光源或培养基成分对金花茶愈伤组织中DFR、LAR与PPO基因表达及总儿茶素含量的影响
2017-01-04钟春水赖瑞联刘生财赖钟雄
钟春水, 赖瑞联, 刘生财, 赖钟雄
( 福建农林大学 园艺植物生物工程研究所, 福州 350002 )
光源或培养基成分对金花茶愈伤组织中DFR、LAR与PPO基因表达及总儿茶素含量的影响
钟春水, 赖瑞联, 刘生财, 赖钟雄*
( 福建农林大学 园艺植物生物工程研究所, 福州 350002 )
该研究以富含儿茶素的金花茶愈伤组织为材料,对不同光源、激素、碳源及苯丙氨酸处理30 d的愈伤组织中DFR表达量、LAR表达量、PPO表达量与总儿茶素含量的变化情况及四者两两之间的相关性进行了分析。结果表明:这4个检测项目均对以上处理有显著的响应;在以上各因素处理下,DFR与LAR的表达模式十分相似,其相关系数处在0.710~0.889之间;在不同碳源处理下,PPO表达量与总儿茶素含量的变化呈显著负相关关系,其相关系数为-0.696;在不同苯丙氨酸添加量处理下,DFR与LAR表达量变化均与总儿茶素含量变化呈显著正相关关系,其相关系数分别为0.786和0.564;适宜儿茶素离体生产的金花茶愈伤组织增殖配方为附加4 mg·L-16-BA、0.6 mg·L-12,4-D、30 g·L-1蔗糖与0.660 8 g·L-1苯丙氨酸的MS固体培养基,其总儿茶素含量可达40.11 mg·g-1DW。以上研究表明,与茶树相似,在金花茶中DFR与LAR在儿茶素代谢过程中密切相关;PPO表达量升高导致金花茶儿茶素损失;添加适宜浓度的苯丙氨酸作为前体物质是提高愈伤组织中总儿茶素含量的有效措施。
光, 激素, 碳源, 苯丙氨酸, 相关性分析
金花茶(Camellianitidissima)是一种药用价值很高的木本花卉,其儿茶素具有抗氧化(Lu et al,2011 )、抗炎(Garcia et al,2013)和抑制克氏锥虫生长等作用(Paveto et al,2004)。植物组织培养具有条件可控、性状一致性好且增殖系数高的优势(王蒂,2004)。因此,以植物组织培养为平台,开展金花茶儿茶素代谢的分子机制的研究对于金花茶儿茶素深度开发具有重要意义。
DFR、LAR与PPO基因对于儿茶素的累积具有关键作用。其作用如下:DFR蛋白可逆地作用于正旋二氢槲皮素与二氢杨梅素且这个酶还参与了植物花青素的合成(Stafford & Lester,1985;Heller et al,1985; Fischer et al,2003);LAR蛋白能够催化儿茶素、无色花青素和相关的黄烷-3-羟基阿福豆素与没食子儿茶素这些植物聚合型原花青素或者浓缩单宁合成起始物质的合成(Tanner & Kristiansen,1993;Tanner et al,2003);PPO蛋白是一种含有3个铜的蛋白,这个蛋白专一地催化儿茶酚形成相对应的氧化醌且这个酶也对其他的取代型儿茶酚起作用(Gregory & Bendall,1967;Carsten et al,2002)。目前,国内外对于儿茶素代谢的分子机制研究主要集中在茶树上(Yang et al,2012;Rani et al,2012;Hong et al,2014;Liu et al,2015)而在金花茶上仅有少量报道(Zhou et al,2012,2013)且未见金花茶LAR定量表达与相关性分析的研究报道。因此,该文以富含儿茶素的金花茶愈伤组织为材料对不同光源、激素、碳源及苯丙氨酸处理下DFR基因表达量、LAR基因表达量、PPO基因表达量与总儿茶素含量的变化及相关性进行了研究。希望通过以上试验为金花茶儿茶素代谢的分子机制研究提供基础并为金花茶儿茶素离体生产提供指导。
1 材料与方法
1.1 材料
试验所需金花茶体细胞胚由福建农林大学园艺植物生物工程研究所提供。采用MS+0.5 mg·L-1KT+8.0 mg·L-1NAA+5 mg·L-1AgNO3从上述金花茶体细胞胚上诱导愈伤组织(图1:A),并采用MS+4.0 mg·L-16-BA+0.6 mg·L-12,4-D以30 d为培养周期对获得愈伤组织进行继代增殖培养(图1:B)。
1.2 金花茶愈伤组织的处理
1.2.1 不同光源处理 将愈伤组织接种在含4 mg·L-16-BA、0.6 mg·L-12,4-D和30 g·L-1蔗糖的MS固体培养基上,并分别置于LED红、LED黄、LED蓝、LED绿、LED白、LED暖白、黑暗或日光灯8种光下培养。若无特殊说明,所用培养基均以MS为基本培养基,pH均为5.8,经121 ℃、101 kPa高压灭菌20 min保存备用,培养温度为(25±2)℃,光源为日光灯,光照周期为12 h·d-1,培养时间为30 d,每个处理重复3次,下同。1.2.2 不同碳源种类与添加量处理 采用两因素三水平的完全随机试验设计研究碳源种类(白糖、蔗糖、葡萄糖)与添加量(15、30、45 g·L-1)对DFR、LAR和PPO的表达量及总儿茶素含量的影响。其他培养基成分为4 mg·L-16-BA与0.6 mg·L-12,4-D。
1.2.3 不同6-BA与2,4-D浓度处理 采用两因素三水平的完全随机试验设计研究6-BA浓度(3、4、5 mg·L-1)与2,4-D浓度(0.3、0.6、0.9 mg·L-1)对DFR、LAR和PPO的表达量及总儿茶素含量的影响。其他培养基成分为30 g·L-1蔗糖。

图 1 富含儿茶素的金花茶愈伤组织诱导与培养 A. 从体胚诱导的愈伤组织;B. 继代增殖的愈伤组织; C. 0.660 8 g·L-1苯丙氨酸处理下的愈伤组织。Fig. 1 Induction and culture of calli rich in catechins from Camellia nitidissima A. Calli induced from somatic embryo; B. Calli cultured in vitro; C. Calli treated under 0.660 8 g·L-1 PHE.
1.2.4不同苯丙氨酸添加量处理 将愈伤组织分别接种在含有0、0.082 6、0.165 2、0.330 4、0.660 8、1.321 5和2.643 0 g·L-1苯丙氨酸的7种MS固体培养基中置于日光灯下培养。其他培养基成分为4 mg·L-16-BA、0.6 mg·L-12,4-D 与30 g·L-1蔗糖。
1.3 金花茶DFR,LAR与PPO基因定量表达分析
1.3.1 金花茶RNA提取与逆转录 采用天根 RNA提取试剂盒(天根,北京)对以上不同光源、激素、碳源及苯丙氨酸添加量处理30 d后的金花茶愈伤组织进行总RNA提取。质量及浓度检测合格后,采用TAKARA PrimerscriptTMRT Reagent Kit试剂盒进行RNA逆转录用于qPCR试验。
1.3.2 qPCR引物、反应体系与反应程序 以18S为内参基因,根据文献报道的DFR(周兴文,2012)与PPO(FJ597757.1)及克隆获得的LAR(KR045740.1)基因序列分别设计引物并送北京六合华大科技有限公司进行合成。反应体系与反应程序参照Lin & Lai(2002)报道的方法并根据相关引物TM值进行改良。各引物的序列及退火温度见表1。

表 1 18S、DFR、LAR和PPO定量PCR所用引物及退火温度
1.4 金花茶儿茶素总量测定
采用香荚兰素法(魏毅等, 1999)对烘干至恒重且粉碎后过40目筛子的愈伤组织粉末进行儿茶素总量的测定。
1.5 数据分析
采用SPSS 19.0软件对数据的方差、差异显著性及相关性进行分析。
2 结果与分析
2.1 不同光源处理下愈伤组织中DFR、LAR与PPO基因的表达量和总儿茶素含量变化及相关性分析
由表2可知,LED绿与日光灯处理下DFR表达量较高; LED红、LED蓝、LED绿、LED白及日光灯处理下LAR表达量较高;LED绿处理下PPO表达量较高;LED黄处理下总儿茶素含量较高。

表 2 不同光源处理下基因表达量及总儿茶素含量的测定
注: 不同小写字母表示在0.05水平上有显著差异。下同。
Note: Different lowercase letters mean having significant differences at 0.05 level. The same below.
方差分析结果表明,光对DFR、LAR和PPO这3个基因的表达及儿茶素总量均有极显著影响。相关性分析结果表明,在不同光处理下,四项指标两两之间仅DFR与LAR之间存在显著的相关关系且其相关系数为0.71。
2.2不同6-BA与2,4-D浓度处理下愈伤组织中DFR、LAR与PPO基因的表达量和总儿茶素含量变化及相关性分析
由表3可知,6-BA3+2,4-D0.3处理下DFR表达量较高;6-BA3+2,4-D0.3处理下LAR表达量较高;6-BA4+2,4-D0.3、6-BA4+2,4-D0.9及6-BA5+2,4-D0.9处理下PPO表达量较高;6-BA3+2,4-D0.6处理下总儿茶素含量最高且与其他八个处理存在显著差异。
方差分析结果表明,6-BA、2,4-D及6-BA×2,4-D均对DFR、LAR与PPO基因这3个基因的表达及儿茶素总量存在极显著影响。相关性分析结果表明,在不同6-BA与2,4-D浓度处理下,四项指标两两之间仅DFR与LAR之间存在显著相关关系且其相关系数为0.889。

表 3 不同6-BA与2,4-D浓度处理下基因表达量及总儿茶素含量的测定
2.3 不同碳源种类与添加量处理下愈伤组织中DFR、LAR与PPO基因的表达量和总儿茶素含量变化及相关性分析
由表4可知,45 g·L-1蔗糖处理下DFR基因表达量相对较高; 45 g·L-1蔗糖处理下LAR基因表达量相对较高;15 g·L-1蔗糖或15 g·L-1葡萄糖处理下PPO基因表达量相对较高;30 g·L-1白糖处理下总儿茶素含量最高,均值为25.1 mg·g-1,且与其他8个处理均存在显著差异。
方差分析结果表明,种类、添加量及种类×添加量均对DFR、LAR与PPO3个基因的表达及儿茶素总量存在极显著影响。相关性分析结果表明,在不同碳源种类与添加量处理下,4项指标两两之间DFR与LAR之间存在极显著正相关,其相关系数为0.867,且PPO与总儿茶素含量存在显著负相关,其相关系数为-0.696。
2.4 不同苯丙氨酸添加量处理下DFR、LAR和PPO的表达量及总儿茶素含量变化及相关性分析
由表5可知,1.321 5或2.643 0 g·L-1苯丙氨酸处理下DFR基因表达量相对较高; 1.321 5或2.643 0 g·L-1苯丙氨酸处理下LAR基因表达量相对较高 0、0.082 6、0.165 2、0.330 4或2.643 0 g·L-1苯丙氨酸处理下的愈伤组织中PPO基因表达量相对较高; 2.643 04 g·L-1苯丙氨酸处理下的金花茶愈伤组织中总儿茶素含量最高,其均值为41.56 mg·g-1,且与其他7个处理存在显著差异。
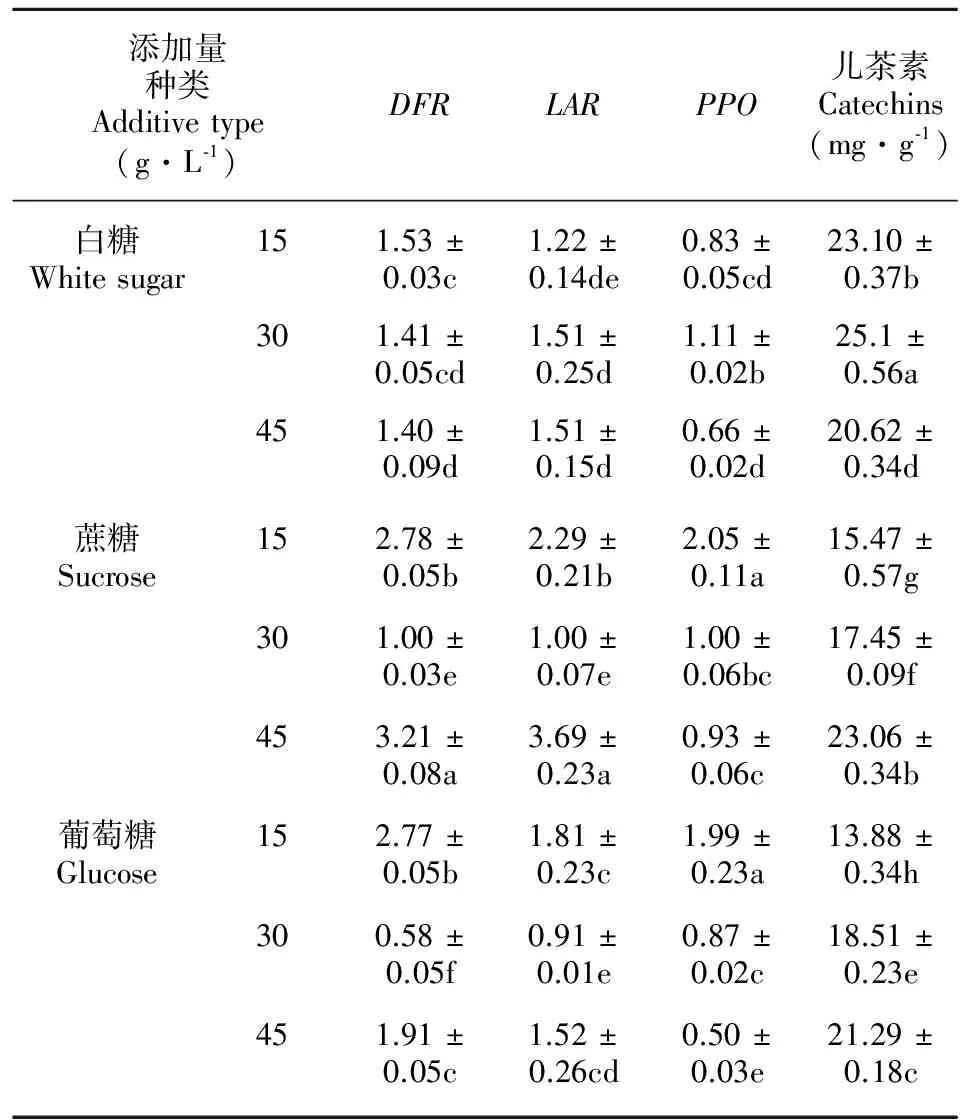
表 4 不同碳源种类与添加量处理下基因表达量及总儿茶素含量的测定

表 5 不同苯丙氨酸添加量处理下基因表达量及总儿茶素含量的测定
方差分析结果表明,苯丙氨酸添加量对DFR、LAR和PPO这3个基因的表达及儿茶素总量均有极显著影响。相关性分析结果表明,在不同苯丙氨酸添加量处理下,四项指标两两之间DFR与LAR之间存在显著正相关关系,其相关系数为0.861;DFR与总儿茶素含量存在极显著正相关关系,其相关系数为0.786;LAR与总儿茶素含量存在极显著正相关关系,其相关系数为0.564。
3 讨论
3.1 在金花茶愈伤组织中DFR与LAR在儿茶素代谢过程中密切相关
在不同光照或培养基成分处理下DFR基因与LAR基因之间均存在显著甚至极显著正相关关系。换而言之,这两个基因在这些情况下具有十分相似的表达调控模式。在茶树中,DFR蛋白与LAR蛋白在进行催化反应时,前者的产物正是后者的底物(Hong et al,2014),而且在叶片的酶学试验中也发现了两者之间有着不可分割的相关性((Punyasiri et al,2004)。
故由此推测:在金花茶中DFR蛋白与LAR蛋白在进行催化反应时前者的产物也是后者的底物且两者在儿茶素物质转化过程中密切相关。
3.2 PPO表达量升高导致金花茶愈伤组织中儿茶素损失
在不同碳源种类与添加量的处理下的愈伤组织中,PPO基因表达量与总儿茶素含量存在显著负相关关系。当碳源为蔗糖15 g·L-1或葡萄糖15 g·L-1时,PPO基因的表达量尤其高而总儿茶素的含量尤其低。前人研究表明,PPO蛋白具有催化儿茶酚形成相对应的氧化醌的功能(Gregory & Bendall,1967;Carsten et al,2002)。
故由此推测,在不同碳源处理情况下个别处理总儿茶素含量显著降低的现象可能是由PPO基因表达量显著上升造成的。也就是说PPO表达量升高导致金花茶愈伤组织中儿茶素损失。
3.3 外源添加的苯丙氨酸通过提高金花茶愈伤组织中DFR与LAR等关键基因的表达从而促进儿茶素的积累
苯丙氨酸是类黄酮途径物质合成的前体物质。在不同苯丙氨酸添加量处理下,DFR基因与LAR基因的相对表达量分别与总儿茶素总量存在极显著正相关关系而在不同光、不同碳源种类与添加量或不同6-BA与2,4-D浓度处理下,DFR基因与LAR基因的相对表达量与含总量不存在显著的相关关系。故此推测,在金花茶愈伤组织培养基中添加苯丙氨酸能够起到作为儿茶素合成前体的作用且通过提高DFR与LAR等关键基因的表达从而提高愈伤组织中总儿茶素的含量。
综合比较不同光源、激素、碳源及苯丙氨酸处理下愈伤组织中儿茶素含量,添加适宜浓度的苯丙氨酸是促进金花茶愈伤组织中总儿茶素累积更为有效的方式。
CARSTEN G, CHRISTOPH E, BERNT K, 2002. The crystal structure of catechol oxidase: new insight into the function of type-3 copper proteins [J]. Acc Chem Res, 35(3): 183-191.
FISCHER TC, HALBWIRTH H, MEISEL B, et al, 2003. Molecular cloning, substrate specificity of the functionally expressed dihydroflavonol 4-reductases fromMalusdomesticaandPyruscommuniscultivars and the consequences for flavonoid metabolism [J]. Arch Biochem Biophys, 412(2): 223-230.
GARCIA M, MICHELANGELI F, FERNANDEZ A, et al, 2013. Anti-inflammatory effects of (+)-catechin isolated from the bark ofByrsonimacrassifolia[J]. Plant Med, 79(13): 1173.
GREGORY RP, BENDALL DS, 1967. The purification and some properties of the polyphenol oxidase from tea (CamelliasinensisL) [J]. Biochem J, 101(3): 569-581.
HELLER W, FORKMANN G, BIRTSCH L, et al, 1985. Enzymatic reduction of (+)-dihydroflavonols to flavan-3,4-cis-diols with flower extracts fromMatthiolaincanaand its role in anthocyanin biosynthesis [J]. Planta, 165(2): 284-287.
HONG G, WANG J, ZHANG Y, et al, 2014. Biosynthesis of catechin components is differentially regulated in dark-treated tea (CamelliasinensisL.) [J]. Plant Physiol Biochem, 78: 49-52.
LIN YL, LAI ZX. 2010. Reference gene selection for qPCR analysis during somatic embryogenesis in longan tree [J]. Plant Sci, 178(4): 359-365.LIU M, TIAN HL, WU JH, et al, 2015. Relationship between gene expression and the accumulation of catechin during spring and autumn in tea plants (CamelliasinensisL.) [J]. Hortic Res, 2: 15023.
LU N, CHEN PQ, YANG Q, et al, 2011. Anti- and pro-oxidant effects of (+)-catechin on hemoglobin-induced protein oxidative damage [J]. Toxicolinvitro, 25(4): 833-838.
PAVETO C, GUIDA M, ESTEVA MI, et al, 2004. Anti-trypanosoma cruzi activity of green tea (Camelliasinensis) catechins [J]. Antimicrob Agents Chemother, 48(1): 69-74.
PUNYASIRI P, ABEYSINGHE I, KUMAR V, et al, 2004. Flavonoid biosynthesis in the tea plantCamelliasinensis: properties of enzymes of the prominent epicatechin and catechin pathways [J]. Arch Biochem Biophys, 431(1): 22-30.
RANI A, SINGH K, AHUJA PS, et al, 2012. Molecular regulation of catechins biosynthesis in tea [Camelliasinensis(L.) O. Kuntze] [J]. Gene, 495(2): 205-210.
STAFFORD HA, LESTER HH, 1985. Flavan-3-ol biosynthesis: The conversion of (+)-dihydromyricetin to its flavan-3,4-Diol (Leucodelphinidin) and to (+)-gallocatechin by reductases extracted from tissue cultures ofGinkgobilobaandPseudotsugamenziesii[J]. Plant Physiol, 78(4): 791-794.
TANNER GJ, FRANCKI KT, ABRAHAMS S, et al, 2003. Proanthocyanidin biosynthesis in plants-purification of legume leucoanthocyanidin reductase and molecular cloning of its cDNA [J]. J Biol Chem, 278(34): 31647-31656.
TANNER GJ, KRISTIANSEN KN, 1993. Synthesis of 3,4-cis- [3H]leucocyanidin and enzymatic reduction to catechin [J]. Anal Biochem, 209(2): 274-277.
WANG D, 2004. Tissue culture of plant [M]. Beijing: China Agriculture Press: 7. [王蒂, 2004. 植物组织培养 [M]. 北京: 中国农业出版社: 7.]
WEI Y, WANG J, LUO JY, 1999. Detection of content of catechins in tea polyphenols buccal tablet by vanillin colorimetric method [J]. Chin J Chin Mat Med, 24(6): 347-347. [魏毅, 王娟, 罗杰英, 1999. 香荚兰素比色法测定茶多酚口含片中儿茶素含量 [J]. 中国中药杂志, 24(06): 27-28.]YANG D, LIU Y, SUN M, et al, 2012. Differential gene expression in tea (CamelliasinensisL.) calli with different morphologies and catechin contents [J]. J Plant Physiol, 169(2): 163-175.
ZHOU XW, LI JY, FAN ZQ, 2012. Cloning and expression analysis ofChalconeIsomerasegene cDNA fromCamellianitidissima[J]. For Res, 25(1): 93-99.ZHOU XW, LI JY, YIN HF, et al, 2013. Cloning ofCamellianitidissimaflavonol synthase cDNA and construction of sense RNA interference expression vectors [J]. Bull Bot Res, 33(1): 58-65.
ZHOU XW, 2012. Clonning and function reach of pigment genes fromCamellianitidissima[D]. Beijing: Chinese Academy of Forestry Sciences. [周兴文, 2012. 金花茶花色相关基因的克隆及其功能研究 [D]. 北京: 中国林业科学研究院.]
Effects of light or medium components on gene expression ofDFR,LARandPPOand content of catechins in calli ofCamellianitidissima
ZHONG Chun-Shui, LAI Rui-Lian, LIU Sheng-Cai, LAI Zhong-Xiong*
( Institute of Horticultural Biotechnology, Fujian Agriculture and Forestry University, Fuzhou 350002, China )
Studies onCamelliasinensisshow that light and medium components had significant effects on metabolism of catechins in materials culturedinvitroand thatDFRgene,LARgene andPPOgene all have close relationship with it. For further study on molecular mechanism of metabolism of catechins and providing theoretical guidance for the deep development of catechins inC.nitidissima, calli rich in catechins were used as materials to study the variation ofDFRgene expression,LARgene expression,PPOgene expression and content of catechins and the correlation with each other in calli under different light source, hormones, carbon source or PHE treatments for 30 d. The results showed that all the above four detecting items reacted toinvitrotreatments significantly. Under the above treatments, the expression pattern ofDFRgene andLARgene was very similar. The correlation coefficients between the two under these treatments were between 0.710 and 0.889. The correlation ofPPOgene expression and content of catechins was significantly negative under different carbon source treatments and their correlation coefficient was -0.696.DFRgene expression was significantly positively related to the content of catechins under different PHE adding quantity treatments and the correlation coefficient was 0.786.LARgene expression was also significantly positively related to the content of catechins under different PHE adding quantity treatments and the correlation coefficient was 0.564. MS solid medium supplented with 4 mg·L-16-BA, 0.6 mg·L-12,4-D, 30 g·L-1sucrose and 0.660 8 g·L-1PHE was suitable forinvitroproduction of catechins. Content of catechins in calli cultured in this medium for 30 d was about 40.11 mg·g-1DW. Based on the above research we concluded that similar toC.sinensisthere was a close connection betweenDFRgene andLARgene during metabolism of catechins inC.nitidissimaand that the increase of expression ofPPOgene caused the loss of catechins inC.nitidissimaand that it was an effective way to add suitable amount of PHE for the increase of content of catechins in calli ofC.nitidissima.
light, hormone, carbon source, PHE, correlation analysis
10.11931/guihaia.gxzw201601015
2016-01-15
2016-03-31
福建省科技厅重大科技专项(2015NZ0002-1) [Supported by Major Science and Technology Projects of Fujian Provincial Science and Technology Department (2015NZ0002-1)]。
钟春水(1990-),男,福建龙岩人,硕士研究生,主要从事花卉生物技术研究,(E-mail)291768260@qq.com。
*通讯作者: 赖钟雄,博士,研究员,主要从事园艺植物生物技术研究,(E-mail)Laizx01@163.com。
Q945,S685.14
A
1000-3142(2016)12-1410-06
钟春水, 赖瑞联, 刘生财, 等. 光源或培养基成分对金花茶愈伤组织中DFR、LAR与PPO基因表达及总儿茶素含量的影响 [J]. 广西植物, 2016, 36(12):1410-1415
ZHONG CS, LAI RL, LIU SC, et al. Effects of light or medium components on gene expression ofDFR,LARandPPOand content of catechins in calli ofCamellianitidissima[J]. Guihaia, 2016, 36(12):1410-1415
