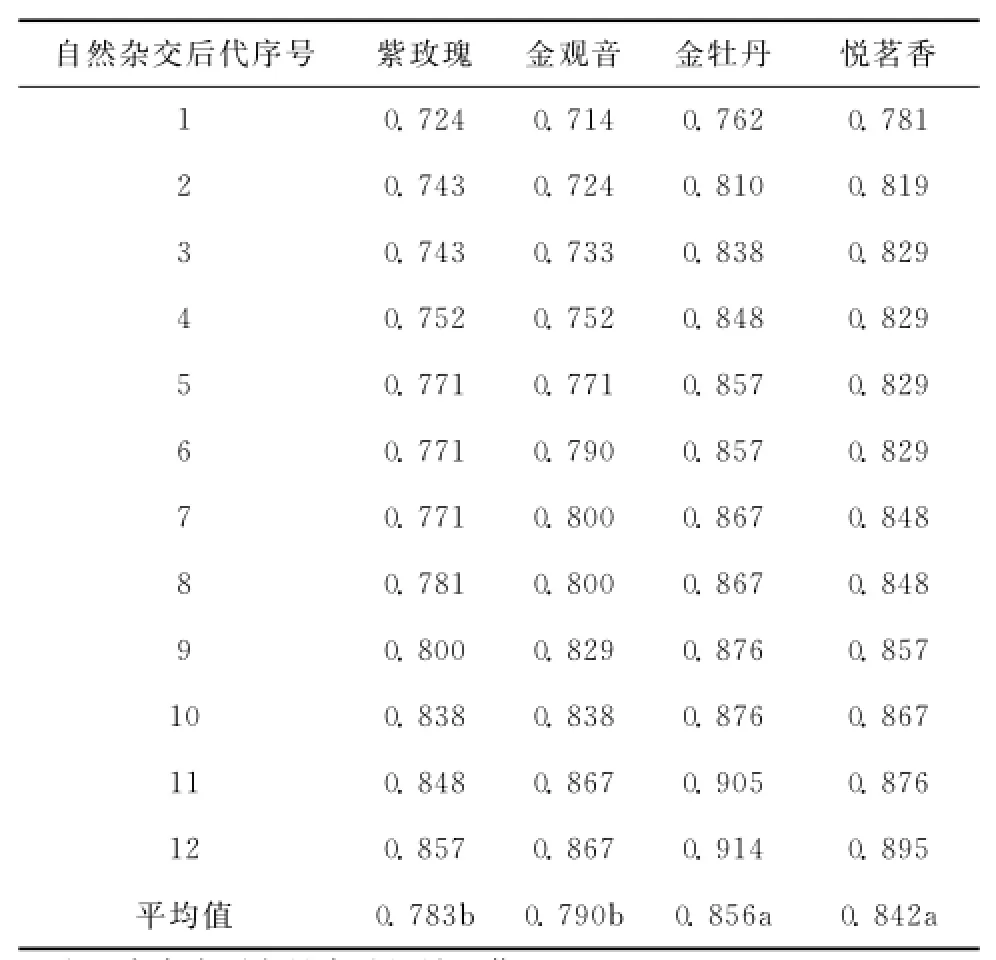
表2 不同母本与其自然杂交后代的相似系数比较Table2 Similarity coefficients on various females and theirnatural hybrid progenies

图1 遗传相似系数聚类图Fig.1 Dendrogram of genetic similarity
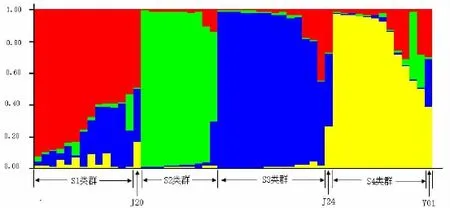
图2 假设K=4时群体按照所估计的成分 (Q)值的结构分析结果Fig.2 Structures under assumed cluster number(k=4)and sorted by estimated membership coefficients of individual samples in 4 clusters
2.4 群体间聚类与遗传多样性情况
软件PopGen计算按来源分类群的相似系数和按Structure软件结果分类群的相似系数,并用NTSYS绘制UPGMA图3。在相似系数为0.80处,2种分群聚类结果都显示金观音、金牡丹、紫玫瑰及其自然杂交后代聚类在一起,悦茗香及其杂交后代群体单独聚类。从表4中看出,按材料来源分类群,4个类群Shannon信息指数的方差分析未达到显著差异;而按Structure软件结果分类群,5个类群在Shannon信息指数的方差分析达到显著差异;说明按Structure软件结果分类群,类群间遗传组成特征更显著。

表3 52份材料的群体结构分析结果统计Table 3 Statistic analysis on population structures of 52 samples

图3 群体遗传相似系数聚类图Fig.3 Dendrogram on genetic similarity among groups
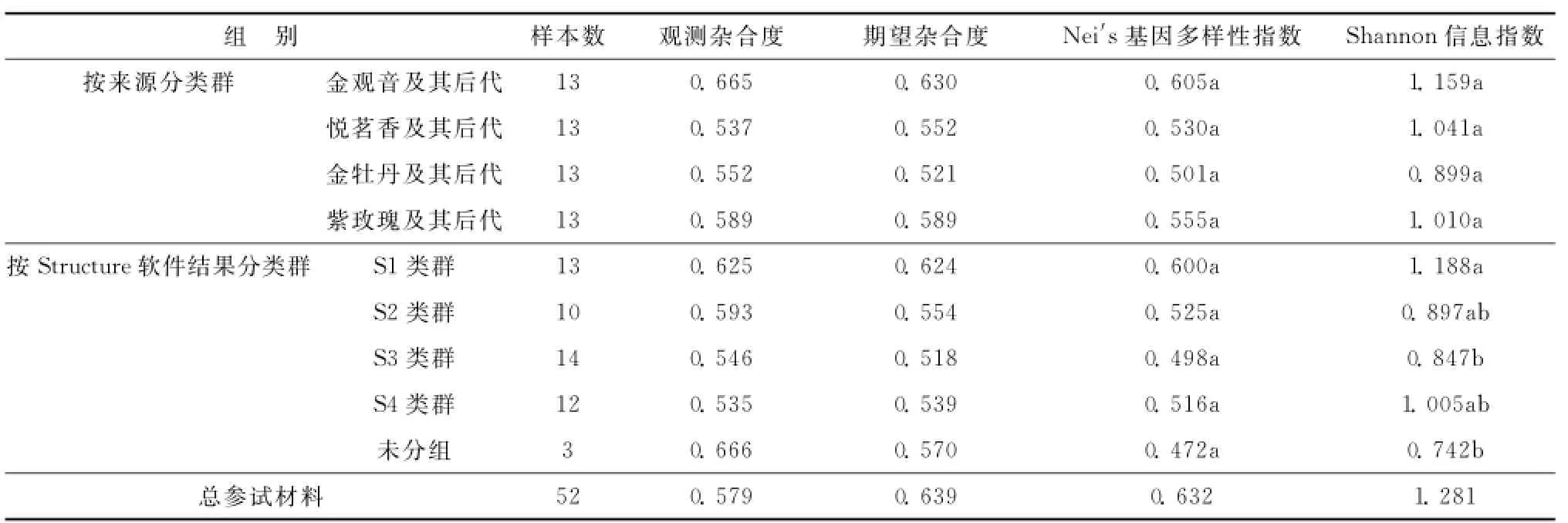
表4 群体遗传多样性比较Table 4 Genetic diversity of various groups
3 讨论
金观音、金牡丹、紫玫瑰都为铁观音与黄棪的人工杂交后代[16],亲缘关系较近[17]。Struture软件将参试材料分为4个类群,其中悦茗香及其自然杂交后代、紫玫瑰与其自然杂交后代类群,与NTSYS聚类结果相似,金观音、金牡丹及其自然杂交后代类群与NTSYS聚类结果有所不同,NTSYS聚类仅将金牡丹、金观音及其杂交后代分为1个类群。Struture群体结构分析显示,将金牡丹、金观音及其杂交后代分为2个类群,其中类群S1为金观音、金牡丹、紫玫瑰的自然杂交后代,参试自然杂交后代受母本遗传效应影响较大,类群S1反映的类群属性可能由于金观音、金牡丹、紫玫瑰具有相同的铁观音或黄棪亲本来源,与参试材料来源背景更为相符。综合分析,Struture软件能更好的分析参试材料的群体遗传结构,与魏世平[5]分析中国栽培大豆群体结构不同分类方法比较的结果一致。
刘金[18]等认为小扁豆种质资源的PCA分析和Structure遗传结构分析的结果相互间完全吻合。按材料来源分类群和Struture软件结果分类群结果进行PopGen检测,看出对于亲缘关系来源比较近的群体,用Struture软件结果进行分类优于按材料来源对参试材料进行分类,对于来源亲缘关系比较远的群体,2种分类群方法结果差异不大,如本研究中对紫玫瑰、悦茗香及其杂交后代的分类。
[1]虞富莲.论茶树原产地和起源中心[J].茶叶科学,1986,6 (1):1-8.
[2]姜燕华,段云裳,王丽鸳,等.福建茶树品种的SSR分析及其人为选择的影响[J].浙江林业科技,2010,30(3):12-16.
[3]王旭,董丽娟,段继华,等.84个茶树品种遗传多样性及亲缘关系的SSR分析[J].湖南农业大学学报,2011,37(3):260-266.
[4]周萌,李友勇,孙雪梅,等.基于EST-SSR标记的云南野生茶树遗传多样性分析[J].江苏农业科学,2013,41(12):22-27.
[5]魏世平,刘晓芬,杨胜先,等.中国栽培大豆群体结构不同分类方法的比较[J].南京农业大学学报,2011,34(2):13-17.
[6]刘志斋,吴迅,刘海利,等.基于40个核心SSR标记揭示的820份中国玉米重要自交系的遗传多样性与群体结构 [J].中国农业科学,2012,45(11):2107-2138.
[7]陈斐,魏臻武,李伟民,等.基于SSR标记的苜蓿种质资源遗传多样性与群体结构分析[J].草地学报,2013,21(4):759-768.
[8]刘文,萧凤回,武剑,等.大白菜和白菜群体的遗传结构分析[J].云南农业大学学报,2011,26(2):156-163.
[9]王丽鸳,姜燕华,段云裳,等.基于SSR分子标记的龙井群体种的遗传多样性及遗传分化研究 [J].茶叶科学,2011,31 (1):40-44.
[10]姚明哲,刘振,梁月荣,等.利用EST-SSR分析江北茶区茶树资源的遗传多样性和遗传结构[J].茶叶科学,2009,29 (3):243-250.
[11]乔婷婷,马春雷,陈亮,等.浙江省茶树地方品种与选育品种遗传多样性和群体结构的EST-SSR分析 [J].作物学报,2010,36(5):744-753.
[12]吴晓梅,姚明哲,陈亮,等.利用EST-SSR标记研究适制绿茶与乌龙茶品种的遗传多样性与遗传结构 [J].茶叶科学,2010,30(3):195-202.
[13]金基强,崔海瑞,龚晓春,等.用EST-SSR标记对茶树种质资源的研究[J].遗传,2007,29(1):103-108.
[14]Ma Jian-qiang,Yao Ming-zhe,Ma Chun-lei,et al.Construction of a SSR-based genetic map and identification of QTLs for catechins content in tea plant(Camellia sinensis) [J].PLOS ONE,2014,9(3):e93131.
[15]Prichard J K,Stepphen M,Donnelly P.Inference of population structure using multilocus genotype data.Genetics,2000,155:945-959.
[16]郭吉春,杨如兴,叶乃兴,等.10个茶树高优新品种的育成与应用[J].福建茶叶,2003,(3):19-21.
[17]王让剑,郭吉春,杨军,等.应用RAPD和EST-SSR分析铁观音与黄棪杂交一代的遗传差异 [A].第六届海峡两岸茶业学术研讨会论文集,2010:16-19.
[18]刘金,关建平,徐东旭,等.小扁豆种质资源SSR标记遗传多样性及群体结构分析[J].作物学报,2008,34(11):1901-1909.
Genetic Structure of Natural Hybrids of Four Tea Varieties
YANG Jun,WANG Rang-jian,KONG Xiang-rui,ZHENG Guo-hua
(Tea Research Institute,Fujian Academy of Agricultural Sciences/Fujian Branch,National Center for Tea Improvement,Fu’an,Fujian355015,China)
EST-SSR was used to analyze the genetic polymorphism,relationship,and population structure of Zimeigui,Jinmudan,Mingke 1(Jinguanyin),Yuemingxiang,and the natural hybrids of Zimeigui,Jinmudan,Jinguanyin,and Yuemingxiang.A total of 125alleles were amplified using 16pairs of EST-SSR primers.The average PIC was 0.617,varying from 0.159to 0.840.The average observed heterozygosity,expected heterozygosity,Nei′s genetic diversity,Shannon information diversity index among them were 0.579,0.639,0.632,and 1.281,respectively.A strong maternal parental effect seemed apparent as shown by the genetics of the natural hybrids.The population structures of the 4groups had distinctive genetic characteristics.It was concluded that the software applied could be adequately used for analyzing the genetic structures of tea populations.
tea plant;natural hybrids;EST-SSR;genetic relationship;population structure
S571.1;Q311
:A
:2096-0220(2016)02-0059-04
2016-02-23初稿;2016-05-04修改稿
福建省现代农业 (茶叶)产业技术体系 (2014~2016);农业部福建茶树及乌龙茶加工科学观测实验站 (2011~2015)。
杨军(1981-),男,硕士,助理研究员,从事茶树种质资源与遗传育种研究。E-mail:yangjun007@qq.com
