甘薯细胞遗传学研究进展
2017-01-09李宗芸陈孚尧蒋姣姣俞立璇孙健英韩永华
李宗芸,陈孚尧,蒋姣姣,俞立璇,孙健英,韩永华,*
(1 江苏师范大学 生命科学学院,整合植物生物学研究所,徐州 221116;2 江苏师范大学 江苏省药用植物生物技术重点实验室,徐州 221116;3 中国矿业大学 环境与测绘学院,徐州 221116)
甘薯细胞遗传学研究进展
李宗芸1,2,陈孚尧1,3,蒋姣姣1,2,俞立璇1,2,孙健英1,2,韩永华1,2,*
(1 江苏师范大学 生命科学学院,整合植物生物学研究所,徐州 221116;2 江苏师范大学 江苏省药用植物生物技术重点实验室,徐州 221116;3 中国矿业大学 环境与测绘学院,徐州 221116)
指出了甘薯细胞遗传学研究仍存在的很多技术瓶颈,如功能基因定位等,总结了甘薯细胞遗传学的研究进展,包括染色体行为、杂交种染色体分析、甘薯起源与亲缘关系等. 同时展望了甘薯细胞遗传学的发展和亟待加强的方向,为甘薯遗传背景的研究提供了新的方法和思路.
甘薯;细胞遗传学;染色体
甘薯[Ipomoeabatatas(L.) Lam.],亦称番薯,旋花科(Convolvulaceae)番薯属(Ipomoea)植物,俗称地瓜、红薯、山芋、红苕等,原产美洲的秘鲁、墨西哥及厄瓜多尔一带[1],现在在世界粮食作物生产中总产量位列第五,由于其具有高产、稳产、抗逆性强等优点,被广泛种植于100多个国家.番薯属植物数量众多,经国内外专家的多年研究分类,将月光花属、笃萝属和牵牛属也并入番薯属,作为其中的亚属、组或亚组[2, 3].我国甘薯资源的保存数量虽远低于美国和日本,但保存量也达到2000余份,种类含20余种,且栽培面积和总产量仅次于水稻、小麦和玉米[4],其中番薯IpomoeaBatatas(L.) Lam.和蕹菜IpomoeaaquaticaForssk.为重要的粮食和蔬菜作物.为了弄清甘薯的起源与进化,各种近缘野生种也成为研究人员关注的重点,例如三浅裂野牵牛Ipomoeatrifida(H.B.K)Don、三裂叶薯IpomoeatrilobaL.、五爪金龙Ipomoeacairica(L.) Sweet、椴树野牵牛Ipomoeatiliacea(Willd.) Choisy等.
随着人们对食品营养与身体健康越来越重视,甘薯受到了极大的关注,因为它是世界上最营养密集的块根作物之一,含有促进健康的低分子量的抗氧化剂如维他命、花青素和类胡萝卜素[5],还是碳水化合物、食用纤维、糖类、蛋白质、铁和钙极好的来源[6].按照不同用途,甘薯可分为以下三种:一是食用型甘薯(包括菜用甘薯),其块根中的粗淀粉含量高达37.6%~77.8%,可溶性糖含量达1.68%~36.02%,粗蛋白含量达2.24%~12.21%;李爱贤等[7]2009年对广菜2号、台农71等菜用型甘薯的培育也是当前的一种新特优甘薯培育的方向.二是保健型甘薯,甘薯的营养成分如胡萝卜素、维生素B1、B2、C和铁、钙等矿物质的含量都高于大米和小麦粉.红薯中含有的大量花青素和甲基花青素类成分,能抑制杂环胺类致突变物质,还具有抗氧化、抗菌等作用,其中的糖蛋白还具有明显的降血脂及增强免疫力等功能[8].三是能源型甘薯,甘薯的生产量高且富含淀粉,是生产燃料乙醇的理想原料[9].近年来,国内外许多学者就甘薯的起源、生理生化特性、核型分析、基因功能等开展了多方面研究,但是研究大都缺乏系统性;由于甘薯基因组复杂,该物种的起源和进化等研究都远远落后于其他物种.本文全面回顾了甘薯染色体核型特点、杂交种染色体分析、起源与亲缘关系等研究进展,并对甘薯的细胞遗传学研究进行了展望.
1 甘薯染色体核型研究
甘薯具有营养繁殖的特点,且存在自交不亲和性、交配不亲和群[8],因此甘薯的育种背景非常复杂,也为其起源和系统进化分析带来了诸多困难.20世纪20~30年代,Heitz等[10]分析了甘薯属植物的染色体数目,确定了大多数物种的染色体基数为n=15; Nakajima[10]发现了n=14和n=24的非整倍体情况.
但染色体的基数研究仅是第一步,众多科学家对甘薯属的染色体进行了很多后续的研究.1963年,Goichi[11]利用乙酰-地衣紫色素涂片法对番薯属的6个物种(25个品系)的染色体核型进行了分析,发现大多数染色体均为曲棍状或者杆状,染色体之间形态差异较小,只能通过着丝粒的位置和随体的有无及数量对染色体进行大致区分,难以通过染色体核型判断品系间的亲缘关系.2012年,Rane A等[12]人分析了10种二倍体甘薯属植物粗线期染色体的核型,其中蕹菜I.aquaticaForsk.具有最小的染色体长度为1.25~2.67 μm,平均长度为(1.99±0.38) μm,单倍体合计长度为29.88 μm;而IpomoeacarneaJacq.具有最长的染色体长度为2.13~4.79 μm,平均长度为(3.39±0.74) μm,单倍体总长度50.83 μm.通过分析这些甘薯粗线期的核型,Rane A提出:一个对称核型的特点是中着丝粒或亚中着丝粒占主要.在核型的进化过程中,对称性的核型更加原始,而不对称的核型有更高的进化地位.通过着丝粒位置的移位增加染色体的不对称性,或通过互补染色体之间相对大小的差异的积累使更多的染色体核型发生变化.
近年来,汤佳立,安婷婷等[13,14]利用自身基因组荧光原位杂交和银染等技术对甘薯(包括栽培种和野生种)的染色体及核型进行了分析,发现徐薯18间期核有6对、8对和9对银染点;栽培种甘薯徐薯18 (IpomoeabatatasXushu18)核型公式为2n=6x=90=72m+18sm(9SAT),美国野生甘薯核型公式为2n=2x=30=30m(2SAT),香港野生甘薯核型公式为2n=2x=30=20m+10sm (2SAT);而野生种Ipomoeatrifida“698001” 、“698011”、 “P-875-6”核型公式分别为2n=2x=30=18m (2SAT)+12sm (2SAT); 2n=2x=30=17m(2SAT)+13sm (2SAT); 2n=6x=90=54m (2SAT) +36sm (2SAT).曹清河等[15]利用压片法和花粉粒扫描电镜法对甘薯近缘种裂叶牵牛IpomoeahederaceaJacq.的体细胞染色体数目、核型及其分类地位进行分析和界定,此近缘种的体细胞染色体数目为30,属于二倍体种,染色体长度为4.19~8.83μm,核型公式为2n=2x=30=26m (2SAT)+4sm (2SAT).
对于番薯属植物染色体上随体的研究,安婷婷等[14]发现:栽培种甘薯有9对随体,位于第1、3、6染色体上,2种近缘野生种(IpomoeahederaceaJacq.)有2对随体,均位于第6、12染色体上;曹清河等[15]的研究表明,产地为美国的近缘野生种(I.hederaceaJacq.)有2对随体;Srisuwan S等[16]的研究发现二倍体甘薯属物种中,I.trifida和I.trilobaL.有2对随体,I.setosaKer Gawler.只含有1对随体,而在IpomoealeucanthaJacq.和I.tiliacea(Willd.) Choisy.中有4对随体,这对于进一步判断甘薯属各物种的亲缘关系的远近提供了一定的理论依据.
2 杂交种染色体研究
甘薯种质资源非常丰富,据中国植物志记载大约300种(广义的约500种),其中包含众多的抗病、抗虫、抗逆性基因,科研及生产价值非常高,但非所有近缘野生种都能与甘薯进行杂交得到杂交种.1979年日本科学家寺村提出确定甘薯种间关系的主要依据是种间可交配性、杂种能育性和性状分离情况.根据种间的可交配性将甘薯组分为两类:杂交亲合群I和杂交不亲和群II,而杂交亲合群I资源有限.一直以来记载的能够与甘薯成功杂交并得到种间杂种的近缘野生种只有:三浅裂野牵牛Ipomoeatrifida(H.B.K)Don (2x,4x,6x)、三裂叶薯IpomoeatrilobaL.、五爪金龙Ipomoeacairica(L.) Sweet.、Ipomoeagrandifolia(Dammer) O′Donell.、野甘薯IpomoealacunosaL.、圆叶牵牛Ipomoeapurpurea(L.) Roth.[17]。
最早对甘薯近缘野生种进行杂交利用的是日本.20世纪70年代初,以Nishiyama为首的一批研究者利用K123与甘薯进行了杂交,并成功培育出含有I.trifida血统的新品种“南丰”,该品种具高产、高淀粉、高抗线虫病等多种优良性状[18],“南丰”的成功培育也成了史上近缘野生种种质资源利用的一个重要里程碑.随后Orjeda等[19]采用5个栽培种甘薯与41份二倍体I.trifida进行杂交,获得了248个杂种植株,大多数杂种植株为四倍体,并且在杂种植株中检测到了野生种的优良抗性基因.
近几十年,我国不少研究者也采用不同倍性的I.trifida与甘薯进行杂交获得优良性状的杂种品系,如河北农科院粮油所利用K123杂交培育出高干率品系冀Y1、冀Y25等[20],引进美国的I.trifida(6x)实生苗选系Y-2、Y-4、Y-6,培育出冀薯(98、99、71等)系列的几个新品种.我国的栽培种甘薯的育种背景比较狭窄,育成的甘薯品种近94%具有南瑞苕和胜利百号的血缘[21],应加强甘薯资源交换,充分利用外引亲本来拓宽国内甘薯的遗传背景(图1).

图1 栽培种甘薯“徐薯18” 系谱图(盛家廉等,1981)Fig.1 Family tree of cultivated sweet potato I. batatas “Xushu18” (Sheng Jialian et al, 1981)
1977年以来,江苏徐州甘薯研究中心先后从CIP (国际马铃薯中心)、美国路易斯安那州立大学、NPGS(美国国家种质资源库)引进了131份近缘野生种资源,加上国内收集的12份近缘种,共筛选出可长期保存的43份不同种或存在倍性差异的甘薯近缘野生种.曹清河等[17]通过将多种近缘野生种与甘薯进行配对杂交创制出了优良的“桥梁种”,它们具备了父本的抗性基因,同时根部也逐渐膨大化,可为下一步的回交转育提供优良基因.但由于甘薯染色体众多,形态大小相似,染色体之间差异难以辨识,对于杂交种的染色体分析较少.
3 甘薯起源与亲缘关系分析
从1955年开始,在墨西哥发现了大量的甘薯野生种和近缘种.通过形态学、细胞学和遗传学的研究发现I.leucantha(2n=30),I.littoralis(2n=60)和I.trifida(2n=90)这三个物种极有可能是甘薯Ipomoeabatatas(2n=90)的祖先种,这三个物种是同源多倍体,因为它们都有I.eucantha的B基因组加倍.利用I.leucantha(2n=30)和I.littoralis(2n=60)产生新的人工诱导形成的六倍体,用以研究甘薯的起源.人工诱导的六倍体I.leucantha生长情况较差且在减数分裂时期形成一些多价体,同时人工六倍体和I.batatas在形态和生理特性上存在差异.这些植物的地理分布表明栽培种甘薯可能起源于中美洲(墨西哥伊比利亚半岛和委内瑞拉奥里诺科河口之间),并由哥伦布于1492年第一次远洋后将其引入西欧[16],16世纪初又由葡萄牙探险家引入非洲、东南亚及东印度等地,随后被西班牙的水手带至菲律宾的马尼拉和摩鹿加岛,再传至亚洲各地[22].
而有关栽培种甘薯是由哪个原始物种进化而来以及如何进化这个问题,自20世纪初就有许多学者研究和争论,至今没有定论.美国学者认为栽培种甘薯是异源六倍体,源于I.trifida(2x)与I.triloba(2x)基因组的异源多倍化[23],不少学者采用多种二倍体和四倍体的野生种作为父本,与甘薯进行杂交均未成功;而以Nishiyama为代表的日本学者则认为甘薯的祖先种是野生种I.trifida(6x)的同源六倍体.I.trifida是包括二倍体、四倍体和六倍体等的复合体,广泛分布于热带美洲,在与甘薯杂交时显示了杂交亲和性.另外,由于不同品种甘薯杂交可分离出与六倍体野生种形态学上相似的类型,不少学者推测甘薯是I.trifida复合种的一个成员或者是派生出来的某个种[24].
关于甘薯的形成途径有3种推测:一是由二倍体野生种自然突变加倍形成四倍体野生种,四倍体和二倍体杂交形成三倍体,再由三倍体加倍形成了原始的六倍体野生种;二是二倍体野生种和四倍体野生种,在一定条件下形成了六倍体后被驯化为栽培型;三是二倍体发生突变导致其出现栽培化性状,随后染色体数加倍,使其栽培化性状更加显著,形成栽培型的四倍体及六倍体种[24](图2).未找到栽培型的二倍体或者四倍体,而上世纪Nishiyama等[25]公布在墨西哥采集到了六倍体的野生种 [K123,后定名为I.trifida(H. B. K) Don.];接着有研究者从墨西哥、哥伦比亚及委内瑞拉多地收集到一种可以与甘薯品种直接杂交的二倍体野生种K221,后归类为I.trifida(H. B. K) Don;又有研究者在哥伦比亚的高海拔地区发现了四倍体的野生型I.trifida,这些结果贯通了第一种推测途径.

注: ----系统发育过程中遗传与染色体的分化;人为染色体组加倍;Ⅰ、Ⅱ、Ⅲ分别表示形成6x的三种途径图2 甘薯及其祖先种自然和人工多倍体的系统发生(Nishiyama, 1975)Fig.2 Phylogeny of natural and artificial polyploids in I. Batatas and its progenitors. (Nishiyama, 1975)
2006年,Saranya S[16]利用FISH的方法对21种甘薯5S rDNA和18S rDNA进行分布和起源的研究,提出了甘薯起源的三种假说:第一种假说是野生二倍体I.trifida自动多倍化形成六倍体甘薯[I.batatas(L.) Lam.];第二种假说是野生二倍体I.trifida和野生二倍体I.triloba杂交形成的后代是甘薯野生祖先种,再在自然条件下加倍形成了六倍体甘薯;第三种假说是野生二倍体I.leucantha和野生四倍体I.littoralis杂交形成了某个三倍体的物种,这个三倍体在自然条件下加倍形成了六倍体甘薯(见图3).
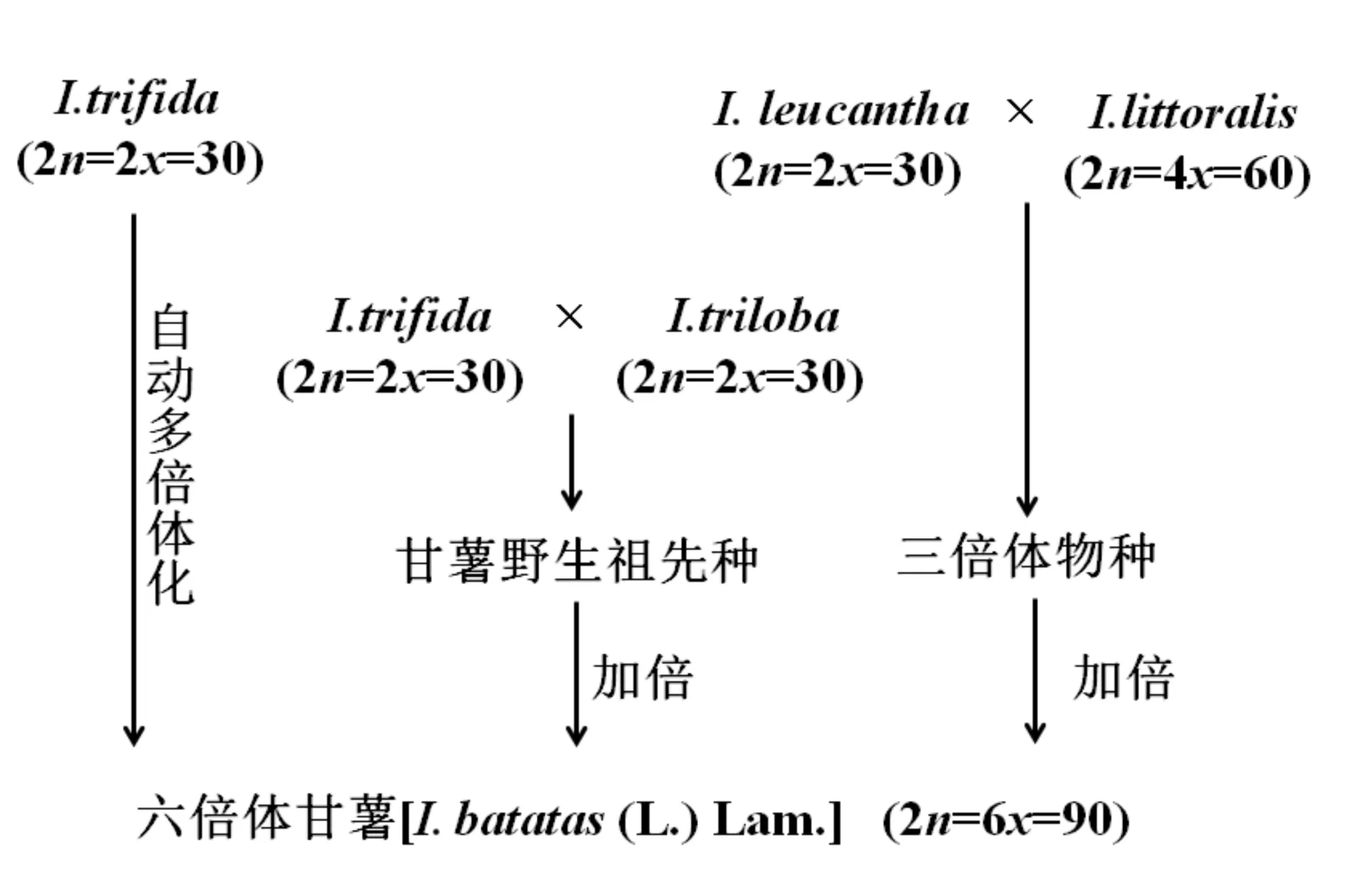
图3 甘薯的系统进化图Fig.3 Evolution of Ipomoea batatas(L.) Lam.
4 甘薯细胞遗传学研究展望
4.1 基于基因组测序的染色体识别
不同物种的染色体有各自相对稳定的特定的形态结构特征(包括染色体的长度、着丝点位置、臂比、随体大小等),因此,染色体核型分析是植物种质资源遗传性研究的重要内容. DNA序列的荧光原位杂交(FISH)信号可为染色体研究提供有效的细胞遗传学标记,如何群燕[26]利用FISH技术对萝卜染色体的两个重复序列CL1和CL25在染色体上的分布特征进行研究,发现CL1定位在染色体的着丝粒区域,CL25位于次端粒区,再将其与5S rDNA和45S rDNA结合同时对萝卜体细胞中期染色体进行分析,很好地识别萝卜染色体.
甘薯染色体数量众多,且染色体形态相近,单凭染色体的形态学特征难以识别全部的染色体.通过分析测序结果,筛选重复序列并以此作为探针,结合染色体的形态学特征准确识别染色体,由此建立精确的核型,是目前甘薯细胞遗传学发展的一个方向.2015年,Hirakawa H等[27]用自交型Mx23Hm和高度杂合型0431-1两个品系的Ipomoeatrifida(H. B. K.) Don.结合Illumina HiSeq平台,对全基因组序列进行分析和鉴定,而中美韩等国都在进行对栽培种甘薯的测序工作.本课题组俞立璇等[28]利用测序结果得到了二倍体I.trifida中的一些重复序列,并对这些序列进行了筛选,获得了两个重要重复序列Itf_1和Itf_2,并将Itf_1、Itf_2结合45S rDNA、5S rDNA对二倍体I.trifida染色体进行定位,对I.trifida染色体进行了有效识别.
4.2 分子细胞遗传图谱的构建
自1987年Donis-Keller等发表了第一张人类的RFLPs连锁图后,许多生物(包括植物)的RFLP图谱相继问世,由于植物能方便地建立和维持较大的分离群体,分子连锁图构建工作的发展速度超过了动物的同类研究,现已建图的植物已多达几十种,其中包括了所有重要的农作物[29].结合连锁图谱,在大规模的测序完成后,通过建立基因组文库,筛选克隆,可以进行分子细胞遗传图谱的构建.例如孙健英等[30]根据黄瓜基因组组装图谱筛选克隆,利用双色FISH技术在中期染色体上确定了相邻克隆的顺序,在此基础上,探针的混合物同着丝粒特有序列TypeIII杂交到粗线期染色体上,显示出间隔的红绿信号,成功构建了黄瓜7条染色体的分子细胞遗传学图谱.另外利用Oligo探针结合荧光原位杂交技术对染色体进行定位,也可以构建植物分子遗传图谱[31].
目前,甘薯中已经进行了多个分子遗传图谱的构建.2008年,Kriegner等[32]采用非洲甘薯品种Tanzania和美洲甘薯品种Beauregard杂交,建立了250个单株的F1作图群体,采用AFLP技术构建甘薯遗传连锁图谱,该图谱包括176个连锁群,图谱总长11078cM.2009年王红意等[33]以中国30个甘薯主栽品种为材料,对甘薯RAPD指纹图谱的构建进行了探讨,聚类分析表明:地域分布相近的甘薯品种和具有同一亲本的甘薯品种聚在一起,与这些甘薯品种的系谱图一致.2010年李爱贤等[34]用SRAP标记技术结合JoinMap3.0软件和“双假测交”策略,构建甘薯高密度分子连锁图谱.2015年,张安世等[35]用分子标记SRAP技术,构建22个甘薯品种的DNA指纹图谱并进行了遗传多样性分析.同时,甘薯基因组序列测定也在进行中,因此,甘薯分子细胞遗传学图谱的构建将成为甘薯细胞遗传学研究的重点.此外,比较基因组原位杂交等技术也是甘薯细胞遗传学研究中需要深入研究和改进的技术手段,这对于进一步明晰甘薯的系统发生、种间亲缘关系、起源与进化,指导甘薯育种有着积极的意义.
[1] 王永顺,姚应才. 江苏科学技术志[M].北京:科学技术文献出版社, 1997: 36-39.
[2] Austin D F. An infrageneric classification forIpomoea(Convolvulaceae) [J]. Taxon,1979, 28(4): 359-361.
[3] Austin D F. Additional comments on infrageneric taxonomy inIpomoea(Convolvulaceae) [J]. Taxon, 1980, 29(4): 501-502.
[4] 李 俊,王章英,罗忠霞,等.分子生物学技术在甘薯育种中的应用[J].广东农业科学, 2011, 38 (15): 108-112.
[5] Teow C C, Truong V D, McFeeters R F, et al. Antioxidant activities, phenolic and b-carotene contents of sweetpotato genotypes with varying flesh colours[J]. Food Chem, 2007, 103(3): 829-838.
[6] Chen P N, Chu S C, Chiou H L, et al. Cyanidin 3-glucoside and peonidin 3-glucoside inhibit tumor cell growth and induce apoptosisinvitroand suppress tumor growthinvivo[J]. Nutr Cancer, 2005, 53(2): 232-243.
[7] 李爱贤,刘庆昌,王庆美,等.我国甘薯育种研究现状及展望[J].山东农业科学, 2009(1): 38-42.
[8] 曹清河,刘义峰,李 强,等. 菜用甘薯国内外研究现状及展望[J].中国蔬菜, 2007(10): 41-43.
[9] Wang M, Shi Y, Xia X, et al. Life-cycle energy efficiency and environmental impacts of bioethanol production from sweet potato[J]. Bioresour Technol, 2013, 133(4): 285-292.
[10] Nakajima G. Chromosome numbers in some crops and wild angiosperms [J]. Jap J Genet, 1936, 12: 211-218.
[11] Nakajima G. Karyotype of genusIpomoea[J]. Cytologia, 1963, 28: 351-359.
[12] Rane V A, Patel B B, George J . Karyotype analysis of ten species ofIpomoeaJacq[J]. Cytologia, 2012, 77 (2): 239-249.
[13] 汤佳立,戚大石,张 俞,等. 荧光原位杂交技术分析栽培种甘薯(Ipomoeabatatascv.Xushu No.18)染色体[J]. 遗传, 2010, 32 (2): 177-182.
[14] 安婷婷,汤佳立,李宗芸,等. 甘薯栽培种及其近缘野生种的DAPI核型及rDNA-FISH分析[J]. 西北植物学报, 2012, 32 (4): 682-687.
[15] 曹清河,张 安,马代夫.甘薯近缘种染色体核型及花粉粒超微结构分析[J].西北植物学报, 2008, 28 (10): 1610-1613.
[16] Srisuwan S, Sihachakr D, Sonja S Y. The origin and evolution of sweet potato (IpomoeabatatasLam.) and its wild relatives through the cytogenetic approaches[J]. Plant Sci, 2006, 171 (3): 424-433.
[17] 曹清河,张 安,李 鹏,等.甘薯近缘野生种的抗病性鉴定与新型种间杂种的获得[J].植物遗传资源学报, 2009, 10 (2): 224-229.
[18] 沈稼青.甘薯近缘植物三浅裂野牵牛(I.trifida)杂交不亲和性的研究[J].作物学报, 1982, 8 (1): 9-13.
[19] Orjeda G, Freyre R, Iwanaga M. Use ofIpomoeatrifidagerm plasm for sweet potato improvement. 3. Development of 4xinterspecific hybrids betweenIpomoeabatatas(L.) Lam. (2n=6x=90) andI.trifida(H. B. K) G. Don. (2n=2x=30) as storage-root initiators for wild species[J]. Theor Appl Genet, 1991, 83(2): 159-163.
[20] 张松树, 刘兰服.甘薯野生种质资源在甘薯杂交育种中的利用[C]//中国作物学会.全国甘薯育种与产业化学术研讨会论文集.成都:中国作物学会,2005: 29-32.
[21] 胡 玲, 李 强, 王 欣, 等.甘薯地方品种和育成品种的遗传多样性[J].江苏农业学报, 2010, 26 (5): 925-935.
[22] Austin D F.Ipomoealittoralis(Convolvulaceae) - taxonomy, distribution, and ethnobotany[J]. Economic Botany, 1991, 45 (2): 251-256.
[23] Magoon M L, Krishnan R, Vijaya Bai K. Cytological evidence on the origin of sweet photo[J]. Theor Appl Genet, 1970, 40(8): 360-366.
[24] 董玉琛, 郑殿升.中国作物及其野生近缘植物:粮食作物卷[M].北京:中国农业出版社, 2006: 490-493.
[25] Nishiyama I, Miyazaki T, Sakarnoto S. Evolutionary autoploidy in the sweet potato (Ipomoeabatatas(L.) Lam.) and its Progenitors[J]. Euphytica, 1975, 24(1): 197-208.
[26] He Q, Cai Z, Hu T, et al. Repetitive sequence analysis and karyotyping reveals centromere-associated DNA sequences in radish (RaphanussativusL.) [J]. Bmc Plant Biology, 2015, 15(1): 1-12.
[27] Hirakawa H , Okada Y, Tabuchi H, et al. Survey of genome sequences in a wild sweet potato,Ipomoeatrifida(H. B. K.) G. Don [J]. DNA Res, 2015, 22 (2): 171-179.
[28] 俞立璇.重复序列Itf_1, Itf_2在甘薯核型分析中的应用与甘薯及其近缘野生种ITS序列鉴定[D].徐州:江苏师范大学, 2014.
[29] 张德水, 陈受宜.DNA分子标记、基因组作图及其在植物遗传育种上的应用[J]. 生物技术通报, 1985(5): 15-22.
[30]SunJ,ZhangZ,ZongX,etal.Ahigh-resolutioncucumbercytogeneticmapintegratedwiththegenomeassembly[J].BMCGenomics, 2013, 461: 461-485.
[31]HanY,ZhangT,JiangJ,etal.Chromosome-specificpaintingincucumisspeciesusingbulkedoligonucleotides[J].Genetics, 2015, 200 (3): 771-779.
[32] 梁雪莲, 罗忠霞. 甘薯分子遗传图谱研究进展与展望[J]. 广东农业科学, 2014, 41(3): 145-148.
[33] 王红意, 翟 红, 王玉萍. 30个中国甘薯主栽品种的RAPD指纹图谱构建及遗传变异分析[J].分子植物育种, 2009, 7(5) :879-884.
[34] 李爱贤, 刘庆昌, 王庆美. 利用SRAP标记构建甘薯分子连锁图谱[J].作物学报, 2010, 36 (8): 1286 -1295.
[35] 张安世, 张利民, 徐九文.甘薯SRAP指纹图谱构建及遗传多样性分析[J].生物技术, 2015, 25 (2): 161-164.
Cytogenetics Research Progress of Sweet Potato
Li Zongyun1,2, Chen Fuyao1,3, Jiang Jiaojiao1,2, Yu Lixuan1,2, Sun Jianying1,2, Han Yonghua1,2
(1 Institute of Integrative Plant Biology, School of Life Science, Jiangsu Normal University, Xuzhou 221116, China; 2 Key Laboratory of Biotechnology for Medicinal Plants of Jiangsu Province, Jiangsu Normal University, Xuzhou 221116, China; 3 School of Environment Science and Spatial Informatics, China University of Mining and Technology, Xuzhou 221116, China)
It is pointed out that there are still a great many technical bottlenecks in the cytogenetics of sweetpotato, such as functional gene mapping, etc. So the research progress of cytogenetics of sweet potato were summarized, including chromosome behavior, hybrid chromosome analysis,sweet potato origin and genetic relationship. Moreover, the development of sweetpotato cytogenetics and the direction to be strengthened were prospected, providing new methods and ideas for the study of sweet potato genetic background.
sweet potato; cytogenetics; chromosome
2016-09-07 *通讯作者 韩永华(1971-),女,教授, 研究方向:植物遗传学研究, E-mail:hanyonghua@jsnu.edu.cn
李宗芸(1964 -),女, 教授, 博士生导师, 研究方向:植物遗传学研究, E-mail: zongyunli@jsnu.edu.cn
国家自然科学基金资助项目(31271698);江苏省自然科学基金基础研究计划资助项目(BK2012579);江苏省高校自然科学研究重大资助项目(12KJA180001);徐州市科技计划资助项目(XF13C056) ,江苏省高校优势学科建设项目(PAPD)
Q343
A
1672-4321(2016)04-0034-06
