腺苷A2A受体拮抗剂SCH58261减轻胎兔缺氧缺血性脑损伤*
2017-01-03柳艳丽唐震海王能里林振浪
柳艳丽, 唐震海, 王能里, 林振浪
(温州医科大学附属育英儿童医院新生儿科, 浙江 温州 325027)
腺苷A2A受体拮抗剂SCH58261减轻胎兔缺氧缺血性脑损伤*
柳艳丽, 唐震海, 王能里, 林振浪△
(温州医科大学附属育英儿童医院新生儿科, 浙江 温州 325027)
目的: 研究腺苷A2A受体拮抗剂SCH58261减轻成熟胎兔缺氧缺血性脑损伤的作用。方法:选择孕29 d的普通级新西兰孕白兔随机分为假手术组(SO组)、缺氧缺血组(HI组)、SCH58261 0.04 mg/kg剂量组、SCH58261 0.12 mg/kg剂量组和药物溶剂组(DMSO组)。制备胎兔缺氧缺血性脑损伤模型,孕兔均在术后24 h行剖宫产,将存活新生兔置入已提前预热为35 ℃的婴儿暖箱内保暖。记录新生兔的一般状况;评估存活新生兔神经行为学改变;利用质谱法测孕兔血清、新生兔血清及脑组织的SCH58261浓度;采用real-time PCR检测新生兔脑组织皮质、海马和纹状体区Bcl-2和Bax的mRNA表达;利用Western blot实验检测新生兔脑组织皮质、海马和纹状体区p-P38 MAPK的蛋白水平。结果:新生兔血清和脑组织中均能检测到SCH58261的存在。与HI组比较,SCH 0.04 mg/kg组和SCH 0.12 mg/kg组新生兔脑皮质、海马和纹状体区Bcl-2 的mRNA表达均增加(P<0.05),而Bax的mRNA的表达均减少(P<0.05)。将p-P38 MAPK在新生兔脑皮质、海马和纹状体区的表达水平进行比较,SCH 0.04 mg/kg组和SCH 0.12 mg/kg组的P38 MAPK磷酸化水平均低于HI组(P<0.05),且SCH 0.12 mg/kg组与SCH 0.04 mg/kg组相比稍降低(P<0.05)。结论:腺苷A2A受体拮抗剂SCH58261可能通过阻断腺苷A2A受体,抑制神经元P38 MAPK的磷酸化,进一步抑制脑细胞凋亡而减轻宫内缺氧缺血性脑损伤。
A2A腺苷受体拮抗剂; 缺氧缺血性脑损伤; 围产期
新生儿缺氧缺血性脑病(hypoxic-ischemic encephalopathy, HIE)是指由于围生期窒息缺氧引起脑组织缺氧缺血性损害,在缺氧缺血性脑损伤(hypoxic-ischemic brain damage, HIBD)的病因中,由宫内窒息引起者占50%,娩出过程中窒息占40%,先天疾病所致者占10%[1]。然而对于HIE,目前仍缺乏极其有效的治疗措施,亚低温被认为是目前唯一有效的治疗方法;而临床研究表明仅在生后6 h内开始使用亚低温对中度HIBD有效[2]。本研究前期已经建立近足月胎兔持续宫内缺氧缺血性脑损伤模型,其通过暂时性阻断孕兔子宫血供造成胎兔宫内缺氧缺血,很好地模拟了宫内窒息的临床过程,属胎儿期全身性缺氧缺血性脑损伤动物模型[3]。新生儿脑损伤包括线粒体损伤、细胞凋亡及第三阶段损伤,其中线粒体功能损伤继发细胞凋亡被认为在未成熟脑神经细胞死亡中起主导作用[4]。而Bcl-2、Bax在HIBD对未成熟脑后皮质、海马及纹状体区神经细胞凋亡的进程中扮演重要角色[5]。当脑缺血发生后,神经元细胞A2A受体表达水平明显提高[6],细胞外腺苷浓度水平显著提高,升高的腺苷作为一种重要的神经调质,与腺苷受体(adenosine receptor,AR)A1AR、A2AAR、A2BAR及A3AR结合,特别是内源性A2AAR的激活,主要通过早期刺激谷氨酸释放、促进神经系统炎症因子如COX-2、PGE2等的释放,及负性调控NF-κB 路径和P38促分裂原活化蛋白激酶(P38 mitogen-activated protein kinase,P38 MAPK)路径,加重缺血性脑损伤[7]。本研究利用新西兰孕白兔建立胎兔宫内缺氧缺血性脑损伤模型,在缺氧缺血后即刻,给予A2A腺苷受体拮抗剂SCH58261(SCH),观察胎兔脑组织凋亡基因的表达及P38 MAPK磷酸化的改变,为临床HIE防治提供新方法。
材 料 和 方 法
1 分组及动物模型制备
孕29 d健康新西兰白兔23只,体重为3.7~4.5 kg,由上海实验动物中心提供。实验动物许可证编号为SCXK(沪)2007-0005。饲养条件为室温20℃,相对湿度40%~60%。其中20只按照随机数字表法分成假手术(sham operated,SO)组、缺氧缺血(hypoxic-ischemic,HI)组、SCH 0.04 mg/kg组、SCH 0.12 mg/kg组和药物溶剂组(DMSO组),每组4只。另外3只普通级新西兰孕白兔,缺氧缺血后即刻予SCH58261 0.04 mg/kg用于检测孕兔血清、新生兔血清和脑组织的SCH58261浓度。采用全身麻醉和腰麻的方法对孕兔进行麻醉,全身麻醉诱导阶段混合溶液速度为7.5 mL·kg-1·h-1,其中枸橼酸芬太尼为0.075 mg·kg-1·h-1,氟哌利多为3.75 mg·kg-1·h-1,用生理盐水混合后沿耳缘静脉泵入;全身麻醉维持阶段混合溶液速度为2.5~5 mL·kg-1·h-1,枸橼酸芬太尼0.025~0.05 mg·kg-1·h-1,氟哌利多1.25~2.5 mg·kg-1·h-1;腰麻为向蛛网膜下腔内注入0.25%盐酸布比卡因0.3~0.5 mL,当孕兔后肢完全放松表示麻醉成功。从右侧股动脉置入动脉导管进行有创血压监测,监测孕兔实验过程中的心率、血压、氧饱和度等生命体征。从左侧股动脉插入4F Fogarty 动脉取栓导管,插入深度约10 cm,至降主动脉与子宫动脉分支处,即肾动脉开口下端、子宫动脉上端。HI组、SCH 0.04 mg/kg组、SCH 0.12 mg/kg组及DMSO组的动物向导管球囊内注入0.3 mL生理盐水扩张球囊,完全阻断子宫血供造成胎兔缺血,持续25 min,造成胎兔宫内缺氧缺血;SO组不扩张球囊,留置导管25 min。股动脉阻断时间结束即刻,实验组即通过耳缘静脉注射提前几分钟配制好的2 mL药物(SCH58261或溶剂DMSO)。SCH58261溶于10% DMSO,按剂量给药, DMSO组给予10% DMSO 2 mL,HI组不给任何试剂,缝闭切口,局部消毒,返回兔笼。部分实验过程图片见图1。

Figure 1.Photos in the process of experiments. A:NS (0.3 mL) was injected into the 4F artery catheter which was in the left femoral artery. The blood pressure was completely detectable. B: slow withdrawal of the NS evacuated the sacculus and resulted in the blood pressure recovery to normal.
图1 实验过程照片
2 方法
2.1 实验动物管理和一般情况记录 根据实验分组,均在术后24 h(孕30 d)行剖宫产,将存活新生兔置入已提前预热为35 ℃的婴儿暖箱内保暖。记录一般请况包括单窝产崽数、新生兔的死胎率、出生体重等。
2.2 神经行为学评分 在新生兔生后5 min内进行,本研究重点评估翻正反射,记录10次翻正反射中,能成功从仰卧位翻为俯卧位的次数及得分,并计算平均值。在神经行为学评分完成后经腹腔注射5%水合氯醛后直接断头取脑。在雪花碎冰上铺一个灭菌的弯盘,操作者带无菌手套在弯盘内分离脑组织,分别取双侧额皮质、海马、纹状体,放入2 mL无菌冻存管内,立即放液氮罐暂时保存,当次实验结束时,转移至-80 ℃冰箱保存。
2.3 质谱法测孕兔血清、新生兔血清及脑组织的SCH58261浓度 将3只孕兔在缺氧缺血后即刻耳缘静脉内给予SCH58261 0.04 mg/kg,10 min后剖宫产。3只孕兔共产崽16只,其中死产新生兔4只,剩余12只新生兔取血液和脑组织标本检测SCH58261浓度。将新生兔通过腹腔内注射5%水合氯醛镇静,按每100 g体重给予25 mg水合氯醛。暴露心脏,从心尖区插入黑色头皮针,立即抽取心脏血液。待心脏采血完成后,新生兔即刻断头取脑组织,用生理盐水洗去淤血,保存于液氮罐中。(1) 血液标本:在4 ℃冰箱静置4 h后,置于4 ℃低温离心机中,12 000 r/min离心15 min,取上清淡黄色样液于新的无菌去酶1.5 mL EP管,保存于-80 ℃超低温冰箱中。(2) 脑组织标本:将整个脑组织解冻,称重,置于玻璃组织匀浆器中,按1∶5加入0.1 mol/L HCl并研磨匀浆,移至玻璃管中,加5 mL氯仿,涡旋60 s,室温下400×g离心10 min,离心过程重复3次,最后取下清液转移至无菌去酶1.5 mL的EP管中,于55~58 ℃,去塞,水浴氮气吹干,获得约2 mL左右含SCH58261溶液,保存于-20 ℃低温冰箱中。用100 μL流动相溶解残余物,取20 μL注入质谱仪,按本法条件测定胎兔血、脑的SCH58261浓度。
2.4 实时荧光定量聚合酶链反应(real-time PCR)检测各脑区Bcl-2和Bax的mRNA表达 取各脑区(皮质、海马和纹状体)脑组织80~100 mg,用TRIzol试剂、氯仿和异丙醇提取总RNA。用可见光分光光度计测总 RNA的浓度和纯度,A260/A280为1.8~2.2,浓度为1.5~2.0 g/L。逆转录体系含RNA 2 μg、引物1 μL、去离子水加至12 μL,再加入逆转录酶缓冲液4 μL、逆转录酶1 μL、RNasin 1 μL、dNTP混合液2 μL,将上述总共20 μL逆转录体系充分混匀,行普通PCR先确认标本具有阳性表达后,再用real-time PCR反应体系扩增。引物由上海基康生物技术有限公司合成,Bcl-2(153 bp)的上游引物为5’-GGACGCGAAGTGCTATTGGT-3’,下游引物为5’-TCACGATCTCCCGGTTATCATA-3’;Bax(149 bp)的上游引物为5’-CCCAGCATCCTTCCCACG-3’,下游引物为5’-CCCAAGGAAGACAAGAGCATCA-3’;GAPDH(172 bp)的上游引物为5’-GCAAGTTCAACGGCACAG-3’,下游引物为5’-GCCAGTAGACTCCACGACAT-3’。反应条件为95 ℃ 30 s; 95 ℃5 s、60 ℃ 34 s,共40个循环。运用2-ΔΔCt方法,得出mRNA的相对表达量。最终结果由PCR仪配套软件自动计算。
2.5 Western blot实验测定各组胎兔各脑区p-P38蛋白的水平 取冻存于-80 ℃的各脑区(皮质或海马或纹状体)脑组织约100 mg,并加入400 μL裂解液(含5 μL PMSF)迅速在冰上研磨,将匀浆液冰浴 30 min,使其充分裂解。4 ℃高速低温离心机,12 000 r/min离心30 min,取上清装入1.5 mL的EP管;4 ℃高速低温离心机 12 000 r/min离心5 min,取上清,用BCA蛋白浓度试剂盒测定蛋白浓度,各蛋白样本分别加入样本缓冲液调浓度至2 g/L,100 ℃煮10 min进行蛋白变性。制备10%丙稀酰胺分离胶和5%浓缩胶进行蛋白分离(每泳道20 μL),预先将PVDF膜浸泡甲醇数秒,用于转印蛋白,5%脱脂牛奶封闭1 h,分别在不同孵育小盒中加入抗P38抗体(1∶1 000)、抗p-P38抗体(1∶1 000)4 ℃孵育过夜(约16~18 h); 分别加入相应的Ⅱ抗室温孵育1.5 h。TBST洗膜,最后于暗室曝光,将胶片扫描至电脑中,用 Quantity One 凝胶图像分析软件分析P38和p-P38的吸光度。
3 统计学处理
所有数据采用SPSS 17.0软件进行统计分析。采用W检验判断资料是否为正态分布。正态分布的计量资料用均数±标准差(mean±SD)表示。采用完全随机设计的单因素方差分析,两两比较采用SNK-q检验。以P<0.05为差异有统计学意义。
结 果
1 孕兔及新生兔的一般情况
各组孕兔的体重、单窝产崽数和新生兔的出生体重差异均无统计学显著性;SCH 0.04 mg/kg组和SCH 0.12 mg/kg组孕兔的死胎率明显低于HI组,差异有统计学显著性(P<0.05);SCH 0.04 mg/kg组和SCH 0.12 mg/kg组孕兔的死胎率较DMSO组明显下降,差异有统计学显著性(P<0.05),见表1。
2 新生兔行为学评估
新生兔的翻正反射能力可作为神经行为学评估结果。SCH 0.04 mg/kg组和SCH 0.12 mg/kg组绝大多数新生兔可维持俯卧位姿势,有前向运动或圆周运动,较快速地从仰卧位翻正为俯卧位,翻正反射得分的均值在2分左右,吞咽协调功能和对酒精嗅觉刺激的反应能力良好。HI组存活的新生兔出现不同程度的神经行为学异常表现,较多不能维持正常的俯卧位姿势,肢体常常偏向一侧;往往出现翻身困难,10次翻身反射中,成功从仰卧位翻正为俯卧位的次数明显减少,翻正反射得分低于SCH 0.04 mg/kg组和SCH 0.12 mg/kg组,差异有统计学显著性(P<0.05)。但SCH 0.04 mg/kg组和SCH 0.12 mg/kg组的得分均比SO组低,差异有统计学显著性(P<0.05)。DMSO组翻正反射得分与HI组相比差异无统计学显著性(P>0.05),见表1。
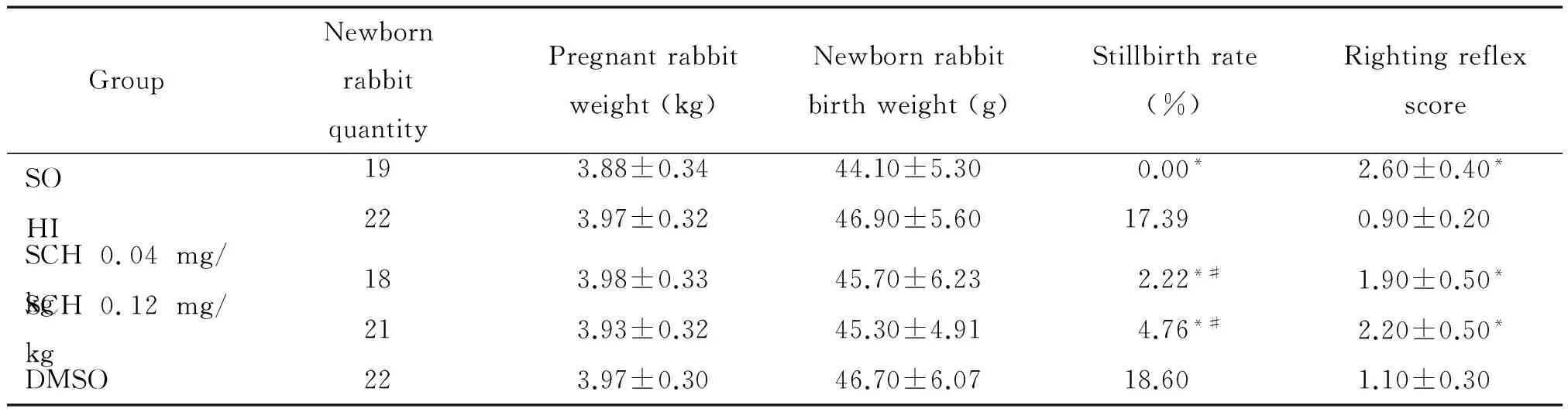
表1 新生兔的一般情况
*P<0.05vsHI group;*P<0.05vsDMSO group.
3 孕兔血清、新生兔血清及脑组织的SCH58261浓度
新生兔血清和脑组织中均能检测到SCH58261的存在,由于将新生兔脑组织按1∶5比例加入0.1 mol/L HCl制成约1 mL脑匀浆,再加5 mL氯仿萃取约2 mL含SCH58261溶液,此时,脑组织SCH58261浓度被稀释约2倍,因此,新生兔脑组织SCH58261浓度应为新生兔脑组织萃取液SCH58261浓度的2倍左右,平均浓度为40 μg/L左右。新生兔血清SCH58261的浓度为(81.5±23.1) μg/L,约为孕兔血清SCH58261浓度[(816.8±135.1) μg/L]的1/10,新生兔实际脑组织SCH58261的浓度为(21.2±6.2) μg/L,约为新生兔血清SCH58261浓度的1/2。其中一组新生兔脑组织SCH58261的浓度测定结果见图2。
4 皮质、海马及纹状体区Bcl-2和Bax的mRNA表达情况
Real-time PCR结果显示,SCH 0.04 mg/kg组和SCH 0.12 mg/kg组皮质、海马和纹状体Bcl-2的mRNA表达均高于HI组(P<0.05),DMSO组虽然与HI组相比略有增加,但差异无统计学显著性(P>0.05);SCH 0.12 mg/kg组较SCH 0.04 mg/kg组Bcl-2的mRNA表达增加(P<0.05),见表2。
SCH 0.04 mg/kg组、SCH 0.12 mg/kg组皮质、海马、纹状体Bax的mRNA表达低于HI组(P<0.05),SCH 0.12 mg/kg组较SCH 0.04 mg/kg组Bax的mRNA表达减少(P<0.05),见表2。
5 皮质、海马及纹状体区p-P38的蛋白水平
SCH 0.04 mg/kg组和SCH 0.12 mg/kg组皮质、海马及纹状体区p-P38 MAPK/P38 MAPK的比值均小于HI组(P<0.05),DMSO组与HI组相比略有减小,但差异无统计学显著性;SCH 0.12 mg/kg组皮质区p-P38 MAPK/P38 MAPK的比值较DMSO组减小(P<0.05),且与SCH 0.04 mg/kg组相比稍减少(P<0.05);与SO组皮质区相比,SCH 0.12 mg/kg组p-p38 MAPK/p38 MAPK的比值仍较大(P<0.05),见图3。
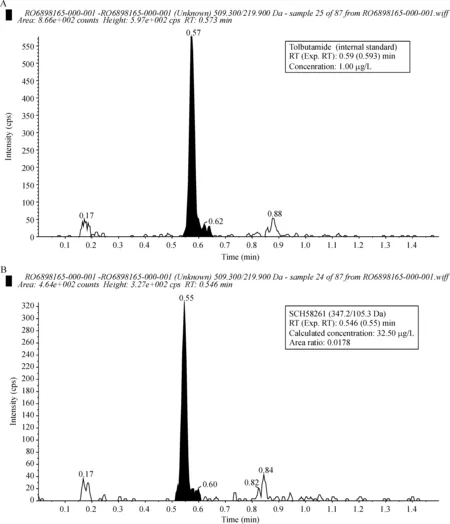
Figure 2.Determination of SCH58261 concentration in the brain tissue of newborn rabbits. A: the figure showed that the concentration sensitivity of SCH58261 was 1 μg/L; B: the figure showed that SCH58261 concentration in the brain tissue of the newborn rabbits was measured.
图2 新生兔脑组织萃取液测得的SCH58261浓度

表2 皮质、海马及纹状体Bcl-2和 Bax的mRNA表达
*P<0.05vsHI group;#P<0.05vsSCH 0.04 mg/kg group.

Figure 3.The protein level of p-P38 MAPK in the cortex (A), hippocampus (B) and striatum (C) of the newborn rabbits determined by Western blot. Mean±SD.n=8.*P<0.05vsHI group;▲P<0.05vsSCH 0.04 mg/kg group.
图3 皮质区、海马区及纹状体区p-P38的蛋白水平
讨 论
缺氧、缺血是新生儿脑损伤的常见原因,目前临床上尚无特效治疗方法,而其较高的致死率和致残率又决定了寻求新的治疗策略具有重要的临床意义。新生动物脑组织发育不成熟,对缺氧缺血刺激的病理改变与成年动物不尽相同。Bcl-2和Bax在未成熟脑HIBD后神经细胞凋亡中扮演重要角色[8],而P38 MAPK磷酸化后可通过多种途径介导细胞凋亡[9]。此外,研究证实MAPK家族,特别是P38 MAPK,通过阻断IL-1β介导的炎症反应而发挥至关重要的作用[10]。
本研究通过左侧股动脉向4F Fogarty动脉取栓导管球囊内注入0.3 mL生理盐水,经右侧股动脉连接生命监测仪,观察到右侧股动脉血压监测不到,说明已完全阻断子宫血供,成功造成宫内缺氧缺血性脑损伤模型,当抽空球囊后,右侧股动脉恢复血供,血压恢复阻断前水平,提示此过程中孕兔模拟出人的急性胎盘功能不全的临床过程。
目前国内外使用宫内窒息模型的研究尚少,因此未发现关于SCH58261能否通过胎盘屏障的报道。本实验通过孕兔耳缘静脉内单次给予SCH58261,首先需证实SCH58261能通过胎盘屏障从而经过胎兔血脑屏障而到达胎兔脑组织。目前尚未发现利用高效液相色谱法或质谱分析技术直接测定脑组织中腺苷A2A受体拮抗剂SCH58261的报道,本研究查阅相关文献,根据Xie等[11]的研究,利用SCH58261是咖啡因的衍生物,与其有相似的分子结构,参考其提取小鼠脑组织中咖啡因的具体方法,提取新生兔脑组织和血清SCH58261,通过质谱分析技术测得标本SCH58261在新生兔血液和脑组织的浓度,证实其能通过胎盘屏障和血脑屏障,从而保障了本实验的进一步研究结果的可靠性。
另外,成年小鼠的脑缺血研究中,通过腹腔单次给SCH58261 0.01 mg/kg,本研究的给药剂量根据动物的体表面积换算成基础给药剂量0.004 mg/kg,并且最终为此给药剂量10倍和30倍即0.04 mg/kg和0.12 mg/kg,通过孕兔耳缘静脉注射,此方法便捷,并且对胎兔的创伤性小。本研究HI后通过孕兔耳缘静脉注射SCH58261 0.04 mg/kg,得到新生脑组织SCH58261浓度约为40 μg/L,推测SCH58261在新生兔脑组织中已达到起效浓度。
缺氧缺血性脑损伤的发病机制涉及多个环节,细胞凋亡被认为在未成熟脑神经细胞死亡中起主导作用,而Bcl-2和Bax在凋亡途径中起着重要作用,Bcl-2通过阻止线粒体细胞色素C的释放而发挥抗凋亡作用,凋亡的细胞又使Bcl-2 的mRNA表达量明显下降。脑损伤发生后,脑细胞ERK水平急剧升高,NF-κB磷酸化,P38 MAPK过度磷酸化[12]。P38 MAPK磷酸化后可作为上游因子通过多种途径如接受信号刺激引起线粒体膜的通透性增加,Bax从胞质向线粒体的转移而介导细胞凋亡等[13]。
本研究通过建立胎兔宫内缺氧缺血性脑损伤模型后,利用real-time PCR和 Western blot实验检测胎兔脑组织匀浆发现,HI组与假手术组相比,HI组新生兔脑组织皮质、海马及纹状体区的P38 MAPK磷酸化水平均明显升高,Bcl-2 的mRNA表达均出现下调,Bax的mRNA表达均呈上调状态,提示Bcl-2和Bax在未成熟脑HIBD后皮质、海马及纹状体区神经细胞凋亡中起着重要作用。P38 MAPK信号通路在缺氧缺血的应激下,可能作为上游因子,诱导细胞凋亡的发生,提示其在脑缺血性损伤中发挥重要作用。
腺苷A2A受体在中枢神经系统主要分布在纹状体,在皮质和海马也有分布;星形胶质细胞、神经元、小胶质细胞和少突胶质细胞均表达A2A受体[14]。小胶质细胞是中枢神经系统固有的免疫效应细胞,在神经系统炎症反应中发挥重要作用。P38 MAPK磷酸化是小胶质细胞活化的关键步骤,而腺苷A2A受体可能通过引起P38 MAPK磷酸化,而加重脑损伤[15]。Silva等[16]研究证实腺苷A2A受体直接影响一系列的有害刺激,促进神经元的凋亡,认为阻断腺苷A2A受体能够减少海马区神经元的凋亡。
由于处于发育期的脑组织与成熟脑组织具有十分明显的差异,且胎儿在子宫内处于一个相对缺氧的环境中,近年来关于腺苷A2A受体在很多HIBD模型中发挥作用的作用存在争议。Ådén等[17]将7日龄的新生小鼠腺苷A2A受体基因敲除后进行单侧颈总动脉结扎造成缺氧缺血性脑损伤,腺苷A2A受体基因敲除小鼠脑损伤和运动异常率比对照组反而增高,提示敲除腺苷A2A受体基因后加重脑损伤[17],推断若阻断新生小鼠腺苷A2A受体可能会加重脑损伤,这与成年动物研究发现的使用腺苷A2A受体拮抗剂能够阻止脑损伤后的神经元坏死的结果刚好相反[18]。
本研究证实了HIBD后给予SCH58261,各脑区如皮质、海马及纹状体区Bcl-2的mRNA表达均出现上调,Bax的mRNA表达均呈下调状态,说明p-P38 MAPK的减少具有抑制细胞凋亡的作用,且real-time PCR和 Western blot实验结果显示,与HI组相比,SCH 0.04 mg/kg组与SCH 0.12 mg/kg组各指标有改善,提示宫内HIBD后予SCH58261 0.04 mg/kg能减轻围产期HIBD,增加剂量至0.12 mg/kg,其神经保护作用较0.04 mg/kg更明显,是否再增加剂量而保护作用呈上升趋势有待于进一步实验考究。
[1] 段 涛, 陈 超. 新生儿缺血缺氧性脑病(VI) [J]. 中华医学杂志, 2005, 85(21):1508-1509.
[2] Lin ZL, Yu HM, Lin J, et al. Mild hypothermia via selective head cooling as neuroprotective therapy in term neonates with perinatal asphyxia: an experience from a single neonatal intensive care unit[J]. J Perinatol, 2006, 26(3):180-184.
[3] 王能里, 南 燕, 柳艳丽, 等. 近足月胎兔持续宫内缺氧缺血性脑损伤模型的建立[J]. 中国病理生理杂志, 2012, 28(4):760-764, 768.
[4] Thornton C, Rousset CI, Kichev A, et al. Molecular mechanisms of neonatal brain injury[J]. Neurol Res Int, 2012, 2012:506320.
[5] Chipuk JE, Green DR. How do BCL-2 proteins induce mitochondrial outer membrane permeabilization?[J]. Trends Cell Biol, 2008, 18(4):157-164.
[6] Paterniti I, Melani A, Cipriani S, et al. Selective adenosine A2Areceptor agonists and antagonists protect against spinal cord injury through peripheral and central effects[J]. J Neuroinflammation, 2011, 8:31.
[7] Mohamed RA, Agha AM, Nassar NN. SCH58261 the selective adenosine A2Areceptor blocker modulates ischemia reperfusion injury following bilateral carotid occlusion: role of inflammatory mediators[J]. Neurochem Res, 2012, 37(3):538-547.
[8] Gibson ME, Byung HH, Choi J, et al. BAX contributes to apoptotic-like death following neonatal hypoxia-ischemia: evidence for distinct apoptosis pathways[J]. Mol Med, 2001, 7(9):644-655.
[9] Morillo SM, Abanto EP, Román MJ, et al. Nerve growth factor-induced cell cycle reentry in newborn neurons is triggered by p38MAPK-dependent E2F4 phosphorylation[J]. Mol Cell Biol, 2012, 32(14): 2722-2737.
[10]Zhang R, Sun L, Hayashi Y, et al. Acute p38-mediated inhibition of NMDA-induced outward currents in hippo-campal CA1 neurons by interleukin-1β[J]. Neurobiol Dis, 2010, 38(1):68-77.
[11]Xie XB, Mhaskar Y, Arbogast YA, et al. Adenosine receptor antagonists and behavioral activation in NF-κB p50 subunit knockout mice[J]. Life Sci, 2009, 85(5-6): 226-234 .
[12]Guo RB, Wang GF, Zhao AP, et al. Paeoniflorin protects against ischemia-induced brain damages in rats via inhibiting MAPKs/NF-κB-mediated inflammatory responses[J]. PLoS One, 2012, 7(11):e49701.
[13]Chen ZL, Jiang H, Wan YW, et al. H2O2-induced secretion of tumor necrosis factor-α evokes apoptosis of cardiac myocytes through reactive oxygen species-dependent activation of p38 MAPK[J]. Cytotechnology, 2012, 64(1): 65-73.
[14]Stevens B, Porta S, Haak LL, et al. Adenosine: a neuron-glial transmitter promoting myelination in the CNS in response to action potentials[J]. Neuron, 2002, 36(5): 855-868.
[15]Neumann J, Sauerzweig S, Ronicke R, et al. Microglia cells protect neurons by direct engulfment of invading neutrophil granulocytes: a new mechanism of CNS immune privilege[J]. J Neurosci, 2008, 28(23):5965-5975.
[16]Silva CG, Porciúncula LO, Canas PM, et al. Blockade of adenosine A2Areceptors prevents staurosporine-induced apoptosis of rat hippocampal neurons[J]. Neurobiol Dis, 2007, 27(2):182-189.
[17]Ådén U, Halldner L, Lagercrantz H, et al. Aggravated brain damage after hypoxic ischemia in immature adenosine A2Aknockout mice[J]. Stroke, 2003, 34(3):739-744.
[18]Gomes CV, Kaster MP, Tomé AR, et al. Adenosine receptors and brain diseases: neuroprotection and neurodegeneration[J]. Biochim Biophys Acta, 2011, 1808(5):1380-1399.
(责任编辑: 林白霜, 罗 森)
Adenosine A2Areceptor antagonist SCH58261 attenuates hypoxic-ischemic brain damage in a fetal rabbit model
LIU Yan-li, TANG Zhen-hai, WANG Neng-li, LIN Zhen-lang
(DepartmentofNeonatology,YuyingChildren’sHospitalofWenzhouMedicalUniversity,Wenzhou325027,China.E-mail:linzhenlang@hotmail.com)
AIM: To study the effect of adenosine A2Areceptor antagonist SCH58261 on hypoxic-ischemic brain damage (HIBD) in a mature fetal rabbit model.METHODS: Pregnant New Zealand white rabbits at gestational day 29 were selected and were randomly divided into sham-operated group, hypoxic-ischemic group, SCH58261 0.04 mg/kg group, SCH58261 0.12 mg/kg group and DMSO group. The intrauterine rabbit HIBD model was established. All pregnant rabbits were subjected to cesarean section 24 h after the sham operation or experimental procedure to induce hypoxic-ischemic injury in the fetus. The survival neonatal rabbits were kept in a neonatal incubator at 35 ℃. The general conditions of the newborn rabbits were recorded. The degree of neurobehavioral damage in the newborn rabbits was estimated by a neurobehavioral scoring protocol. The concentration of SCH58261 in the serum of pregnant rabbits, the serum of neonatal rabbits and the brain tissues of neonatal rabbits was measured by mass spectrometry. The mRNA expression of Bcl-2/Bax and protein levels of p-P38 mitogen-activated protein kinase (MAPK) in the cortex, hippocampus and striatum area in the brain of the neonatal rabbits were determined by real-time PCR and Western blot. RESULTS: SCH58261 was detected in the serum and brain tissues of the newborn rabbits. The SCH58261 concentration was approximately 40 μg/L in the brain tissue of the newborn rabbits. The mRNA expression of Bcl-2 in the cortex, hippocampus and striatum of brain tissues in SCH58261 0.04 mg/kg group and SCH58261 0.12 mg/kg group was higher, and the mRNA expression of Bax was lower than those in HI group (P<0.05). The protein level of p-P38 MAPK in the cortex, hippocampus and striatum of brain tissues was reduced in SCH58261 0.04 mg/kg group and SCH58261 0.12 mg/kg group compared with HI group (P<0.05). The protein level of p-P38 MAPK in SCH58261 0.12 mg/kg group was a little lower than that in SCH58261 0.04 mg/kg group (P<0.05). CONCLUSION: Adenosine A2Areceptor antagonist SCH58261 attenuates hypoxia-ischemia induced neonatal brain injury by blocking adenosine A2Areceptor, subsequently inhibiting p-P38 MAPK phosphorylation to reduce neuronal apoptosis.
Adenosine A2Areceptor antagonist; Hypoxic-ischemic brain damage; Peripartum
1000- 4718(2016)12- 2139- 08
2016- 06- 18
2016- 09- 06
卫生部新生儿疾病重点实验室开放课题
R722; R363
A
10.3969/j.issn.1000- 4718.2016.12.004
杂志网址: http://www.cjpp.net
△通讯作者 Tel: 0577-88002798; E-mail: linzhenlang@hotmail.com
