水稻对褐飞虱的抗性鉴定及其抗性机制研究
2016-12-29薛艳霞覃宝祥邱永福李容柏
陈 曙,刘 芳,2,3,薛艳霞,祝 亚,覃宝祥,2,3,韦 政,2,3,邱永福,2,3*,李容柏,2,3*
(1.广西大学农学院,广西 南宁 530005;2.广西大学亚热带农业生物资源保护与利用国家重点实验室,广西 南宁 530005;3.广西大学植物遗传育种重点实验室,广西 南宁 530005)
水稻对褐飞虱的抗性鉴定及其抗性机制研究
陈 曙1,刘 芳1,2,3,薛艳霞1,祝 亚1,覃宝祥1,2,3,韦 政1,2,3,邱永福1,2,3*,李容柏1,2,3*
(1.广西大学农学院,广西 南宁 530005;2.广西大学亚热带农业生物资源保护与利用国家重点实验室,广西 南宁 530005;3.广西大学植物遗传育种重点实验室,广西 南宁 530005)
为了研究水稻品种在不同时期对褐飞虱的抗性变化及其抗性机制,对15份水稻材料进行苗期和成株期抗虫鉴定。并通过测定水稻抗生性及趋避性,对这15份水稻材料进行抗性机制研究。结果表明,15份水稻材料有13份在苗期和成株期的抗性没有或呈现微弱变化,其余2份材料T12、OB667在苗期抗性鉴定为感虫,在成株期表现为中抗。在抗生性试验下,其抗生性大小与抗虫能力基本保持一致;而在趋避性试验中,水稻的趋避性强弱并未与其抗虫性强弱表现一致性。
水稻;稻褐飞虱;成株期;抗生性;趋避性
稻褐飞虱(Nilaparvatalugens)是水稻的主要害虫之一,主要从刺吸、产卵和传病三个方面对水稻造成危害[1-2]。利用抗性品种是有效防治褐飞虱危害的措施之一,由于不少品种在苗期检测中不表现出抗性,但在田间试验中会表现出中抗或抗稻褐飞虱,若仅仅使用苗期集团鉴定法,能够将一些高抗性的品种鉴定出来,一些中等抗性或耐虫性的品种却因苗期表现不抗或弱抗性而没被筛选甚至淘汰是非常可惜的[3]。要准确地鉴定出水稻对褐飞虱的抗性强弱,不仅需要鉴定水稻苗期抗性,还需研究水稻的成株期抗性以及水稻对褐飞虱的抗性机制,因此成株期抗性鉴定和水稻抗性机制的研究对育种或品种的推广具有实用的意义。目前,许多学者对水稻关于褐飞虱抗性的研究主要集中在改进水稻的抗性检测方法上。刘光杰等[4]通过苗期鉴定法和分蘖盛期接虫鉴定法检测稻飞虱对不同品种的抗性反应,结果表明,苗期鉴定法能够鉴定大部分水稻品种对稻飞虱的抗性水平,但对于一些应用价值高的品种,必须还要进行分蘖盛期抗性检测才能准确、全面评价其抗性水平。陈建明等[1]利用改进的苗期集团筛选法,对国家和浙江省水稻育种攻关项目的769个水稻新品种进行褐飞虱的抗性检测,发现利用水稻苗期和成株期的受害级别和持抗期长短鉴定耐褐飞虱品种的抗性是一种比较可行的方法。陈英之等[5]采用苗期群体筛选鉴定、分蘖期单株鉴定和大田自然诱虫鉴定3种方法,分别测定5个新育成的优质高产新品系对稻褐飞虱的抗性反应,结果显示,在苗期接虫鉴定情况下的抗性表现不稳定,但随着苗龄的增大,其抗虫性有不同程度地提高,分蘖期和抽穗期鉴定更能准确地反映水稻品种成株期的抗性水平。目前对水稻的抗性研究主要集中在对水稻抗性鉴定方法的改良之上,但苗期与成株期对稻飞虱的抗性关系尚不清楚,对水稻抗性机制的研究了解尚不深入。本研究结合水稻品种苗期和成株期的抗性鉴定结果,对供试品种分别进行抗性机制研究,评价15份代表性水稻抗性材料对褐飞虱的抗性作用,为后续抗稻飞虱的水稻材料的筛选、推广和生产应用提供参考依据。
1 材料与方法
1.1 供试材料
选用15份不同来源含不同抗稻褐飞虱基因的材料,分别是Mudgo(Bph1)、 ASD7(Bph2)、 Rathu Heenati(Bph3)、 Babawee(bph4)、 ARC10550(bph5)、 Swarnalata(Bph6)、T12(bph7)、Chin Saba (bph8)、Pokkali(Bph9);TN1作为感虫对照品种,PTB33作为抗虫对照品种。水稻材料OB677,ARC5984为广西农科院水稻研究所提供,BPH660、08BPH327为广西大学农学院野生稻课题组近几年育成的水稻抗性鉴定品种。
1.2 虫源
供试的虫源采集自南宁市郊的稻田,主要为混合生物型,在26~30 ℃条件下,在养虫室内用品种TN1来饲养繁殖,当采集的褐飞虱种群繁殖1~2代后,取其1~2龄若虫用于抗性鉴定。
1.3 水稻材料对褐飞虱的抗性
1.3.1 苗期群体的抗性鉴定 使用58 cm×38cm×10 cm的托盘,加入2 cm厚的水稻田泥,取各水稻品种的种子播于其中,每个品种播1行,随机排列,重复3次。每盆加入感性对照TN1和抗性对照PTB33,行间距3 cm。当幼苗长至3叶期,剔除其中弱苗,每行留下20株生长状态均匀一致的壮苗。按平均每株5头接入1~2龄稻褐飞虱若虫,并用透光性良好的纱网笼将其罩住防止虫飞走。当感虫对照TN1全部死苗时调查各材料幼苗的受害情况,并进行统计,参照巫国瑞[5]、邱永福[7]等的方法,根据死苗率评价抗性等级(表1)。
1.3.2 苗期单株的抗性鉴定 将发芽的种子分别播于小塑料盆钵中,每钵1苗。待秧苗长到3叶期时,按每苗接入8头1~2龄的若虫,并罩上透明的塑料罩,每个品种各设15个重复。当感虫品种TN1苗全部死亡时,根据其死亡率来评价各材料的抗性级别(表1)。

表1 水稻苗期对褐飞虱的抗性评价标准

表2 水稻材料成株期抗褐飞虱鉴定评价标准
1.3.3 水稻成株期单株抗性鉴定 取播种后约40 d的健壮苗,去掉次生分蘖,保留主茎并分别将其移栽于小塑料盆钵中,按每株接入10头3~4龄的褐飞虱若虫,并罩上透明的塑料罩,每个品种设置10个独立重复。结合刘光杰[3]提出的分蘖盛期抗性鉴定方法,当感虫对照品种TN1秧苗全部死亡时统计各材料的死苗情况(表2)。
1.4 抗生性试验
1.4.1 褐飞虱成虫蜜露分泌量的测定 选用15份供试水稻材料播种后40 d的健壮苗,去掉次生分蘖,保留主茎,将封口薄膜剪成2.0 cm×3.5 cm的小片,折成小袋固定在主茎第一或第二叶鞘的顶部,每株固定一个小袋,同时在小袋中接入1头新近羽化的褐飞虱雌虫(经饥饿处理),1个小袋为一个重复,每个品种共设置15个重复。48 h后小心拆下小袋,放出里面的褐飞虱成虫,用万分之一的电子天平称量附着有蜜露的小袋总重,然后用脱脂棉轻擦掉蜜露,称量小袋重量,用总重减去袋重即得到蜜露重量,根据蜜露重量评价材料的抗性水平。
1.4.2 褐飞虱生长速率的测定 取播种后40~45 d左右的健壮苗,去掉次生分蘖,保留主茎,将主茎洗净后移栽于容积为0.4 L的塑料杯中,每杯一苗,上罩一个同样大小、底端有通气孔的塑料罩,从通气孔放入10头预称重的2龄褐飞虱若虫,褐飞虱取食4 d后,将每杯中存活的褐飞虱取出称重,并根据邱永福等[8]提出的褐飞虱生长速率计算方法计算其平均生长速率(mg/BPH/d),每份材料分别设置15个重复。褐飞虱生长速率计算方法为:4 d后存活的褐飞虱的平均重量记为Wtotal,最初放入褐飞虱的平均重量记为Worig,利用它们的差值除上取食天数(Number of days),计算所得即为褐飞虱的平均生长速率。
1.4.3 褐飞虱存活率和羽化率的测定 褐飞虱成活率的测定与生长速率测定的方法类似,将主茎移栽于容积为0.4 L的塑料杯后,罩一个同样大小、底端有通气孔的塑料罩,按每株放入10头2龄的褐飞虱若虫。在接虫后第6天统计褐飞虱的存活情况。同理,羽化率的测定方法与成活率一样,在接虫后第9 d统计褐飞虱的羽化情况。每份材料分别设置10个重复。
1.5 趋避性试验
水稻材料的处理同上述类似,取40 d左右的健壮苗,保留主茎。每个材料设15个重复。将15份材料共计225个单株随机移栽于1.5 m×1.5 m的鉴定圃内,按平均每株15头2龄褐飞虱于鉴定圃内,并用特制的3 m×3 m(便于进入观察统计)无底纱网笼子套住。分别在接虫后第3、9、24、48、72和96小时观察记录各水稻植株上褐飞虱的着虫数。通过比较不同寄主上附着褐飞虱的数量来评价不同水稻材料对褐飞虱的趋避性。
2 结果与分析
2.1 不同品种在苗期和成株期对稻褐飞虱的抗性鉴定
通过使用苗期群体筛选、苗期单株筛选和成株期单株筛选3种抗性鉴定方法对15份抗性材料进行鉴定,实验结果见表3。Babawee、Pokkali、08BPH327在苗期和成株期都表现出了很强的抗性(抗性值大小≈3);Rathu Heenati、Babawee、ARC5984、Pokkali、Swaralata、08BPH327、 PTB33和BPH660八份材料苗期和成株期抗性水平强弱并没有发生变化,其苗期和成株期的抗性水平保持一致;而ARC10550、T12和OB677三份材料成株期比苗期抗性有显著提高,其中T12在苗期群体筛选和单株筛选法中,其平均抗性大小分别为7.20和7.46,但其成株期抗性平均值却达到了3.75,根据国际水稻研究所公布的水稻对褐飞虱抗性水平评价标准(表1),抗性值越小其抗性水平越高,即T12在成株期抗性检测试验中表现出较强抗性,较苗期而言,抗性大幅增强。
2.2 水稻品种对褐飞虱的抗性机制研究
2.2.1 抗生性试验 从蜜露量的测定结果中可以看出,褐飞虱取食48 h后的蜜露量在1.3 mg(PTB33)到34.0 mg(TN1)之间。在ARC5984、Chin saba和TN1中的蜜露量达到最高,达30.4~34.0 mg;在Pokkali、T12、Babawee、Mudgo、OB667、ARC10550、Swarnalata、ASD7中相对较低,为12.7~25.6 mg;而在Rathu Heenati、BPH660、08BPH327和PTB33中仅为1.3~6.1 mg。很明显在含有不同抗性基因的品种中,褐飞虱取食的蜜露量差异非常显著(表4)。
在生长速率的测定中发现,在TN1上褐飞虱的生长速率最大,达到0.1004 mg/BPH/d,在品种ASD7、ARC5984、Chin saba 、Swarnalata 中褐飞虱的生长速率也相对较大,为0.0784~0.0971 mg/BPH/d,而在PTB33中最低,仅为0.0050 mg/BPH/d(表4)。
褐飞虱的平均存活率范围为31 %~70 %。其中在TN1上的褐飞虱平均存活率最高,达到了70 %;在ARC10550、Chin saba和ASD7上褐飞虱存活率次之(≥60 %),在材料T12、OB677、Mudgo、ARC5984、Swarnalata、08BPH327、Pokkali上的存活率则相对较低(40 %至60 %),而在Babawee(39%)、PTB33(31 %)、BPH660(33 %)上的褐飞虱存活率最低。
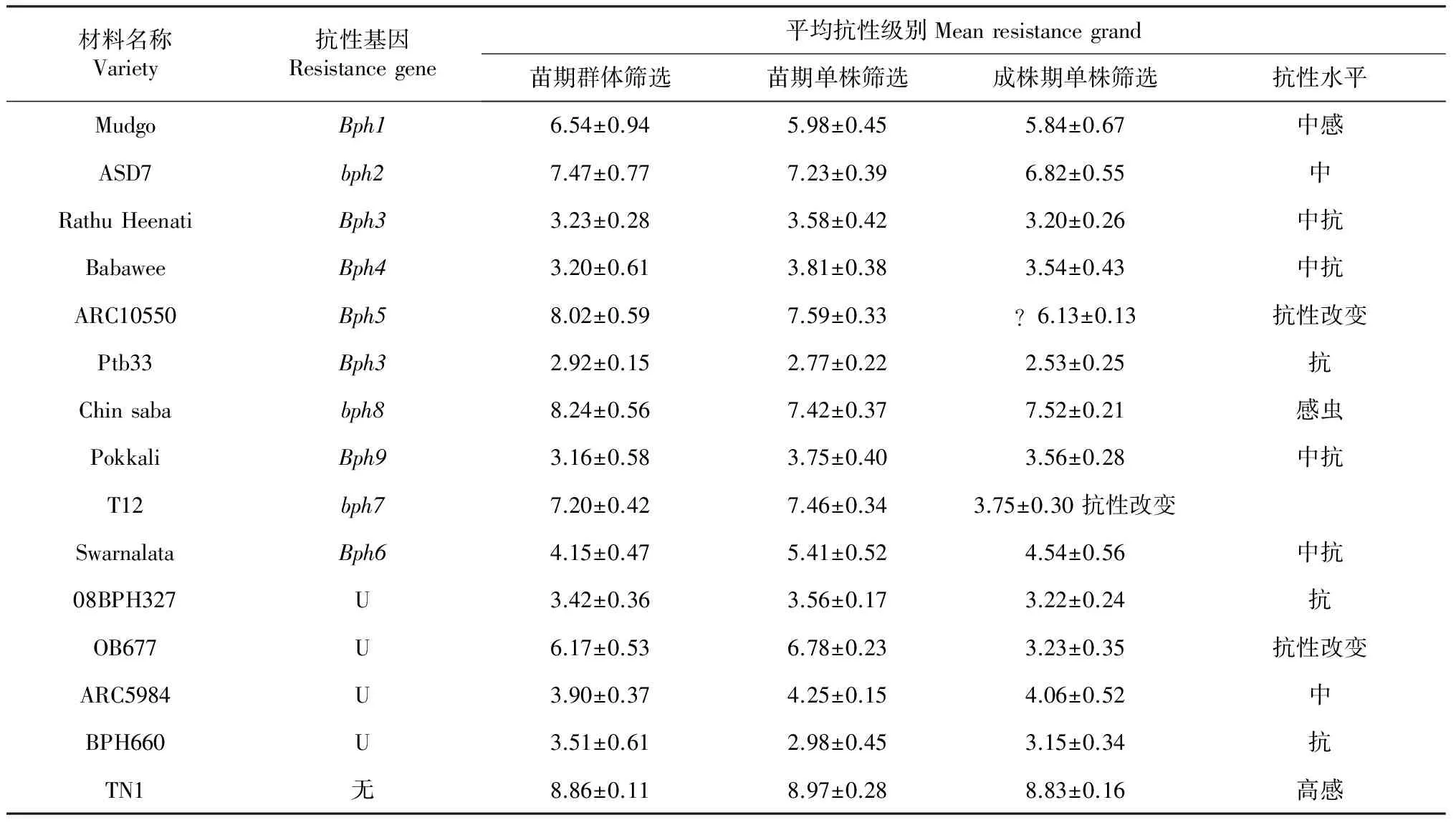
表3 水稻苗期和成株期对褐飞虱的抗性鉴定
褐飞虱羽化率的平均变化范围则为39 %~59 %,在PTB33中褐飞虱羽化率最高,达到59 %;在材料Swarnalata、Babawee、Pokkali、BPH660、08BPH327、Rathu Heenati、OB677中次之(50 %~58 %),在材料Chin saba (41 %)、Swarnalata(41 %)、TN1(39 %)中羽化率则表现出最低水平。结合相关性分析结果(表5),发现相对羽化率而言褐飞虱的存活率更能准确的反应出不同材料的抗性差别。

表4 水稻材料对褐飞虱的抗生性
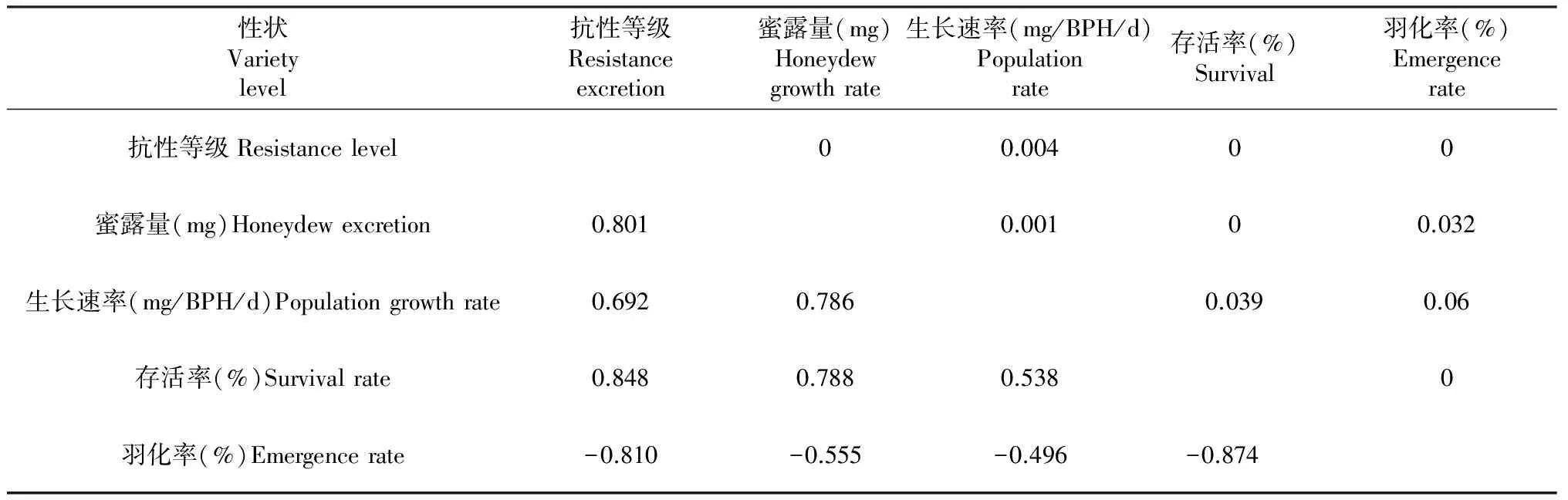
表5 褐飞虱抗生性与水稻抗性强弱的相关性分析
注:左下角是相关系数r,右上角是P值;r0.05=0.514,r0.01=0.641。
Note:The data in the lower left corner of the table are correlation coefficientrand right corner in the top areP;r0.05=0.514,r0.01=0.641.
2.2.2 水稻抗性大小与抗生性分析 为了解褐飞虱取食水稻后的抗生性生理指标变化是否与宿主水稻的抗性强弱有关,我们以实验所测定的15份水稻材料成株期抗性结果作为相应水稻材料的抗性等级,分别与褐飞虱取食这15份水稻材料后的蜜露量、生长速率、存活率以及羽化率测定的结果作相关性分析,结果(表5)表明,宿主水稻材料的抗性强弱与褐飞虱取食水稻后的存活率、羽化和蜜露量密切相关,其相关系数绝对值大小分别为0.848、0.810、0.801,表明选用此三者中的任意一种均能够较准确的反映出宿主水稻材料的抗性强弱;而褐飞虱取食水稻材料后的生长速率的大小与宿主的抗性强弱之间的相关系数为0.692,达到了1 %显著水平。说明对于水稻材料,褐飞虱取食后蜜露量、生长速率、羽化率和存活率均能体现出其抗性强弱,其中褐飞虱的存活率与其相关性最大,最能表现出其抗性水平;羽化率和蜜露量次之,而生长速率与其相关性最差。2.2.3 趋避性试验 如表6所示,在接虫后3 h内,不同水稻材料间褐飞虱的着虫数出现了差异,但由于接虫后时间较短,褐飞虱分布并不均匀,各材料之间差异不明显;随着接虫时间的延长,差异逐渐显著。在第96小时,感性对照TN1上的平均着虫数为26.3只/株,达到最高;水稻材料PTB33、BPH660分别为1.5和2.3只/株,显著小于感性对照TN1上的着虫数,说明水稻材料PTB33和BPH660对褐飞虱有趋避作用;而处于中间的其它13份供试水稻材料在第96小时的着虫数目的范围为7.8~22.3只/株。

表6 水稻材料对褐飞虱的趋避性
3 结论与讨论
3.1 水稻不同时期的抗性鉴定
鉴定水稻品种对褐飞虱抗性的方法有多种,本研究中采用了苗期群体筛选法、苗期单株筛选法、成株期抗性鉴定以及通过褐飞虱蜜露量、存活率和生长速率等方法,测定水稻对褐飞虱的抗性。结果表明:15份试验材料中有13份材料的抗性表现十分接近或者有微小幅度的变化,其余两份材料T12和OB667,T12在苗期和成株期的抗性平均值分别为7.20和3.75,而OB667分别为6.17和3.23,它们的抗性随着秧龄的增大发生了明显的增强,在苗期和成株期的抗性有比较大的变化,与陈建明[1]发现的现象类似。从目前的研究发现,苗期抗稻飞虱的品种其成株期也一定表现出抗性,反之,有的抗性品种在苗期感虫,但在成株期表现出一定的抗性,因此,苗期是进行抗稻飞虱鉴定的敏感度较高时期,一般的抗性育种通过苗期群体筛选法筛选的抗性材料是可靠的。但是在条件具备的情况下,进行成株期抗性鉴定,甚至进行田间抗性检测可更准确、全面的测定目标品种的抗性,提高抗性鉴定的准确性,这对于育种工作中选育抗性品种来说具有重要的意义。
3.2 水稻对褐飞虱抗性机制的研究
为了深入了解水稻对褐飞虱的抗性,我们结合苗期和成株期的抗性鉴定结果,对试验所用的15份水稻材料进行了抗性机制的研究。研究发现,水稻材料PTB33、BPH660在苗期及成株期的抗性鉴定中,对褐飞虱表现为高抗,在对褐飞虱蜜露量、生长速率、羽化率以及存活率的测定结果中表现出了很强的抗生性和趋避性;抗性检测结果发现(表3),Babawee、Pokkali、T12、08BPH327、OB667对褐飞虱有比较强的抗性,这5份材料对褐飞虱蜜露量、生长速率、羽化率和存活率4个测定指标中的变化程度相对于对照组TN1来说要明显大于虫口密度的变化,说明在这5种材料中,其抗生性作用强于趋避性;而Mudgo、ARC5984、Swarnalata对褐飞虱则表现出中等抗性,同时其抗生性和趋避性检测结果均为中等;而材料ARC10550、Chin saba、Rathu Heenati、ASD7的抗性检测在苗期和成株期都为高感,抗性机制为无抗生性和中等趋避性。从研究结果中发现,褐飞虱的存活率与羽化率密切相关,褐飞虱蜜露量小的材料,通常都具有比较低的存活率和生长速率。所以,对褐飞虱具有较高水平抗性的材料,它们的抗性机制主要由抗生性主导,并且其抗性强弱可以通过抗生性来衡量。
[1]陈建明,俞晓平,程家安,等.水稻新品种(系)对褐飞虱抗性的筛选及评价[J].中国水稻科学,2005,19(6):573-576.
[2]邓 飞,倪 深,朱旭东.水稻抗褐飞虱育种研究的现状与展望[J].中国农学通报,2011,27(24):229-237.
[3]黄所生,黄凤宽,吴碧球,等.水稻新品种(组合)对褐飞虱的抗性评价[J].西南农业学报,2014(5):1919-1923.
[4]刘光杰,付志红,沈君辉,等.水稻品种对稻飞虱抗性鉴定方法的比较研究[J].中国水稻科学,2002,16(1):52-56.
[5]陈英之,李树娟,李容柏.水稻对褐飞虱抗性鉴定的比较研究[J].安徽农业科学,2010,38(13):6686-6688.
[6]徐雪亮,肖叶青,陈大洲,等.2个新培育水稻材料对褐飞虱的抗性机制研究[J].中国农学通报,2015,31(10):171-175.
[7]Qiu Y F, Guo J P,Jing S L.Identification of antibiosis and tolerance in rice varieties carrying brown planthopper resistance genes[J].Entomologia Experimentalis et Applicata,2011,10:1570-7458.
[8]Qiu Y F, Guo J P, Jing S L, et al. High-resolution mapping of the brown planthopper resistance geneBph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds[J]. Theoretical and Applied Genetics,2010,121: 1601-1611.
[9]Heinrichs E A, Aquino G B, Chelliah S .Resurgence ofNilaparvatalugens(Staål) populations as influenced by method and timing of insecticide applications in lowland rice[J]. Environmental Entomology,1982,11: 78-84.
[10]Tang M, Lv L, Jing S L, et al. Bacterial symbionts of the brown planthopper,Nilaparvatalugens(Homoptera:Delphacidae)[J]. Applied and Environmental Microbiology, 2010,76(6): 1740-1745.
[11]Kabis A,Khush G S. Genetic analysis of resistance to brown planthopper in riceOryzasativaL.[J]. Plant Breeding,1988, 100:54-58.
(责任编辑 温国泉)
Study on Evaluation and Mechanism of Rice Resistance to Brown Planthopper
CHEN Shu1,LIU Fang1,2,3,XUE Yan-xia1,ZHU Ya1,QIN Bao-xiang1,2,3, WEI Zheng1,2,3,QIU Yong-fu1,2,3,LI Rong-bai1,2,3*
(1.Agricultural College, Guangxi University, Guangxi Nanning 530005,China; 2. State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources, Guangxi Nanning 530005,China; 3.Guangxi Colleges and Universities Key Laboratory of Plant Genetics and Breeding, Guangxi Nanning 530005,China)
In order to evaluate resistance of rice to brown planthopper (BPH) at different growth stage and to study rice resistance mechanisms,We evaluate the resistance of 15 different rice varieties in seedling and adult stages, respectively. And then to reserach the mechanisms for rice to BPH,we tested the antibiosis and antixenosis The result showed that of the 15 rice varieties tested 13 presented not any or little difference in BPH resistance between seedling test and adult stage, while the other two varieties, T12 and OB677, were susceptible in seedling stage and moderate resistance in adult stage. The study on rice resistance mechanism showed that rice antixenosis to the BPH was usually not consistent with its resistance, but rice antibiosis to the insect was highly consistent with its resistance.
Rice;Nilaparvatalugens; Adult stage; Antibiosis; Antixenosis
1001-4829(2016)09-2125-06
10.16213/j.cnki.scjas.2016.09.020
2016-03-09
国家自然科学基金项目(31160276);广西自然科学基金项目(2013GXNSFGA019009);广西科学研究与技术开发项目(桂科重14121001-1-5)
陈 曙(1988-),男,湖北黄冈人,硕士,主要研究方向为水稻遗传育种,*为通讯作者,E-mail:lirongbai@126.com, E-mail:yfqiu@126.com。
S435.112+.3
A
