近无柄雅榕对盐胁迫的生理响应和引种试验
2017-01-03李贺鹏张晓勉岳春雷高智慧
郭 亮,李贺鹏,卢 刚,张晓勉,岳春雷,高智慧
(1. 浙江省台州市椒江区农业林业局,浙江 台州 318000;2. 浙江省林业科学研究院,浙江 杭州 310023;3. 浙江省林业技术推广总站,浙江 杭州 310020)
近无柄雅榕对盐胁迫的生理响应和引种试验
郭 亮1,李贺鹏2*,卢 刚2,张晓勉2,岳春雷2,高智慧3
(1. 浙江省台州市椒江区农业林业局,浙江 台州 318000;2. 浙江省林业科学研究院,浙江 杭州 310023;3. 浙江省林业技术推广总站,浙江 杭州 310020)
2011年8月,用0、0.3%、0.6%和0.9%质量浓度NaCl溶液对3年生近无柄雅榕(Ficus concinna var. subsessilis)进行处理,在处理10 d、20 d、30 d和40 d时分别测定其叶片的叶绿素含量、光合特性等指标的变化;2006年在台州湾围垦区内引种1年生近无柄雅榕,对其生长情况进行连续观测。结果表明,随着胁迫时间的延长,总叶绿素、叶绿素(Chla)和叶绿素(Chlb)的含量在0.3% NaCl处理下逐渐升高;在0.6% NaCl处理下,Chlb含量随胁迫时间延长逐渐下降,而总叶绿素和Chla含量都先降低后逐渐升高;Chla/Chlb值在0.3% NaCl处理下先升高后下降再升高,0.6% NaCl处理下的则是先下降后逐渐升高。盐胁迫明显抑制了近无柄雅榕的光合速率,在处理10 d时最为显著,随着胁迫浓度的升高抑制作用更加显著。叶片PSII最大光化学效率、潜在光化学效率、最大净光合速率和表观量子效率都随NaCl浓度的上升而逐渐下降;暗呼吸速率和光补偿点在10 d时随着胁迫浓度的上升而逐渐下降,在处理20 d和30 d时逐渐升高,在处理40 d时先升高后降低。引种试验观察表明,近无柄雅榕苗的成活率达100%,平均年高生长约30 cm、地径1.26~2.14 cm;零度以下气温会对5年生以下的树苗造成严重冻害。
近无柄雅榕;盐胁迫;叶绿素;光合特性;滨海滩涂;引种
我国沿海18 000 km的滨海地带和岛屿沿岸广泛分布着各种滨海盐土,总面积达5×106hm2[1]。滨海土往往由滩涂围垦形成,土壤表现为中高度盐碱、板结、养分缺乏、高水位、海水倒灌反盐等,不利于植物生长[2],过多的可溶性盐类严重制约这些地区绿化造林的质量和数量[3]。为了丰富滨海滩涂造林树种,通过对耐盐植物引进、选育、区域试验,发展耐盐植物改良盐碱地是目前全世界两类主导技术中的一类[4]。研究逆境胁迫对植物的影响以及植物的反应,特别是光合作用作为绿色植物逆境胁迫环境下较为敏感的生理生态指标,其在胁迫环境下的变化特征对于阐明植物的盐害机理、改良盐渍地等有重要意义。
近无柄雅榕(Ficus concinna var. subsessilis)又称无柄小叶榕,是雅榕(Ficus concinna)的一个变种,主要分布在浙江南部、云南、广东等地[5],是我国亚热带最北缘的榕属常绿树种,不仅树姿雄伟,且具有一定耐盐性,是城市绿化和绿化造林优良树种[2,5]。有关近无柄雅榕的研究主要集中在种子种苗繁育[6~10],以及高低温胁迫、盐胁迫下的光合作用、抗逆性等方面的研究[5,11~13];尽管林霞等[5]研究了不同NaC1浓度胁迫7 d内对近无柄雅榕气体交换、叶绿素荧光和抗氧化酶活性的影响,以及李琳等[13]开展了海水胁迫下2年生幼苗形态、存活率、SOD活性等相关研究,但是关于NaCl胁迫下近无柄雅榕的光合作用、生理特性等的长期影响等方面未见报道。
近无柄雅榕自然分布北缘为温州市,对干旱和低温霜害相对敏感[6,12]。受温室效应影响,全球气温逐年升高,这为其种群进行北移扩区提供了可能,因此开展北移引种试验研究显得尤为重要。2003年浙江省台州市椒江区从福建省漳州市引种胸径40 cm以上的近无柄雅榕大树与1年生实生裸根小苗,在台州湾围垦滩涂盐碱地苗圃进行培育与试种。本试验通过对近无柄雅榕引种后的试验观测,结合耐盐性试验,研究其耐盐能力和进行北移扩区的可能性,探讨近无柄雅榕光合特性对长期盐胁迫的响应特征,为盐碱地引种、驯化提供理论依据。
1 材料与方法
1.1 耐盐性试验
1.1.1 试验材料 选取的试验材料为3年生近无柄雅榕实生苗,2011年5月中旬在浙江省林业科学研究院温室大棚内进行盆栽适应试验。花盆选用规格为:上口直径25.5 cm,下口直径16.0 cm,高19.0 cm。栽培基质为苗圃熟土,常规浇水培育,并进行适当管护。经过近3个月的适应后,于2011年8月选取长势一致的苗木进行NaCl胁迫处理。试验设置1个对照(CK)清水处理和0.3%、0.6%和0.9%质量浓度NaCl浇注。每个处理各种植3盆,即每盆种植1株;试验期间观察盐胁迫对植物生长的影响情况,主要包括叶片颜色变化、叶片脱落、植株生长等,记录至83 d。
1.1.2 光合-光响应曲线和叶绿素荧光参数的测定 利用LI-6400便携式光合作用测定系统(Licor,USA)对光响应曲线进行测定。选取自顶端向下的第3、第4张叶片,采用标准叶室(2 cm×3 cm)并使用红蓝光源(Red Blue 6400-02B),设定光合有效辐射为0、20、50、100、200、400、600、800、1 000、1 200、1 500和2 000μmol•m-2•s-1。
选用标准荧光叶室(6400-40 Default Flurometer)进行叶绿素荧光参数的测定。首先采用配套的暗适应叶夹对各个处理相同部位功能叶进行30 min的遮光处理,立即测定其初始荧光(Fo)、最大荧光(Fm)、PSII(photosystem II)最大光化学效率(Fv/Fm)、PSII潜在光化学效率(Fv/Fo)。选取的叶片与光合-光响应曲线测量的相同。
上述参数每隔10 d测定一次,均为晴朗天气。
1.1.3 叶绿素含量的测定 选取与测定光合作用和叶绿素荧光相同位置和发育阶段的叶片,用纯丙酮、无水乙醇和蒸馏水的混合液法提取叶绿素,并用紫外分光光度计测定吸光值,然后根据公式计算得出叶绿素含量[14]。
1.2 引种试验
1.2.1 试验地概况 试验地位于浙江省台州市台州湾南段十塘围垦滩涂盐碱地,年平均气温17.0℃,绝对最低气温 -6.8℃,年平均降水量1 537.0mm,其中3-6月春雨和梅雨占全年降雨量的43%,8-9月台汛期占27%,年蒸发量1 334.0 mm。土壤为滨海盐土,由粒径小于63 µm的粘土质粉砂组成,pH值8.1 ~ 9.2,0 ~ 90 cm土层土壤平均含盐量为5.64 g/kg,属重盐化土壤。自然植被主要有碱蓬(Suaeda glauca)、盐蒿(Artemisia halodendron)等群落。
1.2.2 土壤采样与分析 分别于2006年3月和2009年3月,以及2011年4月5日、7月28日、10月18日和2012年1月18日进行土壤取样,在实验地随机选取5个取样点采集表层0 ~ 10 cm土样,充分混合,风干后测定土壤含盐量。土壤含盐量采用LY/T 1251-1999标准测定。
1.2.3 苗木栽植及生长调查 2006年3月选择1年生近无柄雅榕实生裸根小苗按2.5 m×2.5 m的株行距进行栽植试验,引种试验面积15 hm2,于2007-2014年每年的11月,在3个10 m×10 m的固定样地测量46棵植株的树高和地径,并统计植株的存活率。
1.3 数据处理与分析
1.3.1 光响应曲线特征参数提取 采用非直角双曲线模型对光响应曲线进行拟合[15]:
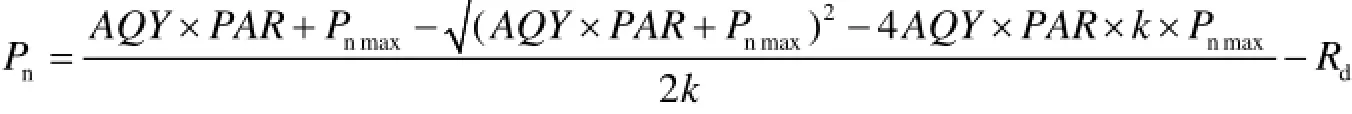
式中:Pn(net photosynthetic rate)为净光合速率(μmol•m-2•s-1);AQY (apparent quantum yield)为表观量子效率(μmol•m-2•s-1);PAR(photosynthetically available radiation)为光合有效辐射(μmol•m-2•s-1);Pnmax为最大净光合速率(μmol•m-2•s-1);k为曲线的曲角;Rd为暗呼吸速率(μmol•m-2•s-1)。
利用在0 ~ 200 μmol•m-2•s-1的 Pn对PAR进行线性回归,运用所得出的直线方程与前述中得出的Pnmax值,可计算出光补偿点(LCP, light compensate point)、AQY等参数。200 μmol•m-2•s-1以下的线性方程,与非直线双曲线模型计算出的Pnmax值和X轴这两条平行直线相交,可得出交点。其与X轴的交点数值即为光补偿点(LCP,μmol•m-2•s-1),其中AQY可表示为该直线方程的斜率[16]。
1.3.2 数据分析 应用SPSS 13.0统计分析软件进行统计分析,并采用LSD法进行多重比较和显著性检验;应用Origin 7.5进行数据图绘制。
2 结果与分析
2.1 盐胁迫对近无柄雅榕生长的影响
处理5 d时,0.6% NaCl处理组的每株近无柄雅榕树苗部分叶片颜色变深,1株树苗脱落10余片叶片;0.9% NaCl处理组的3株树苗均有20多片叶片脱落。处理8 d时,0.6% NaCl处理组的每株树苗脱落叶片数量增加,1株树苗较严重;0.9% NaCl处理组的3株树苗的接近90%叶片脱落。处理11 d时,0.6% NaCl处理组脱落叶片变化不大;0.9% NaCl处理组的3株树苗叶片接近全部脱落,个别残留在枝条上的叶片呈浅黄色或深黑色。处理18–20 d时,0.6% NaCl处理组每株树苗生长变化不大,但叶片脱落数增多,其中1株接近80%的叶片脱落,其它2株接近30%;0.9% NaCl处理组树苗全部脱落。
处理27 d时,0.3% NaCl处理组每株树苗有少量叶片脱落;0.6% NaCl处理组1株树苗接近死亡,枝头仅剩2片叶片,其它2株脱落叶片超过整株的40%。处理32 d时,0.3% NaCl处理组每株树苗变化不大;0.6% NaCl处理组1株树苗死亡,其它2株脱落叶片数量少量增加。处理40 d时,0.3% NaCl处理组每株树苗变化不大;0.6% NaCl处理组2株树苗脱落叶片数量超过60%。处理46 d时,0.3% NaCl处理组每株树苗变化不大;0.6% NaCl处理组2株树苗脱落叶片数量接近70%。处理54 d时,0.3% NaCl处理组每株树苗的叶片脱落接近30%;0.6%NaCl处理组2株树苗脱落叶片数量接近80%。处理60 d时,0.3% NaCl处理组每株树苗都有少量叶片脱落,少量叶片发黄;0.6% NaCl处理组2株树苗变化不大。处理66 d时,0.3%处理组树苗变化不大;0.6%处理组2株树苗变化不大。处理83 d时,0.3% NaCl处理组树苗变化不大;0.6% NaCl处理组2株树苗脱落叶片将近90%。
2.2 盐胁迫对近无柄雅榕叶绿素指标的影响
不同浓度盐胁迫下近无柄雅榕的叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素含量及Chla/Chlb均有显著的变化(图1)。
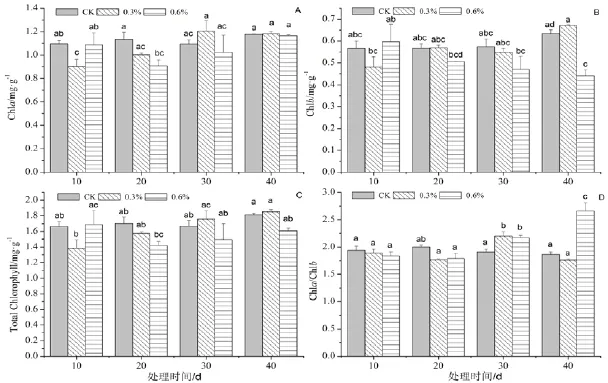
图1 盐胁迫对近无柄雅榕树苗叶片叶绿素指标的影响Figure 1 Effect of NaCl stress on chlorophyll content in leaves of F. concinna var. subsessilis
Chla含量在0.3% NaCl处理组基本上呈逐渐升高趋势、0.6% NaCl处理组呈先降低后逐渐升高的变化趋势(图1A);处理10 d时,0.3%处理组的Chla含量显著低于对照和0.6% NaCl处理组(p<0.05),并且为最低含量,为0.903 mg/g;在处理20 d时,Chla含量随胁迫浓度的升高而明显下降,其中0.6% NaCl处理组的显著低于对照(p<0.05),但与0.3% NaCl处理组的差异不显著(p>0.05);在处理30 d和40 d时,各处理组间的差异不显著。
Chlb含量在0.3% NaCl处理组是随时间延长而逐渐升高,在处理40 d时显著高于其他时段(p<0.05);Chlb含量的变化在0.6% NaCl处理组与0.3% NaCl处理组相反,呈逐渐下降的变化,但在处理40 d时显著低于处理10 d(p<0.05)时的,且为各处理组中最低含量,为0.440 mg/g(图1B)。
总叶绿素含量在0.3% NaCl处理组呈逐渐升高的变化趋势,而0.6% NaCl处理组呈先下降而逐渐升高的变化趋势(图1C);处理10 d时,0.3% NaCl处理组显著低于0.6% NaCl处理组外,但与对照组差异不显著;在处理20 d、30 d和40 d时,各处理组间的差异不显著。
0.3 % NaCl处理组的Chla/Chlb值随时间的变化呈先升高后下降再升高,0.6% NaCl处理组的则是先下降后逐渐升高;处理10 d和20 d时,各处理组间的差异不显著;处理30 d时的各处理组均显著高于对照(p<0.05);处理40 d时,0.6% NaCl处理组的Chla/Chlb值显著高于其他处理(p<0.05),0.3% NaCl处理组的低于对照,但二者间差异不显著(p>0.05)(图1D)。
2.3 盐胁迫对近无柄雅榕光响应曲线的影响
植物光合作用光响应曲线反映了光强与植物Pn的关系,不同浓度盐胁迫影响了近无柄雅榕的Pn(图2)。盐胁迫初期(图2A)近无柄雅榕的光合效率下降最为明显,且随着NaCl浓度的增加而下降,盐胁迫下植物的Pn基本在0点波动;低光强(200 μmol•m-2•s-1)下0.3% NaCl处理组的Pn呈线性上升,但是随着PAR的增强而波动较小。在处理20 d时(图2B),0.3% NaCl处理组得到明显恢复,但0.6% NaCl处理组Pn几乎在0点波动。处理30 d时,各处理组的Pn变化趋势与处理20 d时相比变化不大(图2C)。处理40 d时,不同浓度NaCl胁迫下各处理组的Pn在0点范围内波动(图2D)。这些结果说明盐胁迫明显抑制了近无柄雅榕的光合作用。

图2 盐胁迫下近无柄雅榕树苗叶片的Pn-PAR响应Figure 2 Effect of NaCl stress on Pnto PAR in F. concinna var. subsessilis leaves
2.4 盐胁迫对近无柄雅榕叶绿素荧光的影响
由图3可知,PSII最大光化学效率Fv/Fm随NaCl浓度的增加而呈逐渐下降的变化趋势。0.3% NaCl处理组明显低于对照,但差异不显著(p>0.05);0.6% NaCl处理组随着处理时间的延长呈逐渐下降再上升的变化趋势,除处理10 d时与对照差异不显著外,其他时段都显著低于对照(p<0.05),且处理30 d时的Fv/Fm值最低,为0.597。这说明在盐胁迫下,叶片光能转化效率受到明显抑制作用,且随着盐浓度的增加受到的抑制作用越强,叶片光合机构的功能在盐胁迫后期逐渐恢复。
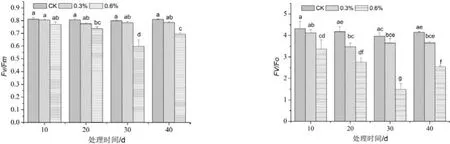
图3 盐胁迫对近无柄雅榕叶片PSⅡ最大光化学效率和潜在光化学效率的影响(p<0.05)Figure 3 Effect of NaCl stress on Fv/Fm and Fv/Fo in leaves of F. concinna var. subsessilis
PSII潜在光化学效率Fv/Fo随着NaCl浓度的增加呈逐渐下降的变化趋势。0.3% NaCl处理组除在处理20 d时显著低于对照组(p<0.05)外,其他各阶段明显低于对照但差异不显著。0.6% NaCl处理组的Fv/Fo值均显著低于其他各处理组(p<0.05),且在处理30 d时为最低,为1.486。这说明盐胁迫对近无柄雅榕叶片的光合器官发生严重的光抑制,高度盐度的胁迫更加明显,但随时间的延长,其抑制作用逐渐减弱。
2.5 盐胁迫对近无柄雅榕叶片光响应特征参数的影响
由表1可知,NaCl胁迫条件下近无柄雅榕的最大净光合速率(Pnmax, maximum net photosynthetic rate)在各个阶段都是随着胁迫浓度的增加而明显下降;Pnmax值的下降比例在处理10 d时最大,随着胁迫时间延长都有不同程度的恢复;0.3%浓度NaCl处理在处理20 d和30 d时的Pnmax值下降比例明显低于胁迫初期,但在处理40 d的的Pnmax值的下降比例又略有上升;0.6%浓度NaCl处理Pnmax值下降比例随着时间延长而逐渐降低。这说明盐胁迫在初期对植物的Pnmax值影响较为明显,植物在后期的生长中主动适应胁迫环境,致使后期各阶段的Pnmax值得到明显的恢复。
AQY的变化趋势与Pnmax值的相似,AQY值随着NaCl胁迫浓度的增加而逐渐降低(但处理40 d时的AQY 值在0.3%浓度NaCl处理低于0.6%浓度NaCl处理除外)。Rd在处理10 d时随NaCl胁迫浓度上升而逐渐下降,以及处理20 d时的0.3%浓度NaCl处理表现出下降外,其他各浓度处理均有不同程度的上升;处理30 d时的Rd值随着胁迫浓度的增加而明显上升,处理40 d时的Rd值的增加比例较大的是0.3%浓度NaCl处理。LCP在处理10 d时随NaCl胁迫浓度升高为明显下降;但在处理20 d和30 d时,LCP值随NaCl胁迫浓度升高为明显增加,且处理20 d时0.6%浓度NaCl处理下LCP值最高,增加了28.27倍。
从近无柄雅榕光合参数对NaCl盐胁迫的总体反应看,初期胁迫对各参数影响明显,且随着胁迫浓度的上升而所受抑制作用越明显,随着胁迫时间延长植物的生理适应性发挥作用,各指标呈逐渐转好的趋势。

表1 盐胁迫对近无柄雅榕叶片光合生理参数的影响Table 1 Effect of NaCl stress on photosynthetic parameters in leaves of F. concinna var. subsessilis

图5 近无柄雅榕种植区不同时期土壤含盐量的变化Figure 5 Soil salt content in sample plots during different periods
2.6 近无柄雅榕引种试验
2.6.1 引种区土壤盐度变化 椒江区十塘围垦区于1992年完成合围,塘内初期形成的土壤立地条件比较差,随着引种时间的延长,围垦区表层土壤含盐量呈明显下降的变化趋势(图5)。近无柄雅榕树苗在2006年初种植于椒江区十塘围垦区苗圃地,当时表层土壤含盐量平均值为5.6 g/kg,3 a后(2009年3月)土壤含盐量明显下降,但与树苗种植初期(2006年3月)差异不显著(p>0.05);但随着时间的延长,5 a后(2011年4月)的土壤含盐量显著(p<0.05)低于种植初期,说明在围垦后盐碱地的含盐量随着时间的延长而呈明显下降的变化。滨海围垦区夏季(2011年8月)表层土壤含盐量显著(p<0.05)高于其他各个时期;秋季(2011年10月)的土壤含盐量与春季和夏季相比都显著下降(p<0.05);冬季(2012年1月)土壤含盐量最低,但与秋季相比略有下降但差异不显著(p>0.05)。这说明围垦区表层土壤含盐量随围垦区近无柄雅榕种植时间延长而逐渐下降,植物的生长活动能不同程度地改良土壤,同时夏季高温可导致表层土壤具有较高含盐量。
2.6.2 近无柄雅榕抗寒能力 近无柄雅榕抗风性强、移植后树势恢复快,另外在移植时不需修剪摘叶、一次成型,主干不易折断,台风等气候灾害后树势也恢复较快。引种期间,2010年12月15日台州市出现暴雪冰冻极端天气,为椒江区自1966年以来12月同期的最低,16日日平均气温 -4℃,对近无柄雅榕造成严重危害,大部分叶片冻枯,当年生枝条大部分受害,2年生枝少数受害。冻害发生后的半个月(2011年1月初),叶片全部变成褐色;冻害发生后3个月(2011年3月),嫩叶、上部枝条及叶片均受害;2011年9月调查时,植株已恢复生长。
2.6.3 近无柄雅榕树高和地径历年生长量 引种试验的近无柄雅榕生长良好,成活率达100%。从表2可以看出,除了2011年的高生长与2010年的相比差异不显著(p>0.05)外,其他各年份都显著增加(p<0.05),平均每年高生长为30 cm左右,其中2013年的高生长最大,约90 cm。近无柄雅榕的地径每年平均增加1.26 ~ 2.14 cm,其中2013年的增加幅度最大,为2.14 cm;8 a间,近无柄雅榕地径每年的增加幅度都非常显著(p<0.05)。生长缓慢的阶段是由于受2010年12月的冻害影响所致。

表2 引种近无柄雅榕生长情况(n=46)Table 2 Growth of F. concinna var. subsessilis at sample plots
3 讨论与结论
已有研究证实近无柄雅榕能够在土壤盐含量0.5% ~ 0.8%的温州市龙湾等地的沿海基干林带或新围垦盐碱地上生长,存活率超过80%[2]。本研究结果与上述研究结果相一致,即在台州市椒江区围垦区引种的近无柄雅榕不仅能够在含盐量为5.6 g/kg的盐碱土中生长,而且即使在夏季返盐季节(含盐量为7.64 g/kg)也不受影响,进一步证实了近无柄雅榕是耐盐碱植物[13]。有些盐胁迫试验中,0.6% NaCl处理下1株树苗死亡,2株叶片脱离严重,这可能与短期内高盐环境影响植物的正常代谢水平导致死亡[17],另外也可能受盆栽容器、大棚内高温等因素影响所致。
逆境中植物叶片叶绿素含量变化是衡量植物抗逆性强弱的重要生理指标之一。目前对盐胁迫下植物叶绿素含量变化的机理尚未形成统一认识,一些研究证实土壤盐胁迫可能破坏植物叶绿体结构,使体内叶绿素含量下降,引起植株光合能力减弱[18~19];但也有学者认为,低钠盐可以增加植物体内叶绿素含量[20~21]。本研究结果中0.6% NaCl处理明显降低了Chla、Chlb和总叶绿素含量,且随胁迫时间的延长而呈明显下降趋势,这与盐分浓度越高、处理时间越长、影响越明显的结论[22~23]相一致;但是0.3% NaCl处理下各指标在不同时期呈差异性的变化(图1A、B和C),如Chla和Chlb分别在处理30 d和40 d时呈明显升高,总叶绿素含量在处理30 d和40 d时呈明显升高,Chla/Chlb值在30 d时明显升高。这进一步证实即使同一种植物在不同盐胁迫浓度和处理时间下,其叶绿素指标的变化规律并不明显[5]。盐胁迫下的植株叶片Chla/Chlb值与其他叶绿素指标相比相对较高,这与其他结果[23~24]相一致,主要是盐胁迫会提高叶绿素酶对Chlb的降解速度,而对Chla的影响相对较小[23]所致。
盐胁迫不仅可以直接影响近无柄雅榕的生长发育,同时也会降低植物的光合速率,抑制光合作用,这与其他研究结果[25~26]相一致。盐胁迫10 d和40 d时的光合速率下降幅度较大,特别是胁迫初期尤为明显,但是海滨木槿[17]和邓恩按[22]的光合速率下降幅度较大的时段则是在胁迫20 d时,这可能与植物的种类有关,不同植物对盐胁迫的响应机制不同所致。
PSⅡ最大光化学效率Fv/Fm和Fv/Fo可反映逆境胁迫对植物光合系统的破坏程度。本研究中近无柄雅榕在盐胁迫下Fv/Fm和Fv/Fo均低于对照,且随着胁迫浓度的升高而明显下降(图3和图4),表明NaCl胁迫破坏叶绿体光合机构,PSⅡ反应中心受损,抑制光合作用的原初反应,阻碍光合电子传递的过程,明显降低PSⅡ原初光能转换效率,抑制PSⅡ潜在活性[27~28],最终导致净光合速率的显著下降,这是植物在逆境下以降低光合速率等为代价维持自身生长的需要。
盐胁迫对近无柄雅榕树苗的Pmax、AQY、Rd、LCP等具有明显影响。一定的环境条件下,叶片的Pmax反映了植物叶片的最大光合能力[29]。本研究中盐胁迫都使近无柄雅榕的Pmax、AQY降低,这可能是胁迫条件降低了Rubisco活性和电子传递速率[30]所致。研究表明植物AQY理论值在0.083~0.125,而在自然条件下远低于理论值(0.04~0.07)[31]。本研究由非直角双曲线模型拟合得到不同盐胁迫条件下的近无柄雅榕AQY在0.002 2~ 0.041 9,表明盐胁迫致使AQY值下降至远低于自然条件的数值。不同植物的光响应曲线不同,Rd、LCP和LSP有很大的差异,同一种植物的LCP和LSP也不是固定数值,它们会随外界条件的变化而变动[32]。本研究中盐胁迫初期,随着胁迫浓度的继续升高,Rd呈现下降趋势,LCP亦下降,这与其他研究结果[33~34]相一致。随着胁迫时间的延长,植株叶片LCP和Rd随胁迫浓度升高而迅速上升,Fv/Fm和Fv/Fo、及AQY则快速下降,表明盐胁迫导致植株光合产物消耗增大,对弱光的吸收、转换和利用能力受到抑制,强光下则发生光抑制,叶片吸收的光能不能有效地转化为化学能,导致叶片光合能力下降。
较高纬度地区的低气温是制约常绿阔叶树种向北方引种栽培的重要限制因子,引种地区的低温冷害对引种植物是严重的自然灾害。近无柄雅榕天然分布区的最北界为温州市的永嘉和乐清,且数量极少[35]。台州地区引种的近无柄雅榕能够正常生长,即使遇到2010年12月的极端低气温[36],也能经过一段恢复期后逐渐恢复生长。另外,对台州市内近无柄雅榕进行了耐寒调查发现,苗圃中3 ~ 5年生幼树在采取防寒并及时清除树冠积雪等措施后,除当年秋梢轻微冻害外,基本无影响;胸径30 cm以上大树在台州市椒江区广场、单位庭院、小区、行道等城市绿化中推广应用较多,栽植已超过7 a的植株能够适应当地的霜、雪、冰冻、台风等恶劣气候环境。
综上所述,近无柄雅榕对NaCl胁迫具有一定的耐受能力,可以耐受盐度0.6%及其以下浓度;盐胁迫条件下,光合生理参数表现出一定的适应性。近无柄雅榕幼树随着树龄增大而抗寒性增强,不足7年生的幼树目前在台州市防护林建设推广中有一定难度(若遇极寒气候需采取防寒措施),但8年生以上的大树可以在台州市及其以南地区的城市园林绿化和椒江河口以南的含盐量低于0.6%的滨海地区防护林建设中推广应用。另外,胁迫条件下不同时期的光合生理参数不仅能更好地反映植物的生态特性,对于了解植物对胁迫环境的适应策略与机制,以及耐性树种的选育等方面都有重要意义。
[1] 徐恒刚. 中国盐生植被及盐渍化生态治理[M]. 北京:中国农业科学技术出版社,2004. 1 112.
[2] 孔强,陈秋夏,吴良文,等. 不同立地条件下东方杉等沿海防护林树种的生长生态特性研究[J]. 中国农学通报,2011,27(28):40-46.
[3] 宋丹,张华新,白淑兰,等. 植物耐盐种质资源评价及滨海盐碱地引种研究与展望[J]. 内蒙古林业科技,2006,1:37-38,44.
[4] 王佳丽,黄贤金,钟太洋,等. 盐碱地可持续利用研究综述[J]. 地理学报,2011,66(5):673-684.
[5] 林霞,郑坚,陈秋夏,等. NaCl胁迫对光合作用和抗氧化酶活性的影响[J]. 北京林业大学学报,2011,33(44):70-74.
[6] 林霞,郑坚,刘洪见,等. 不同基质对无柄小叶榕容器苗生长和叶片生理特性的影响[J]. 林业科学,2010,46(8)8:63-70.
[7] 林霞,郑坚,陈秋夏,等. 无柄小叶榕容器育苗轻型基质配方筛选[J]. 浙江林学院学报,2008,25(3):401-404.
[8] 王法格,李宇,柴芬友,等. 无柄小叶榕扦插育苗试验[J]. 现代农业科技,2007,(10):10-11.
[9] 陈秋夏,郑坚,林霞,等. 不同容器对木荷等3个容器苗生长的影响[J]. 西北林学院学报2009,24(2):75-79.
[10] 郑坚,陈秋夏,李效文,等. 无柄小叶榕容器苗形态和生理质量评价指标筛选[J]. 中国农学通报,2010,26(15):141-148.
[11] 金松恒,李雪芹,王俊刚. 高温胁迫对无柄小叶榕光合作用的影响[J]. 中国农学通报,2009,25(3):83-87.
[12] 张晓勉,高智慧,李明良,等. 无柄小叶榕耐寒性研究[J]. 浙江林业科技,2015,35(3):26-30.
[13] 李琳,林夏珍,刘志高,等. 海水胁迫下4种苗木的生长与生理特性[J]. 西北农业学报,2013,22(5):116-123.
[14] 陈福明,陈顺伟. 混合液法测定叶绿素含量的研究[J]. 浙江林业科技,1984,4(1):4-8.
[12] 张晓勉,高智慧,李明良,等. 无柄小叶榕耐寒性研究[J]. 浙江林业科技,2015,35(3):26-30.
[13] 李琳,林夏珍,刘志高,等. 海水胁迫下4种苗木的生长与生理特性[J]. 西北农业学报,2013,22(5):116-123.
[14] 陈福明,陈顺伟. 混合液法测定叶绿素含量的研究[J]. 浙江林业科技,1984,4(1):4-8.
[15] Farquhar G D, Van Caemmerer S. Modeling of photosynthetic response to environmental conditions[C]. Lange OL, Nobel PS, Osmond CB. Physiological Plant Ecology Ⅱ. Encyclopedia of Plant Physiology. New Series ,Berlin:Springer-Verlag,1982:549-587.
[16] 陆燕元,马焕成,李昊民,等. 土壤干旱对转基因甘薯光合曲线的响应[J]. 生态学报,2015,35(7):2 155-2 160.
[17] 卢刚,李贺鹏,张晓勉,等. 盐胁迫对海滨木槿幼苗生长及光合特性的影响[J]. 浙江林业科技,2015,35(3):16-25
[18] 王素平,李娟,郭世荣,等. NaC1胁迫对黄瓜幼苗植株生长和光合特性的影响[J]. 西北植物学报,2006,26(3):455-461.
[19] 汪贵斌,曹福亮. 盐分和水分胁迫对落羽杉幼苗的生长量及营养元素含量的影响[J]. 林业科学,2004,40(6):56-62.
[20] Munns R. Physiological processes limiting plant growth in saline soils: some dogmas and hypotheses [J]. Plant Cell Environ,1993,16(1):15-24.
[21] Munns R, Temmat A. Whole-plant responses to salinity [J]. Aust J Plant Physiol,1986,13(1):143-160.
[22] 林益,瓮颖,李贺鹏,等. 邓恩桉幼苗生长和光合特性对盐胁迫的响应[J]. 浙江林业科技, 2014,34(3):33-38.
[23] 王丹,万春阳,侯俊玲,等. 盐胁迫对甘草叶片光合色素含量和光合生理特性的影响[J]. 热带作物学报,2014,35(5):957-961.
[24] Carter DR, Cheeseman J M. The effects of external NaCl on thylakoid stacking in lettuce plants[J].Plant Cell Environ,1993,16(2):215-222.
[25] Mehta P, Jajoo A, Mathur S, et al. Chlorophyll a fluorescence study revealing effects of hish salt stress on Photosystem II in wheat leaves[J]. Plant Physiol Biochem,2010,48:16-20.
[26] Kahn N A. NaCl-inhibited chlorophyll synthesis and associated changes in ethylene evolution and antioxidative enzyme activities in wheat[J]. Biol Plant,2003,47(3):437-440.
[27] Everard J D, Gucci R, Kann S C, et al. Gas exchange and carbon partitioning in the leaves of celery (Apium graveolens L.) at various levels of root zone salinity [J]. Plant Physiol,1994,106(1):281-292.
[28] Shabala S N, Shabala S I, Martynenko A I, et al. Salinity effect on bioelectric activity growth Na+ accumulation and chlorophyll fluorcscence of corn leaves: a comparison survey and prospects for screening [J]. Aust J Plant Physiol,1998,25(5):609-6l6.
[29] 林叶春,曾昭海,任长忠,等. 局部根区灌溉对裸燕麦光合特征曲线及叶绿素荧光特性的影响[J]. 作物学报,2012,38(6):1 062-1 070.
[30] Walting J R, Press M C, Quick W P. Elevated CO2 induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum[J]. Plant Physiol,2000,123(3):1 143-1 152.
[31] 余叔文. 植物生理学和分子生物学[M]. 北京:科学出版社,1992:236-243.
[32]卜令铎,张仁和,常宇,等. 苗期玉米叶片光合特性对水分胁迫的响应[J].生态学报,2010,30(2):5-11.
[33] Zhou HH, Chen YN, Li WH, et al. Photosynthesis of Populus euphratica in relation to groundwater depths and high temperature in arid environment,northwest China[J]. Photosynthetica,2010,48(2):257-268.
[34] 王欢利,曹福亮,刘新亮. 高温胁迫下不同叶色银杏嫁接苗光响应曲线的拟合[J]. 南京林业大学学报(自然科学版),2015,39(2):14-20.
[35] 林霞. 无柄小叶榕发芽与生长对不同生态因子的响应研究[D]. 北京:北京林业大学,2010. 27-33.
[36] 李学斌,何剑英. 2010年冬台州市椒江区果树雪害调查浅析[J]. 浙江柑桔,2011,28(2):19-21.
Physiological Responses of Ficus concinna var. subsessilis Seedingsto Salt Stress and Introduction Test
GUO Liang1,LI He-peng2*,LU Gang2,ZHANG Xiao-mian2,YUE Chun-lei2,GAO Zhi-hui3
(1. Jiaojiang Agroforestry Bureau of Taizhou, Taizhou 31880, China; 2. Zhejiang Forestry Academy, Hangzhou 310023, China; 3. Zhejiang Forestry Extension Administration, Hangzhou 310020, China)
Experiments were conducted on 3-year Ficus concinna var. subsessilis by salt stress (0, 0.3%, 0.6% and 0.9%) in 2011. Determinations were made on chlorophyll content and photosynthetic index on the 10th, 20th, 30thand 40thday after treatment. In 2016, introduction was made on 1-year F. concinna var. subsessilis seedlings at reclamation area of Taizhou Bay, Zhejiang province. The results showed that under 0.3% NaCl stress, content of total chlorophyll(Chl), Chla and Chlb gradually increased. Under 0.6% NaCl stress, content of Chlb decreased, while content of Chl and Chla decreased from the 10thto 20thday and then gradually increased later. The Chla/Chlb value increased from 10thto 20thday, decreased from 21stto 30thday, and increased from 31stto 40thday under 0.3% NaCl stress. Under 0.6% NaCl stress, Chla/Chlb value decreased from the 10thto the 20thday, and then increased gradually from 21stday to 40thday. The experiment resulted that salt stress significantly inhibited the net photosynthetic rate of the seedlings, especially on the 10thday, and inhibition was increased with the NaCl concentration. Photochemical efficiency of photosystemⅡ,maximum net photosynthetic rate, apparent quantum efficiency decreased with the increase of salt concentration. Light compensation point and dark respiration rate of the seedling decreased with the increase of salt concentration on the 10thday, increased on the 20thand 30thday. Introduction test and observation from 2006 to 2014 demonstrated that survival rate of introduced seedlings topped to 100%, with annual height growth of 30cm and ground diameter of 1.26-2.14cm. Temperature under zero could cause severe injury to seedlings under 5-year.
Ficus concinna var. subsessilis; salt stress; chlorophyll; photosynthetic characteristics
S718.5
A
1001-3776(2016)04-0001-09
2016-01-25;
2016-05-29
浙江省院所专项项目(2014F50017)、浙江省省院合作项目(2011SY04)、台州市科技局应用技术研究项目(091TG02)、浙江省重点科技创新团队项目(2011R50027)、“台州湾围垦区植物区系及耐盐性研究与种植示范”项目资助
郭亮(1958-),男,浙江温岭人,高级工程师,从事森林生态研究;*通讯作者。
