玉米生长苗期与穗期对土壤外源镉的响应
2017-01-05祁剑英杜天庆郝建平薛建福杨珍平崔振魁陈梦妮
祁剑英,杜天庆,郝建平,薛建福,杨珍平,崔振魁,陈梦妮
(山西农业大学农学院,山西太谷030801)
玉米生长苗期与穗期对土壤外源镉的响应
祁剑英,杜天庆,郝建平,薛建福,杨珍平,崔振魁,陈梦妮
(山西农业大学农学院,山西太谷030801)
采用盆栽试验,研究了不同质量分数的镉胁迫对大丰30玉米苗期与穗期细胞膜透性、丙二醛含量、过氧化物酶活性、叶片枯黄比例以及干物质量的影响,并分析了大丰30玉米生长苗期与穗期对镉胁迫的抗性差异。结果表明,土壤外源镉对玉米生长的抑制作用主要发生在穗期(50~70 d),并在中后期(70~90 d)有所减弱;在玉米生长穗期,当外源镉为1 mg/kg时,过氧化物酶(POD)活性显著低于对照(P<0.05),而其他指标没有显著变化,对玉米造成影响的土壤外源镉临界值小于1 mg/kg;外源镉质量分数与丙二醛(MDA)含量和细胞膜透性均呈二次曲线回归模型,但高质量分数镉胁迫下细胞膜透性变化量大于MDA含量;苗期7 mg/kg外源镉胁迫下叶片的枯黄比例是对照的3.1倍,且显著高于对照(P<0.05)。研究结果可为进一步了解镉胁迫对玉米不同生育时期的影响机制以及丰富玉米重金属污染胁迫研究提供理论依据。
外源镉;玉米;生理响应;生育时期
“十三五”规划重点强调粮食安全生产,而目前我国耕地土壤受重金属污染高达16.67%,其中,山西省耕地受其污染尤为严重[1],对粮食生产安全造成了严重的隐患。目前研究重金属污染土壤的修复问题日益引起国内外专家学者的关注。植物修复技术是重金属污染土壤修复方法之一,但植株在富集重金属的同时其不同器官也会不同程度地受到重金属的胁迫,具体表现为植株光合系统受到影响[2],遭受氧化胁迫和膜损伤[3],活性氧代谢的有关酶系(如抗氧化酶系的活性)受到改变[4-5],从而植物生长发育受到抑制,进而影响植物修复重金属污染土壤。因此,研究重金属胁迫下植物体的生理生化响应是提高植物对重金属毒害耐性的关键。
玉米生长速度快,生物量大,对于土壤中多种重金属具有一定的富集作用[6],可被用于修复重金属污染土壤。全国镉无机污染物点位超标率约为7.0%[7],是最主要的重金属污染之一。镉易被作物吸收[8],但可明显影响作物生理过程。目前,关于玉米对镉胁迫的生理生化响应研究较多,但针对不同生育时期玉米的耐受性研究相对罕见。
本试验以山西主推大丰30玉米品种为供试材料,通过土壤盆栽试验,分析玉米生长苗期与穗期对土壤中不同浓度外源镉胁迫的响应,比较苗期与穗期玉米对土壤外源镉胁迫的抗性差异,并探讨不同生理指标对镉浓度变化的敏感性,以期进一步了解玉米不同生育时期对土壤外源镉的耐受能力,为植物修复重金属污染土壤提供理论依据。
1 材料和方法
1.1 试验设计
试验于2015年5—10月在山西农业大学农作站进行。研究采用盆栽方法,盆内径30cm、高38cm,盆栽用土取自山西农业大学试验田0~20 cm表层土壤,其基本肥力为有机质19.23 g/kg,全氮1.012 g/kg,速效氮46.75 mg/kg,速效磷10.95 mg/kg,速效钾208.6mg/kg,土壤Cd含量本底值为0.54mg/kg。土壤经自然风干,过5 mm筛,以复合肥(N∶P2O5∶K2O=20∶10∶18)为肥源,按比例与土壤混匀并定量分装在盆里,每盆装土12.5 kg。
本试验中不同浓度的镉溶液采用CdCl2· 2.5H2O(分析纯)配制,试验设4种镉质量分数处理(在重金属污染中,轻中度污染是主要污染[1],因此,质量分数设置以国家3级标准为参考),分别为1(C1),3(C2),5(C3),7 mg/kg(C4),并设计空白对照(CK),每个处理15次重复,其中,12次重复用于冠部干物质量测定,3次重复用于生理指标测定。将配制的不同质量分数镉溶液1 000 mL在播种前均匀地渗入各个盆栽土壤中,于土壤平衡14 d后(5月1日)播种,供试玉米品种为山西省主推品种大丰30,每盆播种20粒种子,出苗后3~5叶间苗,每盆留苗5株,胁迫期间进行正常管理,定期定量浇水。
1.2 测定项目及方法
将玉米生长分为4个阶段:第1阶段为播种后0~30 d(苗期),第2阶段为30~50 d(苗期—穗期),第3阶段为50~70 d(穗期),第4阶段为70~90 d(穗期—花粒期)。由于本试验是盆栽试验,所提供土壤空间不足以支持玉米花粒期生长,因此,花粒期各项指标未做测量。
冠部干物质量测定:从玉米苗期(播种后30 d)开始测量冠部干物质量,以后每隔20 d测定一次,共测4次。每次每个处理取样3个重复,用清水洗净,于烘箱105℃杀青,再于75℃烘干,用分析天平称质量。不同外源Cd胁迫下干质量增长量数据统一转换为抑制率,抑制率=(1-处理值/对照值)×100%[9]。
生理指标测定:分别于玉米苗期(播种后30 d)和穗期(播种后70 d)取新鲜倒3片功能叶进行各项生理指标测定。POD活性采用愈创木酚法,以每分钟内A470变化0.01为一个酶活性单位,单位为U/(min·g)[10];MDA含量用TBA(硫代巴比妥酸)法测定,单位为μmol/g[11];细胞膜透性用相对电导率(%)表示;电导率用DS-11电导仪测定。
叶片枯黄比例统计:枯萎叶数以叶片2/3枯黄为标准。于苗期和穗期统计植株的枯萎叶数和展开叶数,并计算叶片枯黄比例。叶片枯黄比例=枯黄叶数/完全展开叶数[12]。
1.3 数据处理
试验数据采用Excel进行整理计算,并用SPSS 19.0数据分析软件对不同指标数据进行方差分析和Duncan法多重比较,文中作图用Sigmaplot 12.0进行。
2 结果与分析
2.1 土壤外源镉对玉米冠部干物质积累量的影响
由图1可知,C1镉胁迫下,第1阶段和第2阶段抑制率均为负值,表明低质量分数镉能够促进玉米生长前期地上部分干质量增长,且第2阶段促进作用略高于第1阶段。C4处理下,第2阶段的抑制率为负,但与第1阶段差异并不显著。不同镉质量分数下,第3阶段的抑制率最高,均显著高于其他阶段(除C1外)。第4阶段的抑制率大部分为负值(C1除外),表明镉对玉米的主要抑制作用在第4阶段前。
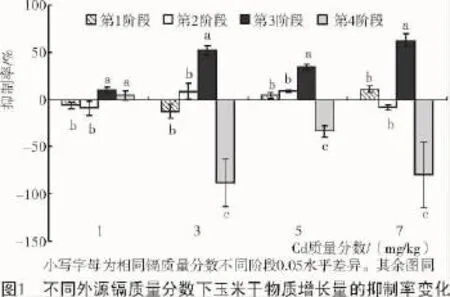
2.2玉米生长苗期、穗期不同指标对外源镉的响应
图1显示,外源镉对玉米穗期干物质增长量的抑制率均大于苗期,为探讨其原因,分析了玉米苗期和穗期不同质量分数外源镉胁迫对植株不同指标的影响。
由图2可知,在玉米生长苗期,叶片的细胞膜透性随镉质量分数的升高而增大,与对照相比,其发生显著变化时的镉质量分数为5 mg/kg。在穗期,当外源镉质量分数≤5 mg/kg时,叶片细胞膜透性没有发生显著性变化,C4处理即镉胁迫达到7 mg/kg时,细胞膜透性显著高于对照以及其他质量分数处理。
随着镉质量分数的增加,苗期MDA含量总体呈逐渐上升的趋势;除C1处理外,其他处理MDA含量均显著高于对照。穗期MDA含量则随镉质量分数增加呈先升高后降低的变化趋势,C3和C4处理显著低于对照。不同质量分数处理下,MDA含量均表现为穗期>苗期。
苗期POD随镉质量分数的升高而增大,且与对照相比,其开始发生显著变化是在5 mg/kg;而穗期则呈逐渐降低趋势,各质量分数处理均显著低于对照。在镉胁迫下,植株POD活性表现为苗期>穗期,而对照处理则表现相反。
在不同质量分数镉胁迫下,玉米苗期的干物质积累量呈先增加后降低的趋势,C4处理显著低于对照;穗期的干物质积累量则呈明显的降低趋势,对照显著高于C2,C3和C4处理,表明穗期干物质积累量对镉胁迫的响应较苗期更敏感。
综上所述,随着镉质量分数的增加,玉米苗期MDA含量发生显著变化的镉质量分数为3 mg/kg,POD活性和细胞膜透性发生显著变化的镉质量分数为5 mg/kg,干物质积累为7 mg/kg,所以,苗期不同指标的敏感性顺序为MDA含量>细胞膜透性=POD活性>干物质积累量。穗期,POD活性比其他指标敏感,在1 mg/kg的镉胁迫下,POD活性就表现出了明显的下降趋势,说明玉米穗期抵御镉胁迫对其所造成的自由基伤害的能力大大下降。另外,穗期干物质积累量在C2处理显著低于对照,表明玉米穗期受镉抑制大于苗期。

2.3 外源镉质量分数与细胞膜透性和MDA含量的回归分析
由2.2可知,苗期MDA含量与细胞膜透性变化趋势不完全一致,为进一步探究二者之间的关系,对不同质量分数镉胁迫与细胞膜透性和MDA含量进行了回归分析(图3)。图3表明,随着镉胁迫质量分数的增加,细胞膜透性、MDA含量均呈二次曲线变化,二者都呈上升趋势,但在高质量分数镉胁迫下(5~7 mg/kg),细胞膜透性的变化率逐渐增加,而MDA的变化率逐渐减小,表明此浓度范围内,细胞膜透性变化量大于MDA的变化量。

2.4 外源镉对玉米苗期与穗期叶片枯黄比例的影响
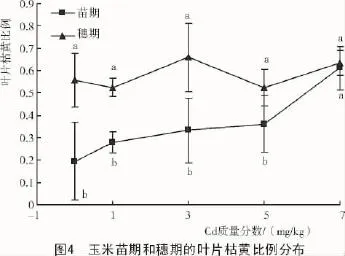
由图4可知,在C1,C2,C3和C4处理下,玉米苗期叶片的枯黄比例分别为对照的1.4,1.7,1.9和3.1倍,且C4处理下显著高于对照。而在玉米穗期,叶片的枯黄比例随镉质量分数的增加没有显著变化,但穗期的叶片枯黄比例均高于苗期。
3 讨论与结论
3.1 外源镉对不同阶段玉米生长的影响
不同外源镉质量分数下,第3阶段抑制率显著高于其他阶段(除1 mg/kg镉质量分数外),表明镉对玉米生长的抑制作用主要发生在第50~70 d,该阶段是玉米生长的穗期,穗期包括拔节期。有研究表明,玉米生长苗期比拔节期抗旱[13],遭遇涝害时,苗期涝害最大减产54.97%,拔节期涝害最大减产100%[14],苗期抗涝害能力大于拔节期。因此,该阶段的抑制率最高可能是由于拔节期抗逆能力小于苗期。本研究表明,不同质量分数外源镉对穗期的抑制率大于苗期,然而镉对玉米苗期与穗期的毒害机制有什么不同,导致镉在穗期抑制率显著高于苗期的原因是什么,这些问题还有待研究。
轻、中度镉胁迫促进生长,重、高度镉胁迫抑制生长[15]。但有研究认为,镉对水稻植株的毒害作用随生育进程而减弱,即镉对地上部干物质生产的毒害作用生育后期相对较轻[16]。本试验结果表明,在玉米生长第4阶段(穗期—花粒期),玉米地上部分干物质增长量抑制率为负值(C1处理除外),且显著低于其他阶段,表明该时期内不同质量分数外源镉对玉米地上部分干物质增长量均有促进作用,原因可能是由于玉米生长最快的时期在第3~4阶段,镉推迟了该最快生长时期,而造成镉的毒害作用在生育后期比较轻[16],因此,在第4阶段内,镉处理下的玉米干物质增长量大于对照。
3.2 苗期、穗期不同指标敏感性及外源镉毒害临界值分析
植物对重金属的响应有先后顺序,首先是避逆性,其次是耐逆性,在耐逆过程中,重金属被细胞壁固定以及被液泡区室化,进而细胞合成渗透调节物质脯氨酸等,最后,如果以上机制不能有效阻止重金属,抗氧化系统会开启[17]。Dalvi等[18]认为,当重金属试图进入根细胞时,首先是被细胞壁固定,产生有机酸、植物螯合物等,如果仍有过量的重金属存在,抗氧化系统开启。因此,随重金属质量分数的增加,不同的生理指标的变化是有先后顺序的。任安芝等[19]研究表明,在不同质量分数的重金属(镉、铬、铅)处理下,青菜中脯氨酸含量的变化先于细胞膜透性的变化,这与上述内容一致。本研究结果表明,随着重金属镉质量分数的增加,生理指标的变化先于生长指标的变化,这与Lin等[20]的研究结果相一致,另外,不论在苗期还是穗期,POD活性随外源镉质量分数变化均是敏感的指标之一,苗期MDA含量也比较敏感,因此,抗氧化酶活性和MDA含量可以对土壤镉污染起到早期指示作用[21]。然而,随重金属质量分数变化,其他不同指标的变化有没有先后顺序,有怎样的先后顺序,为什么会有这样的先后顺序,用怎样的试验方法可以研究这种顺序,这些问题有待于研究。
同时,研究不同指标随重金属质量分数变化的先后顺序可以确定毒害临界值,这与丁枫华等[22]对响应端点的研究观点相一致。综合不同指标可知,对苗期玉米造成影响的土壤外源镉临界值为3 mg/kg,即当土壤中外源镉达到3 mg/kg时就会对玉米生长造成一定的影响。以穗期指标为参考,POD活性先于其他指标显著变化,发生显著变化的镉质量分数为1 mg/kg,因此,在穗期外源镉对玉米产生毒害的临界值为1 mg/kg,该结果与国家土壤环境质量标准中镉的3级标准一致。可见玉米在生长穗期对土壤外源镉更敏感,造成这种结果的原因可能是:(1)增加胁迫时间可以增加重金属镉的毒害作用[19];(2)不同生育时期生理指标有不同的变化模式[11];(3)如3.1所述,镉在玉米穗期的抑制率最高。当土壤中外源镉质量分数小于临界值而未产生明显的生理指标变化,原因可能是:(1)该质量分数下重金属大部分被细胞壁阻拦[17];(2)菌根真菌[23]以及根分泌物对镉胁迫有缓解作用[18,24]。有研究依据规定将植物生物量或产量减少5%~10%土壤有害物质的质量分数作为土壤有害物质的最大允许质量分数,以灯心草为研究对象,确定土壤镉临界值为10 mg/kg[25],该试验研究的实际上是“最小允许度”。Lin等[20]研究表明,以抗氧化酶系统作为评价指标,土壤中外源镉对小麦产生毒性的临界值在3.3~10 mg/kg,与本试验结果接近,造成差异的原因可能是所研究作物不同[26]。
3.3 镉胁迫对MDA含量与细胞膜透性的影响
MDA是膜脂过氧化的产物,其含量可反映膜脂过氧化的程度,是植物膜系统受伤害的重要指标之一[27]。细胞膜透性变化是反映质膜破坏程度的重要指标。重金属胁迫下膜脂过氧化程度与细胞膜透性的变化有关[28],即MDA含量的增加,说明膜脂过氧化加剧,表现为膜透性增加[29]。镉胁迫可以使植物体内活性氧积累,引发生物膜脂过氧化,从而破坏膜系统的功能和结构[3],此外,随着镉质量分数的增加可以引起细胞膜透性增大[19]。研究表明,MDA含量在体内的变化规律与膜透性变化规律一致[30]。亦有研究表明,用MDA含量评价细胞受氧化损伤有局限性[31],即MDA含量与细胞受到氧化损伤的大小不是完全吻合的。本试验结果表明,MDA含量与细胞膜透性的变化规律也不完全吻合,尽管其均随镉质量分数的增加而增加,但在高质量分数镉胁迫下,MDA含量变化率小于细胞膜透性变化率,说明在高质量分数镉胁迫下,除了膜脂过氧化,还有其他因素影响细胞膜透性。这可能是由于重金属影响细胞膜上蛋白质的合成[32],或MDA本身能够加剧细胞膜损伤所致。
另外,黄玉山等[33]研究认为,MDA和干物质积累这2个内在指标和形态指标的变化在镉的作用下是一致的,本研究中,MDA与干物质积累量这2个指标并不完全一致,而是MDA变化先于干物质积累量变化(苗期),造成这种差异的原因可能是试验条件不同(本试验采用的是盆栽,与该试验的水培法不同),也可能是镉的质量分数设置不同。
3.4 镉胁迫对叶片枯黄比例的影响
在玉米生长苗期,叶片枯黄比例在C4处理下显著高于对照,表明叶片枯黄比例可以作为评价重金属毒害的指标,造成枯萎叶提早产生的原因可能是由于镉阻碍了某些营养元素[3,34-35]和水分的吸收[32]。外源镉能够抑制原叶绿素酯氧化酶的活性和影响氨基-γ-戊酮酸的合成,进而导致叶绿素含量下降[12],引起叶片枯黄。但穗期叶片枯黄比例变化不显著,表明用叶片枯黄比例评价重金属的毒害作用有局限性。
综上所述,外源镉对玉米的抑制作用主要发生在玉米生长第50~70 d(穗期),而此后(70~90 d)毒害作用有所减轻。在玉米生长穗期,随镉质量分数的增加,POD活性较其他指标更为敏感,因此,对玉米造成伤害的外源镉临界值小于1 mg/kg。在高质量分数镉胁迫下(5~7 mg/kg),膜脂过氧化不是影响细胞膜透性的主要因素。苗期叶片枯黄比例可以作为评价重金属毒害的指标。以上结果可为进一步了解镉胁迫对玉米不同生育时期的影响机制以及丰富玉米重金属污染胁迫研究提供理论依据。
[1]宋伟,陈百明,刘琳.中国耕地土壤重金属污染概况[J].水土保持研究,2013,20(2):293-298.
[2]Greger M,Ögren A.Direct and indirect effects of Cd2+on photosynthesis in sugar beet(Beta vulgaris)[J].Physiologia Plantarum,1991,83(1):129-135.
[3]宇克莉,邹婧.镉胁迫对玉米幼苗抗氧化酶系统及矿质元素吸收的影响[J].农业环境科学学报,2010,29(6):1050-1056.
[4]ShawB.Effects of mercury and cadmium on the activities of antioxidative enzymes in the seedlings of Phaseolus aureus[J].Biologia Plantarum,1995,37(4):587-596.
[5]王兴明,涂俊芳,李晶,等.镉处理对油菜生长和抗氧化酶系统的影响[J].应用生态学报,2006,17(1):102-106.
[6]赵芸晨,秦嘉海.长期定点施肥对制种玉米土壤理化性状及重金属含量的影响[J].水土保持学报,2012,26(6):204-208.
[7]中华人民共和国环境保护部.全国土壤污染状况调查公报[EB/OL].[2014-04-17].http://www.zhb.gov.cn/gkml/hbb/qt/201404/t20140417_270670.htm.
[8]Daud M,Ali S,Variath M,et al.Differential physiological,ultramor-phological and metabolic responses of cotton cultivars under cadmiumstress[J].Chemosphere,2013,93(10):2593-2602.
[9]杜天庆,杨锦忠,郝建平,等.小麦不同生育时期Cd、Cr、Pb污染监测指标体系[J].生态学报,2010,30(7):1845-1852.
[10]张志良.植物生理学实验指导[M].北京:高等教育出版社,1990:154-155.
[11]廖柏寒,刘俊,周航,等.Cd胁迫对大豆各发育阶段生长及生理指标的影响[J].中国环境科学,2010,30(11):1516-1521.
[12]刘俊祥,孙振元,巨关升,等.结缕草对重金属镉的生理响应[J].生态学报,2011,31(20):6149-6156.
[13]白向历,孙世贤,杨国航,等.不同生育时期水分胁迫对玉米产量及生长发育的影响[J].玉米科学,2009,17(2):60-63.
[14]刘祖贵,刘战东,肖俊夫,等.苗期与拔节期淹涝抑制夏玉米生长发育、降低产量[J].农业工程学报,2013,29(5):44-52.
[15]高芳,林英杰,张佳蕾,等.镉胁迫对花生生理特性、产量和品质的影响[J].作物学报,2011,37(12):2269-2276.
[16]程旺大,姚海根,张国平,等.镉胁迫对水稻生长和营养代谢的影响[J].中国农业科学,2005,38(3):528-537.
[17]Emamverdian A,Ding Y,Mokhberdoran F,et al.Heavy metal stress and some mechanisms of plant defense response[J].The Scientific World Journal,2015(1):1-18.
[18]Dalvi A,Bhalerao S.Response of plants towards heavy metal[J]. Annals of Plant Sciences,2013,2:362-368.
[19]任安芝,高玉葆,刘爽.铬、镉、铅胁迫对青菜叶片几种生理生化指标的影响[J].应用与环境生物学报,2000,6(2):112-116.
[20]Lin R,Wang X,Luo Y,et al.Effects of soil cadmium on growth,oxidative stress and antioxidant system in wheat seedlings(Triticum aestivum L.)[J].Chemosphere,2007,69(1):89-98.
[21]张琳,崔红米,王建军,等.镉胁迫对不结球白菜Vc合成L-半乳糖途径基因表达及抗氧化系统的影响[J].植物生理学报,2015,51(7):1099-1108.
[22]丁枫华,刘术新,罗丹,等.23种常见作物对镉毒害的敏感性差异[J].环境科学,2011,32(1):277-283.
[23]田野,张会慧,孟祥英,等.镉(Cd)污染土壤接种丛枝菌根真菌(Glomus mosseae)对黑麦草生长和光合的影响[J].草地学报,2013,21(1):135-141.
[24]张玲,王焕校.镉胁迫下小麦根系分泌物的变化[J].生态学报,2002,22(4):496-502.
[25]孙健,铁柏清,钱湛,等.单一重金属胁迫对灯心草生长及生理生化指标的影响[J].土壤通报,2007,38(1):121-126.
[26]Benavides M,Gallego S,Tomaro M.Cadmium toxicity in plants[J]. Toxic Metals in Plants,2005,17(1):21-34.
[27]Souza J.Maize and radish sequester excess cadmium and zinc in different ways[J].Plant Science,2003,165(5):1009-1022.
[28]罗立新,孙铁珩.Cd和表面活性剂复合污染对小麦叶片若干生理性状的影响[J].应用生态学报,1998,9(1):95-100.
[29]孔祥生,张妙霞,郭秀璞.Cd+2毒害对玉米幼苗细胞膜透性及保护酶活性的影响[J].农业环境保护,1999,18(3):38-39.
[30]张义贤.重金属对大麦幼苗膜脂过氧化及脯氨酸和可溶性糖含量的影响[J].农业环境科学学报,2006,25(4):857-860.
[31]Chaoui A,Mazhoudi S.Cadmium and zinc induction oflipid peroxidation and effects on antioxidant enzyme activities in bean(Phaseolus vulgaris L.)[J].Plant Science,1997,127(2):139-147.
[32]Pál M,Horváth E,Janda T,et al.Physiological changes and defense mechanisms induced by cadmium stress in maize[J].Journal ofPlant Nutrition and Soil Science,2006,169:239-246.
[33]黄玉山,罗广华,关棨文.镉诱导植物的自由基过氧化损伤[J].植物学报,1997,39(6):522-526.
[34]高芳,张佳蕾,杨传婷,等.钙对镉胁迫下花生生理特性产量和品质的影响[J].应用生态学报,2011,22(11):2907-2912.
[35]李隼,黄胜东,赵福庚.重金属镉对水稻根毛细胞钾离子吸收过程的影响[J].植物生理学报,2011,47(5):481-487.
Responses of Maize during the Seedling and Heading Stage to Cadmium Availability in Soil
QI Jianying,DUTianqing,HAOJianping,XUE Jianfu,YANGZhenping,CUI Zhenkui,CHENMengni
(College of Agronomy,Shanxi Agricultural University,Taigu 030801,China)
To investigate the effects of cadmium availability in soil on maize Dafeng 30 and to analyse the difference of resistance between seeding stage and heading stage,the pot experiment was conducted to study cell membrane permeability,malondialdehyde(MDA)content,peroxidase(POD)activity and dry matter in seeding stage and heading stage of maize.The results showed that the inhibition rate of cadmium a vailability on maize growth was the highest in the heading stage(50-70 days after sowing),while was lower in the late(70-90 days after sowing).During the heading stage,the POD activity under 1 mg/kg of cadmium availability was significant lower than that under the control(0 mg/kg)(P<0.05),with no significant differences on other indicators.The threshold of the effect of cadmium availability on maize was below1 mg/kg.A quadratic model was observed between the cadmium availability concentration and the MDA content,cell membrane permeability,which showed that the variation of cell membrane permeability was more than MDA content under higher cadmium concentration.In addition,the proportion of litter with 7 mg/kg of cadmium availability was 3.1 times greater than the control during the seedling stage(P<0.05).The results could provide a further understand of the mechanism of different growth stage under Cd stress,and some theory to enrich the study on the responses of maize under cadmium stress.
cadmium availability;maize;physiology response;growth stages
S513
A
1002-2481(2016)11-1627-06
10.3969/j.issn.1002-2481.2016.11.13
2016-06-21
山西省自然科学基金项目(2014011031-1)
祁剑英(1992-),男,山西临汾人,在读硕士,研究方向:作物土壤生态。杜天庆为通信作者。
