不同恢复群落的山地常绿落叶阔叶混交林优势种及主要伴生种种群结构与动态
2017-01-13艾训儒
陈 斯,艾训儒*,姚 兰,2,林 勇,黄 伟,陈 俊
(1.湖北民族学院 林学园艺学院,湖北 恩施 445000;2.北京林业大学 自然保护区学院,北京 100083)
不同恢复群落的山地常绿落叶阔叶混交林优势种及主要伴生种种群结构与动态
陈 斯1,艾训儒1*,姚 兰1,2,林 勇1,黄 伟1,陈 俊1
(1.湖北民族学院 林学园艺学院,湖北 恩施 445000;2.北京林业大学 自然保护区学院,北京 100083)
山地常绿落叶阔叶混交林是我国特有的自然植被类型和天然林类型,为研究山地常绿落叶阔叶混交林优势种及主要伴生种群结构和动态,揭示其生物多样性形成及维持机制,作者依托湖北恩施森林生态系统国家定位观测研究站的建设,对木林子自然保护区不同恢复群落的山地常绿落叶阔叶混交林开展了植物群落调查,从物种组成、径级结构和垂直结构等方面分析了优势种及主要伴生种群的群落特征.结果表明:3个恢复群落中,DBH≥1 cm的木本植株分别有148种6 970株、176种4 533株和149种4 048株,隶属于14科23属、19科26属和16科25属;从整体上看,3个恢复群落优势种及主要伴生种的物种丰富度随垂直层自上而下呈现“单峰型”,个体多度随垂直层降低而增加,平均树高分别为5.94、5.21和4.61 m;群落物种丰富度整体上均随径级增大而下降,个体多度均呈倒“J”型分布,小径级个体数量多,平均胸径分别为4.18、4.35和4.48 cm.总之,木林子自然保护区不同恢复群落的山地常绿落叶阔叶混交林群落物种组成丰富,多数种群自然更新良好,群落整体上正处于演替进程中.
山地常绿落叶阔叶混交林;木林子自然保护区;自然恢复;垂直结构;径级结构
植物群落是一定地段内不同植物在长期历史过程中逐渐形成的生态复合体[1],是由生物因子和非生物因子长期相互作用而形成.群落结构是植物群落的基本属性,可反映群落对环境的适应、动态和机能,是认识群落组成及动态变化的基础[2-3].山地常绿落叶阔叶混交林的生物多样性较高,群落结构复杂,这为研究此类森林群落的结构和动态造成了很大难度,而优势种群对群落结构和群落环境起明显的控制作用.因此,对山地常绿落叶阔叶混交林优势种及主要伴生种群结构和动态研究,有助于认识整个森林群落结构,揭示群落更新趋势、稳定性和演替规律[4-7],为阐明此类森林群落的动态变化规律和生物多样性维持机制提供理论依据.
山地常绿落叶阔叶混交林是我国特有的自然植被类型和天然林类型[8],是木林子自然保护区主要植被类型之一.目前的有关研究主要涉及群落物种组成[9]、木本植物生物多样性[10-11]等,极少涉及优势种群结构和动态.本文在建立森林固定监测样地和植被调查的基础上,首次分析了不同恢复群落山地常绿落叶阔叶混交林的优势种及主要伴生种群和与其相关的结构特征及演替趋势,旨在能更好地了解山地常绿落叶阔叶混交林群落结构和动态,为可持续经营和合理管理此类森林提供重要参考,为人工群落生境确定和物种选择与搭配提供科学依据.
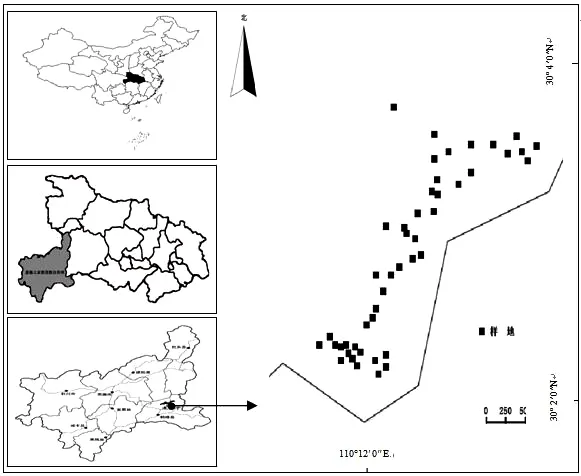
图1 木林子自然保护区地理位置及样地分布图Fig.1 Diagram of thegeographic location and sample plots distribution in Mulinzi Nature Reserve
1 研究区及其自然概况
木林子国家级自然保护区位于湖北省鹤峰县东北部,地理坐标29°55′59″~30°10′47″ N,109°59′30″~110°17′58″ E,海拔为1 100~2 095.6 m,保护区面积20 838 hm2.山地常绿落叶阔叶混交林广泛分布于1 200~1 800 m的海拔梯度上,区域内既有保存完好的原生老龄林(old growth forest,OG),也有处于不同恢复群落(自然恢复20 a、自然恢复35 a)的次生林.境内雨量充沛,年均降水量为1 700~1 900 mm,但季节分配不均;气候温暖湿润,年均气温15.5℃,年均相对湿度为82%;日照充足,无霜期270~279 d;属于中亚热带湿润季风气候.土壤类型多样,自然土壤沿海拔由低到高有山地黄壤、山地黄棕壤和山地棕壤等,土壤pH值在4.5~6.5之间.
2 研究方法及数据处理
2.1 样地设置与植被调查
选取山地常绿落叶阔叶混交林集中分布的典型地段,分别在自然恢复100 a以上的老龄林(OG)、80年代初期皆伐后自然恢复35a的次生林和90年代中期皆伐后自然恢复20a的次生林中,用全站仪设置15个面积为20 m×20 m的固定监测样地(共45个),样地四角均埋设水泥桩作永久标记.按插值法将每个20 m×20 m的样地用红色包装绳分隔为4个10 m×10 m的样方(共180个)和16个5 m×5 m的样块(共720个).
以每个样地为基本调查单元,对胸径(DBH)≥1 cm的所有木本植物进行挂牌、测定点标记和测树因子检测(个体的编号、种名、胸径、树高、萌生情况).以样地西南角为坐标原点,测定每个个体的x轴和y轴.野外不能鉴别的物种通过采集标本到室内查阅相关植物志或请植物分类学专家进行鉴别.
2.2 优势种及主要伴生种群的确定
重要值(IV)是物种的综合数量指标,能表征物种在群落中的地位和作用,可反映物种在群落中的优势程度.本文以物种的IV为数量指标,选取3个恢复群落中IV排列前30的物种作为优势种及主要伴生种(见表2),并进行结构和动态分析.IV的计算公式为[12]:
IV=(RA+RF+RP)/3
式中:RA、RF、RP分别为相对密度、相对频度和相对显著度,a、f、p对应某物种的密度、频度和显著度.

2.3 种群结构划分与数据处理
根据山地常绿落叶阔叶混交林树木的高度特征,按照上限排外法,将优势种群的高度级划分为5个垂直层次:Ⅰ层≥25 m、Ⅱ层 20~25 m、Ⅲ层10~20 m、Ⅳ层5~10 m、Ⅴ层<5 m.
根据山地常绿落叶阔叶混交林物种生长周期,同样按照上限排外法,将优势种群的径级划分为10个等级:Ⅰ级<5、Ⅱ级5~10、Ⅲ级10~15、Ⅳ级15~20、Ⅴ级20~25、Ⅵ级25~30、Ⅶ级30~35、Ⅷ级35~50、Ⅸ级50~100、Ⅹ级≥100 cm.
采用时间代替空间的方法分析不同恢复群落优势种及主要伴生种群的年龄结构,并用多重比较(Tukey-Kramer HSD)分析它们在不同恢复群落的个体高度、个体胸径的差异.
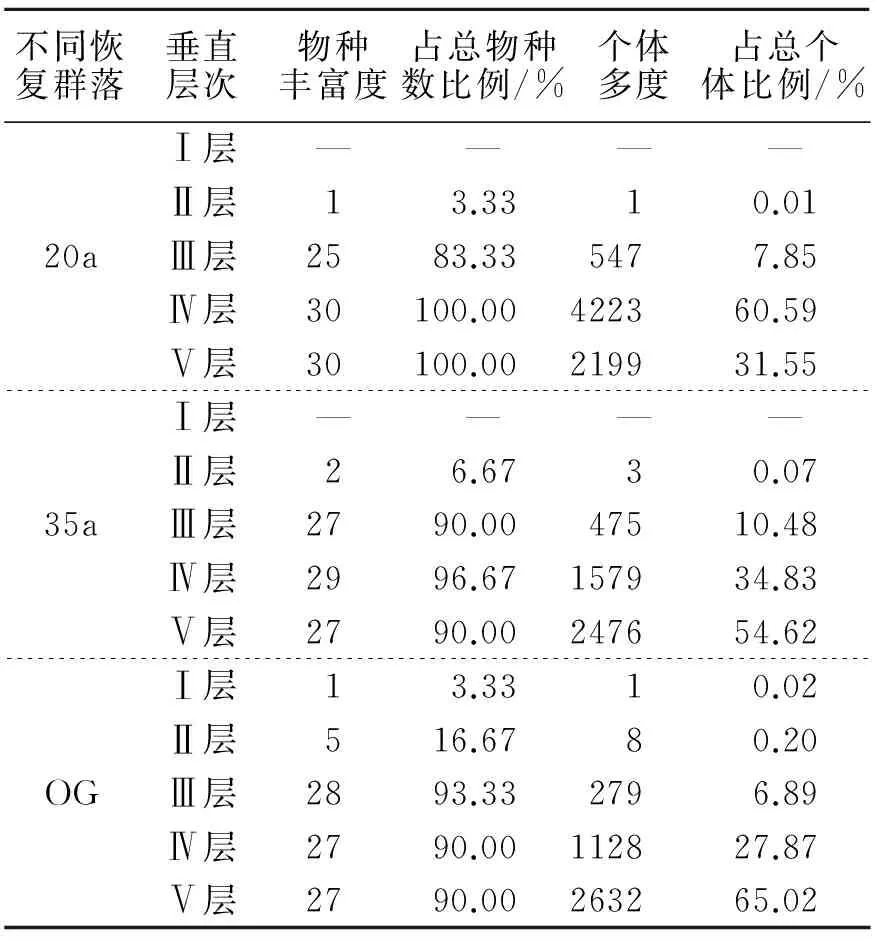
表1 不同恢复群落优势种及主要伴生种的垂直结构Tab.1 Vertical structure of dominant and associated populations in different recovery communities
3 结果与分析
3.1 不同恢复群落优势种及主要伴生种群的物种组成
统计分析可知,3个恢复群落中DBH≥1 cm的木本植物分别有148种、176种和149种.其中,自然恢复20a群落的优势种群占20.3%,隶属14科23属;壳斗科(Fagaceae)植物最多(6种),其次是蔷薇科(Rosaceae)植物(4种);单种及寡种科植物7种.自然恢复35a群落的优势及主要伴生种占17.0%,隶属19科26属;山矾科(Symplocaceae)植物最多(4种),其次是壳斗科、山茱萸科(Cornaceae)和樟科(Lauraceae)(均有3种);单种及寡种科植物有13种.OG群落中优势种占20.1%,隶属16科25属;植物种类最多的是壳斗科(6种),其次是山矾科(5种)、山茶科(Theaceae)(3种);10种为单种及寡种科植物.
3.2 不同恢复群落优势种及主要伴生种群的垂直结构
3.2.1 自然恢复20a群落 物种丰富度随垂直层自上而下呈快速增长趋势(表1).Ⅲ层有25种,占群落优势种及主要伴生种总种数的83.33%,以壳斗科植物(5种)最多;落叶树种(20种)占该层物种数的80%.Ⅳ、Ⅴ层都有30种优势种及主要伴生种,均以壳斗科植物(5种)为主,落叶树种(23种)占优势地位.
个体多度由垂直层自上而下呈现“单峰型”,即个体多度在Ⅳ层(4 223株)达到峰值,占优势种及主要伴生种总个体数的60.59%;个体数超过500株的物种依次是川陕鹅耳枥(688株)、半边月(566株)、青冈(558株).Ⅴ层的个体多度(2 199株)占群落总个体数的31.55%,个体多度最多的物种是半边月(272株),其次是青冈(257株).
3.2.2 自然恢复35a群落 物种丰富度随垂直层自上而下呈现“单峰型”,即Ⅳ层的物种丰富度(29种)较高,占优势种及主要伴生种总种数的96.67%,山矾科植物(4种)最多;落叶树种(17种)所占比例为58.62%,低于自然恢复20 a群落.Ⅲ、Ⅴ层都有27种,都以山矾科植物(4种)和山茱萸科植物(3种)为主,但物种组成并不完全相同.
个体多度随垂直层降低而增加.H≥20 m的物种有亮叶桦(2株)和合欢(1株),占优势种及主要伴生种总个体数的0.07%.Ⅲ层个体数占优势种及主要伴生种总个体数的10.48%,川陕鹅耳枥(80株)、亮叶桦(74株)分别占该垂直层个体数的16.84%和15.58%.Ⅳ、Ⅴ层共占群落总个体数的89.45%,个体数位居前3的是山矾、川陕鹅耳枥和翅柃.
3.2.3 OG群落 物种丰富度随垂直层自上而下也呈现“单峰型”,Ⅲ层物种丰富度(28种)占优势种及主要伴生种总种数的93.33%,壳斗科植物最多(6种),其次是山矾科植物(5种);落叶树种(13种)占该层物种数的46.43%.Ⅳ、Ⅴ层均有27种,有26种物种相同,单种及寡种科种植物较多,表明群落主要伴生种及主要伴生种在空间生态位上产生了一定程度的分化.
个体多度也随垂直层降低而增加.H≥25 m的物种(白辛树1株)占优势种及主要伴生种总个体数的0.02%.粉椴(3株)、锥栗(2株)、亮叶桦(1株)、香椿(1株)和米心水青冈(1株)的树高达到Ⅱ垂直层(20 m≤H<25 m),占优势种及主要伴生种总个体数的0. 20%.Ⅲ层(279株)占优势种及主要伴生种总个体数的6.89%,其中青冈(107株)占该垂直层个体数的38.35%.Ⅳ、Ⅴ层个体数共占优势种及主要伴生种总个体数的92.89%,青冈、翅柃和黄丹木姜子的个体数量较多.
3.3 不同恢复群落优势种及主要伴生种群的径级结构
3.3.1 自然恢复20a群落 物种丰富度随径级增大呈单调下降趋势(图2).DBH<10 cm的物种占优势种及主要伴生种总种数的96.67%以上,仅4种(川陕鹅耳枥、光叶水青冈、灰柯和小叶青冈)物种的DBH≥25 cm,群落最大DBH是灰柯种群(DBHmax=44 cm),表明群落小径级木较多.有20%的物种仅分布于Ⅰ、Ⅱ径级,如茶荚蒾只在Ⅰ径级中分布;26.67%的物种连续分布在前5个径级,说明多数物种的径级分布范围狭窄.
个体多度整体上呈倒“J”型分布.Ⅰ径级(DBH<5 cm)个体占优势种及主要伴生种总个体的73.29%,DBH<10 cm的个体(6 599株)占94.68%,DBH≥30 cm的个体(2株)仅占优势种及主要伴生种总个体的0.03%.结果表明,小径级木所占比例较高,种群自然更新良好,群落正处于演替进程中.
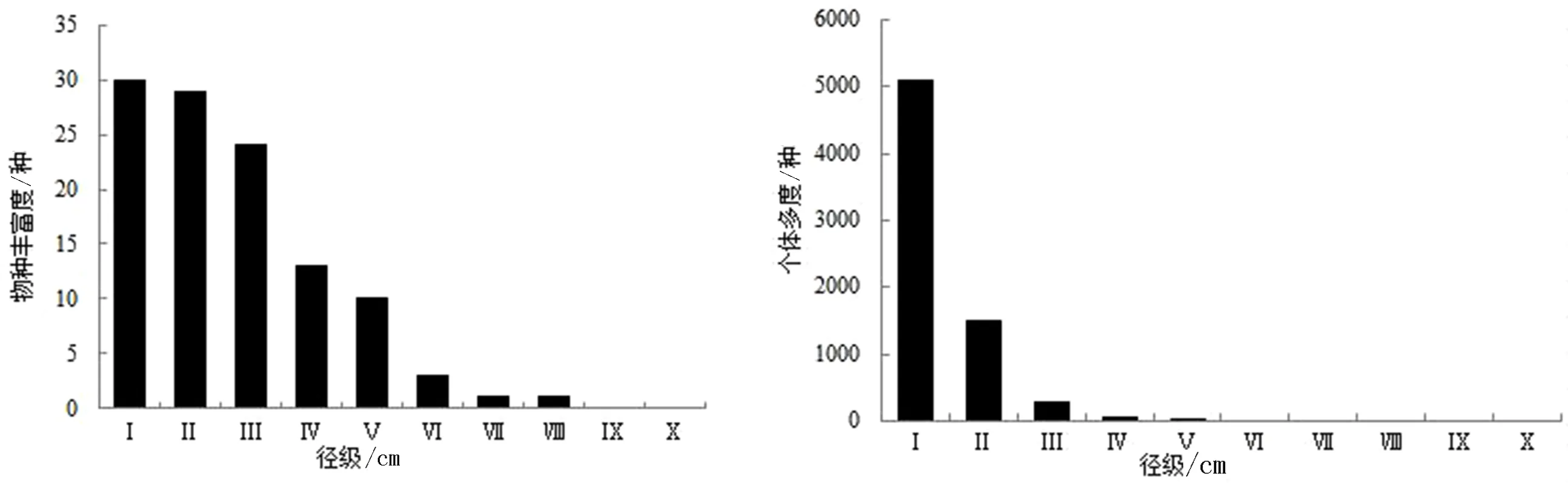
图2 自然恢复20a群落中不同径级的物种丰富度及个体多度Fig.2 Species richness and individual abundance of different diameter classes in 20a of natural recoverystages
3.3.2 自然恢复35a群落 物种丰富度整体上随径级增大而下降.Ⅱ径级分布有大叶杨和盐肤木种群,其物种丰富度高于Ⅰ径级,说明这两种群林下更新受抑制,仅一些DBH较大且对环境适应能力及对资源利用较强的个体存活.个体仅分布在Ⅳ径级范围内(DBH<20 cm)的物种(11种)占优势种及主要伴生种总种数的36.67%,常绿树种10种占总种数的33.33%;DBHmax(杉木)为47.7 cm.可见,群落小径级木和中径级木比例较高,常绿树种逐渐兴起.有10%的物种连续分布在前8个径级,说明优势种及主要伴生种群的径级分布范围在不断扩大.
个体多度整体上呈倒“J”型分布.76.17%的个体分布在Ⅰ径级范围内,前3个径级(DBH<15 cm)共有4 328株,占优势种及主要伴生种总个体数的95.5%.DBH≥30 cm的个体有20株,占优势种及主要伴生种总个体数的0.44%,以锥栗(7株)为主,其次是青冈(4株)和亮叶桦(4株).

图3 自然恢复35a群落中不同径级的物种丰富度及个体多度Fig.3 Species richness and individual abundance of different diameter classes in 35a of natural recoverystages
3.3.3 OG群落 物种丰富度随径级增大呈波动式下降,共有3次峰值.Ⅰ径级有28种物种,占优势种及主要伴生种总个体数的93.33%,出现第1次峰值;有落叶树种12种、常绿树种18种.2、3次峰值分别出现在Ⅵ和Ⅷ径级,均有12种物种,但物种组成并不完全相同,说明该群落优势种及主要伴生种群的径级分布范围较宽且有明显的间断现象.DBH≥100 cm的物种共有2株(香椿和锥栗),群落DBHmax为105 cm,说明群落具有一定的古老性.此外,群落有珙桐(24株)、白辛树(6株)等珍稀濒危植物和国家重点保护植物,说明该群落结构复杂,而复杂的结构孕育了更多的物种.
个体多度整体上也呈倒“J”型分布.Ⅰ径级有3 048株,占优势种及主要伴生种总个体数的75.30%;超过90.34%的优势种及主要伴生种群DBH<10 cm.仅分布在前4个径级的物种有12种,其个体数最多的是青冈(659株)、其次是翅柃(537株)和尖连蕊茶(444株),均为常绿树种.DBH≥50 cm的个体有20株,占优势种及主要伴生种总个体数的0.49%,以锥栗(9株)和香椿(3株)为主.
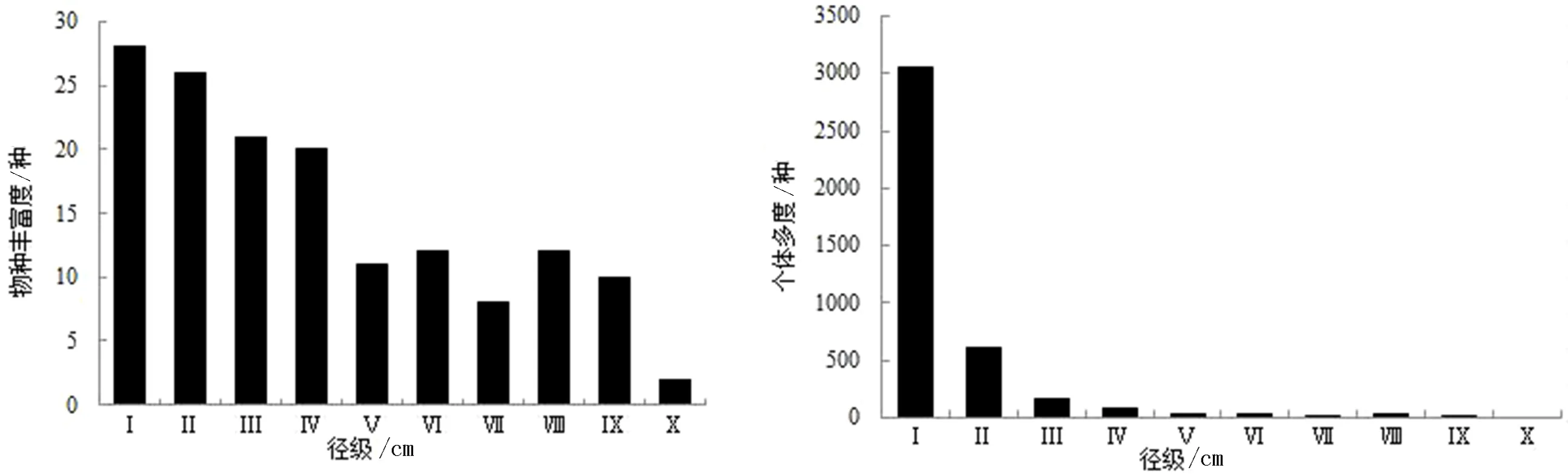
图4 OG群落中不同径级的物种丰富度及个体多度Fig.4 Species richness and individual abundance of different diameter classes in old growth foreststages
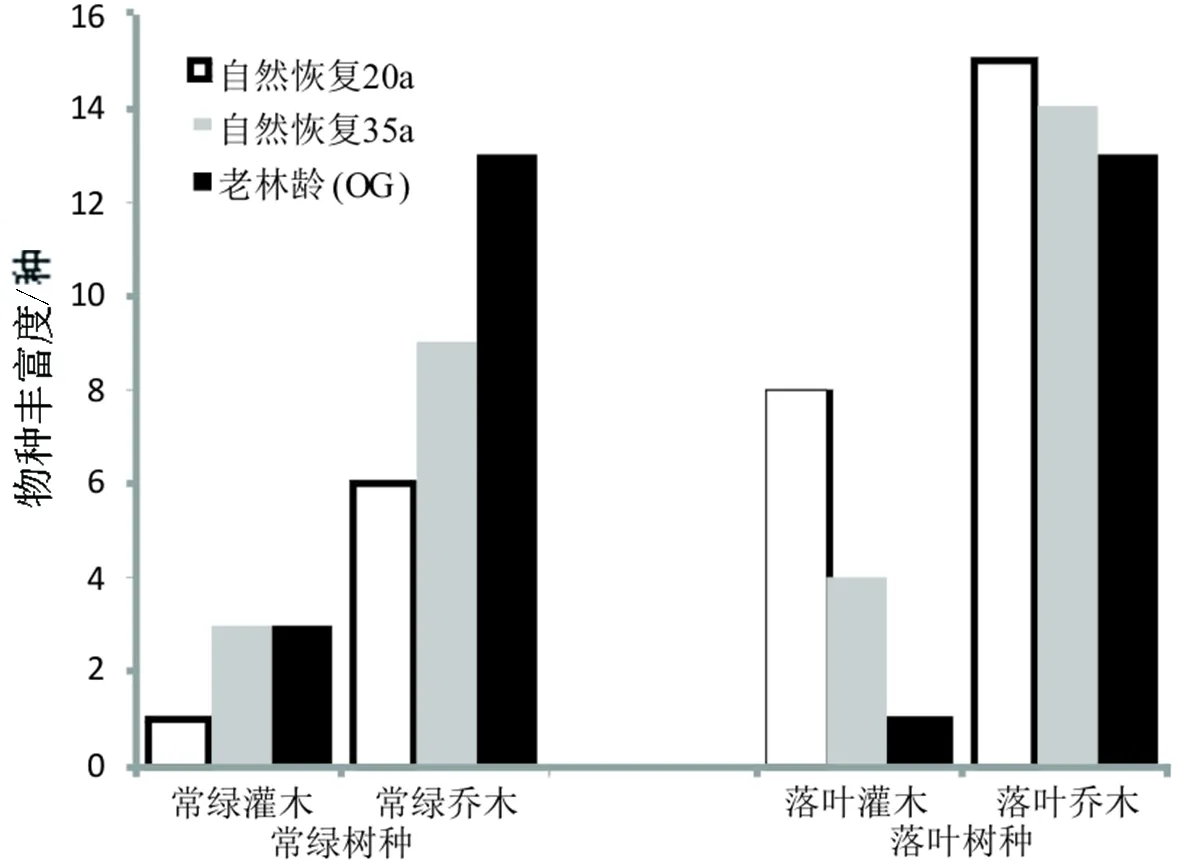
图5 不同恢复群落优势种及主要伴生种群常绿与落叶树种的物种丰富度Fig.5 Species richness of dominantand associated populations of evergreen and deciduous species in differentrecovery communities
3.4 不同恢复群落优势种及主要伴生种群的生活型变化
不同恢复群落中,落叶乔木的物种变化分别为15种、14种、13种,落叶灌木的物种变化分别为8种、4种、1种,均随自然恢复年限增加而减少;常绿乔木的物种变化分别为6种、9种、13种,常绿灌木的变化分别为1种、3种、3种,整体上都随自然恢复年限增加而增加.
4 结论与讨论
4.1 结构特征
4.1.1 垂直结构 就以往研究来看,无论是对大样地[13-14],还是小样地[15-16]的群落垂直结构研究,其林冠层高度一般不超过为20 m.本文按树高将木林子自然保护区山地常绿落叶阔叶混交林优势种及主要伴生种的垂直结构划分为5个垂直层次,其中Ⅰ层H≥25 m,说明该区域山地常绿落叶阔叶混交林的垂直结构较复杂.
3个恢复群落中,多数物种的H<20 m,H≥20 m的个体分别占群落优势种及主要伴生种总个体数的0.01%、0.07%和0.22%,表明树高20 m是山地常绿落叶阔叶混交林物种在高度上的一个分界点,而高于20 m是该类型群落建群种的一个必备属性,这与以往多数研究结果相一致[16-18].
3个恢复种群中,青冈、山矾和翅柃等物种的个体多度随垂直层次降低而增加,说明植株个体主要分布于Ⅴ、Ⅳ垂直层,小径级木所占比例较高,物种自然更新良好,这与该自然保护区巴东栎和多脉青冈(Cyclobalanopsismultinervis)等物种的研究结果相一致[19];亮叶桦、盐肤木、白辛树和香椿等物种的个体多度随垂直层次降低而减少,这与东白山自然保护区马尾松(Pinusmassoniana)的研究结果相一致[16];川陕鹅耳枥、半边月和珙桐等物种的个体多度随垂直层次降低呈单峰曲线,即物种在中间高度层次上的个体多度多,这可能是因为中等径级的物种采取强烈保守的生活史策略,致使生长缓慢、死亡率低,从而在中等径级出现峰值[19-21].总体来看,随群落自然恢复年限增加优势种及主要伴生种的个体多度呈下降趋势,这可能与群落演替阶段等密切相关.越到群落演替早期,可利用的环境资源越丰富,一方面种内及种间竞争较弱,各种群能较好地共存于群落中;另一方面一些阳性落叶树种以R-生态对策可迅速占领群落较大生存空间[11].
4.1.2 径级结构 总体来看,3个恢复群落优势种及主要伴生种的物种丰富度均随径级增大而降低,超过20%的物种仅包含2个径级(Ⅰ、Ⅱ径级),超过60%的物种仅分布在前5个径级(DBH<30 cm),表明优势种及主要伴生种在径级分布上具有明显的间断现象,这与浙江天童山常绿阔叶林中的南酸枣(Choerospondiasaxiliaris)和栲树(Castanopsisfargesii)等物种的径级特征相似[22].
3个群落优势种及主要伴生种的个体多度总体上均呈倒“J”型分布,其结果与鼎湖山厚壳桂(Cryptocaryachinensis)、黄叶树(Xanthophyllumhainanense)等优势物种[18]、八大公山多脉青冈、亮叶水青冈(Faguslucida)等物种[21]及长白山红皮云杉(Piceakoraiensis)种群[23]相同.超过90%的个体集中分布在Ⅰ、Ⅱ径级,说明群落小径级木个体较多,种群自然更新良好.随自然恢复年限增加优势种及主要伴生种的个体多度在Ⅰ-Ⅲ径级范围内快速减少(图2、3、4),其原因可能是自然恢复年限越长群落可利用的环境资源越少,种群立木间对资源的利用性竞争加强,种群自疏和它疏效应增大,从而导致群落个体数量减少[11].自然恢复35a群落中,Ⅳ-Ⅵ径级的个体多度高于自然恢复20a和OG群落,这可能是因为该群落处于演替中期有较多常绿树种逐渐兴起,而一些早期阳生性较强且寿命较长的种群还未完全退出群落,导致群落混交度较高、个体多度较多.
4.2 动态趋势
4.2.1 种类组成 3个群落共有59种优势种及主要伴生种(表1),说明不同恢复群落优势种及主要伴生种的物种组成不尽相同,表明木林子自然保护区山地常绿落叶阔叶混交林群落结构复杂.一般来讲,群落结构越复杂其物种多样性就越高[24].本研究结果显示,自然恢复35a群落中DBH≥1 cm的木本植物有176种,高于浙江天童山20 hm2常绿阔叶林(152种)[22].整体而言,随自然恢复年限增加落叶树种的比例不断下降,而常绿树种的比例不断升高.
4.2.2 IV的动态趋势 3个群落中,排列前3的物种IV>1(表1),即共有6个物种的IV>1(青冈、川陕鹅耳枥、半边月、山矾、翅柃和尖连蕊茶).其中,青冈在3个群落中的IV都大,在OG群落中IVmax=2.36,说明青冈是山地常绿落叶阔叶混交林的建群种.
自然恢复20a~35a期间,除亮叶桦、锥栗、四照花、山矾和翅柃的IV增大以外,其他种群的IV均呈下降趋势;自然恢复35a-OG期间,青冈、翅柃、黄丹木姜子、锥栗、薄叶山矾、交让木、叶萼山矾和尖连蕊茶8个种群的IV增大,其他22个种群的IV降低.
4.2.3 优势种及主要伴生种动态趋势的综合分析 20a群落中,盐肤木、野漆、樱桃、棘茎楤木和望春玉兰5个种群的径级结构呈纺锤形或壶型锥体,属于典型的衰退型种群;半边月、灰柯、城口桤叶树、光叶水青冈和锥栗这5个种群呈部分间断的更新方式,属于进展与衰退相间出现的波动种群;其它20个种群的幼龄级个体多,呈连续更新的方式,属于进展种群.35a群落中,檫木、合欢、亮叶桦、大叶杨和盐肤木种群在Ⅰ径级(DBH<5 cm)的植株个体少于5株,说明种群林下更新不良,仅少数大径级的个体存活,这与猪血木(Euryodendronexcelsum)、裸果木(Gymnocarposprzewalskii)等一些珍稀濒危植物的径级结构相似[25-26],种群衰退趋势明显;川陕鹅耳枥、灯台树、枫香树、杉木、石灰花楸和锥栗这6个种群的植株个体在不同垂直层次和径级中均存在显著的波动,说明这些种群的进展期和衰退期相间出现;其它19个种群的个体数量在垂直结构和径级结构上都呈金字塔型,说明这些种群属于进展种群.OG处于群落演替后期,种群老龄个体所占比重较大.红柴枝、苦枥木、米心水青冈、香椿、锥栗和亮叶桦6个种群的个体数量均随垂直层次和径级变化呈现单峰曲线(DBH在35~100 cm径级范围内分布数量最多),其中亮叶桦和锥栗种群DBH≥30 cm的个体数量分布占种群个体总数的100%、82.35%,这说明群落老龄化发展趋势明显.珙桐幼年时期(10a生以前)喜欢较荫蔽的环境,当进入中龄期后,小树和中树对光的需求不断增加[27-28],由于部分大树和衰退种群的存在,随着群落演替进展,群落内必然会出现林窗,可为阳性树种更新及耐阴性物种快速生长提供可能性;此外,白辛树、青冈、巴东栎、灰柯、稠李、川陕鹅耳枥、粉椴和木荷8个种群与木林子保护区15 hm2大样地中白栎(Quercusfabri)、桦椴(Tiliachinensis)等物种一样,都属于进展与衰退相间出现的波动种群[19].
群落由种群构成,群落动态是所有种群动态的综合表现[19].木林子自然保护区3个恢复群落的山地常绿落叶阔叶混交林优势种及主要伴生种中超过50%的物种属于进展种群,少于16.67%的物种属于衰退种群,这体现了群落的进展状态.优势种及主要伴生种的物种丰富度和个体多度均随树高和径级降低而增加,表明多数物种的自然更新良好,群落整体上正处于演替进程中.

表2 不同恢复群落优势种及主要伴生种群的植物名录(按IV降序排列)Tab.2 Species of dominantand associatedpopulations in different recovery communities (According to the descending order of IV)
[1] JERNVALL J,FORTELIUS M.Maintenance of trophic structure in fossil mammal communities: site occupancy and taxon resilience[J].The American Naturalist,2004,164(5):614-623.
[2] 史作民,程瑞梅,刘世荣,等.宝天曼植物群落物种多样性研究[J].林业科学,2002,38(6):17-23.
[3] 陈伟,杨小波,李时兴,等.海南中部山区植被演变阶段植物物种多样性与群落结构多样性变化规律[J].热带作物学报,2014,35(4):784-790.
[4] LOREAU M,NAEEM S,INCHAUSTI P,et al.Biodiversity and ecosystem functioning: current knowledge and future challenges[J].Science,2001,5543(294):804-808.
[5] TILMAN D,REICH P B,KNOPS J M H.Biodiversity and ecosystem stability in a decade-long grassland experiment[J].Nature,2006,7093(441):629-632.
[6] 刘万德,臧润国,丁易.海南岛霸王岭两种典型热带季雨林群落特征[J].生态学报,2009,29(7):3465-3476.
[7] 宋厚娟,叶吉,师帅,等.长白山区阔叶红松林残留片段木本植物物种组成与群落结构[J].应用生态学报,2014,25(5):1239-1249.
[8] 周光裕.中国的落叶阔叶林:兼论常绿落叶阔叶混交林的归属问题[J].宁波大学学报,1996,9(3):93-102.
[9] 艾训儒,易永梅.清江流域植被研究[M].北京:中国农业科学技术出版社,2005.
[10] 姚兰,崔国发,易咏梅,等.湖北木林子保护区大样地的木本植物多样性[J].林业科学,2016,52(1):1-9.
[11] 汤景明,艾训儒,易永梅,等.鄂西南木林子常绿落叶阔叶混交林恢复过程中优势树种生态位动态[J].生态学报,2012,32(20):6334-6342.
[12] 张金屯.数量生态学[M].北京:科学出版社,2014.
[13] 祝燕,赵谷风,张俪文,等.古田山中亚热带常绿阔叶林动态监测样地:群落组成与结构[J].植物生态学报,2008,32(2):262-273.
[14] 丁晖,方炎明,杨青,等.武夷山中亚热带常绿阔叶林样地的群落特征[J].生物多样性,2015,23(4):479-492.
[15] 卜志国,于清芹,卢银平,等.燕山山脉天然次生油松林群落结构与物种多样性分析[J].西北农业学报,2014,23(4):99-104.
[16] 王云泉,田磊,仲磊,等.东白山自然保护区木荷-马尾松群落结构及物种多样性分析[J].浙江大学学报(理学版),2015,42(1):38-46.
[17] 黄忠良,孔国辉,叶万辉,等.鼎湖山南亚热带常绿阔叶林定位研究(Ⅱ)锥栗、黄果厚壳桂群落种群垂直结构与年龄结构及其动态特征.中国科学院鼎湖山森林生态系统定位研究站.热带亚热带森林生态系统研究[M].北京:中国环境科学出版社,1998:7-17.
[18] 叶万辉,曹洪麟,黄忠良,等.鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J].植物生态学报,2008,32(2):274-286.
[19] 姚兰.湖北木林子保护区15 hm2大样地森林群落结构及多样性[D].北京:北京林业大学,2016.
[20] YUE B,YE W,MULLER-LANDAU H C,et al.Unimodal tree size distributions possibly result from relatively strong conservatism in intermediate size classes[J].Plos One,2012,7(12):491-495.
[21] 卢志军,鲍大川,郭屹立,等.八大公山中亚热带山地常绿落叶阔叶混交林物种组成与结构[J].植物科学学报,2013,31(4):336-344.
[22] 杨庆松,马遵平,谢玉彬,等.浙江天童20 hm2常绿阔叶林动态监测样地的群落特征[J].生物多样性,2011,19(2):215-223.
[23] 李艳丽,杨华,亢新刚,等.长白山云冷杉种群结构和动态分析[J].北京林业大学学报,2014,36(3):18-25.
[24] REINERS W,WORLEY I,LAWRENCE D.Plant diversity in a chronosequence at Glacier Bay,Alaska[J].Ecology,1971,52(1):55-69.
[25] 申仕康,马海英,王跃华,等.濒危植物猪血木(Euryodendronexcelsum H.T.Chang) 自然种群结构及动态[J].生态学报,2008,28(5):2404-2412.
[26] 王立龙,王亮,张丽芳,等.不同生境下濒危植物裸果木种群结构及动态特征[J].植物生态学报,2015,39(10):980-989.
[27] WANG G,HAN S H,WANG H C,et.al.Living characteristics of rare and endangered species Davidia involucrate[J].Journal of Forestry Research,2004,15(1):39-44.
[28] 程芸.珙桐与光叶珙桐的生物学特性及群落结构研究[D].北京:北京林业大学,2008.
责任编辑:高 山
The Structure and Dynamics of Dominant and Associated Populationsin Mixed Evergreen and Deciduous Broad Leaved Forest in Mountainous Region of Different Recovery Communities
CHEN Si1,AI Xunru1*,YAO Lan1,2,LIN Yong1,HUANG Wei1,CHEN Jun1
(1.School of Forestry and Horticulture,Hubei University for Nationalities,Enshi 445000,China;2.School of Nature Conservation,Beijing Forestry University,Beijing 100083,China)
Mixed evergreen and deciduous broad-leaved forest is a typical natural vegetation type and natural forest type in China. In order to study the dominant and associated population structure and its dynamicst and reveal the mechanism of the formation and maintenance of biodiversity,we rely on the construction of National Forest Ecosystem Research Station of Hubei Enshi, to investigate the plant community of mixed evergreen and deciduous broad-leaved forest in different stages of natural recovery in Mulinzi Nature Reserve,and we analyzed the community characteristics of dominant and associated populations from the aspects of species composition, the areal-types, the diameter class structure and the vertical structure. The results show that: Among the dominant and associatedpopulations in different recovery communities, the species number of woody plants at breast height (DBH)≥1 cm is 148,176,149 and the plant number is 6 970,4 533,4 048.They belong to 23 genera of 14 families,26 genera of 19 families and 25 genera of 16 families.Overall, the species richness of the dominant species showed the ″unimodal type″ with the vertical layer drops in the 3 stages of natural recovery,the individual abundance increases with the decrease of the vertical layer.The average tree height is 5.94,5.21 and 4.61 m respectively. The species richness of different recovery communities decreases with the increase of the diameter class in different recovery communities,individual multi degree is inverted ″J″ type distribution. The number of small diameter class is large,the average tree DBH is 4.18,4.35 and 4.48 cm respectively. In short, the species composition of the forest communities is rich in the mixed evergreen and deciduous broad-leaved forest in different stages of natural recovery in Mulinzi Nature Reserve. Most of the population naturally updates well and the whole community is in a positive succession.
mixed evergreen and deciduous broad-leaved forest; Mulinzi Nature Reserve; natural recovery; vertical structure; size structure
2016-11-27.
清江流域退化生态系统植被恢复生态效应评价(D20122901).
陈斯(1990- ),女(土家族),硕士生,主要从事森林生态与生物多样性保护的研究;*
艾训儒(1967- ),男(土家族),博士,教授,主要重生森林生态学、农业生态学和环境生态学等的研究.
1008-8423(2016)04-0467-08
10.13501/j.cnki.42-1569/n.2016.12.023
S718.52
A
