PDXl与PAX4双基因表达腺病毒载体的构建与表达
2016-12-22霍震沈进许礼发
霍震 沈进 许礼发

摘要: 运用pADxsi系统制备可表达人PDX1与PAX4双基因的重组5型腺病毒。从pEGFP-N1-PDX1质粒上酶切下目的基因PDX1并酶连到腺病毒穿梭质粒pShuttle-EGFP-CMV上,替换EGFP而得到pShuttle-CMV-PDX1;再将PAX4从pEGFP-N1-PAX4质粒上酶切下连到pShuttle-CMV-PDX1的多克隆酶切位点而得到穿梭质粒pShuttle-CMV-PDX1/CMV-PAX4;双酶切pShuttle-CMV-PDX1/CMV-PAX4将CMV-PDX1/CMV-PAX4转移到ADxsi骨架质粒上,得到pADxsi-CMV-PDX1/CMV-PAX4腺病毒质粒,然后在293细胞中进行包装并扩增重组腺病毒,并进行病毒滴度测定;体外感染人间充质干细胞。依据酶切、测序和PCR的结果均证明重组人PDX1与PAX4双基因表达腺病毒载体构建正确;而RT-PCR与WB结果也显示目的基因在细胞中稳定表达。应用重组技术成功构建人PDX1与PAX4双基因表达5型腺病毒载体,转录因子PDX1与PAX4在间充质干细胞内稳定表达且定位于细胞核内。
关键词: 5型腺病毒;间充质干细胞(MSCs);胰腺十二指肠同源框蛋白1(PDX1);成对盒基因4(PAX4);转录因子
中图分类号:Q78文献标志码:A
文章编号:1672-1098(2016)02-0080-07
Abstract:With the use of pADxsi system, the recombinant adenovirus of expressing PDX1 and PAX4 is in place.GFP in the pShuttle-GFP-CMV vector is replaced with the target gene of PDX1 obtained from the pEGFP-N1-PDX1 plasmid, resulting in the pShuttleCMV-PDX1 plasmid. With the pEGFP-N1-PAX4 plasmid cut off, PAX4 is inserted into pShuttle-CMV-PDX1 to acquire the pShuttle-CMV-PDX1/CMV-PAX4 plasmid. Afterwards, the CMV-PDX1/CMV-PAX4 fragment is transferred to the ADxsi skeleton vector to get the pADxsi-CMV-PDX1/CMV-PAX4 virus vector. Finally, the recombinant adenovirus is packaged, amplificated, and titrated in HEK293 cells for the infection of human umbilical cord mesenchymal stem cells. The structure of pADxsi-CMV-PDX1/CMV-PAX4 adenovirus expression vector is confirmed by means of a restriction analysis and PCR. RT-PCR and Western blot results have established that the target genes are persistently expressed in the infected cells.The recombinant human PDX1 and PAX4 expressing adenovirus is successfully constructed, packaged, and amplificated.The target genes can be continuously presented in mesenchymal stem cells.
Key words:adenovims vector;pancreatic and duodenal homeobox factor 1;paired box gene 4;mesenchymal stem cells
糖尿病是由于胰岛β细胞功能绝对或相对缺陷,以慢性高血糖为特征的终身性代谢性疾病[1-2]。长期血糖增高,可导致大血管、微血管受损并危及心、脑、肾、周围神经、眼睛、足等,每年因糖尿病死亡者有一半以上是心脑血管所致,10%是肾病变所致;因糖尿病截肢是非糖尿病的10~20倍[3-4]。因此,预防糖尿病的并发症并最终控制血糖是重要的社会问题。
至目前为止,控制患者血糖仍主要是依赖每天胰岛素的补充,给患者生活带来极大的不便与沉重的经济负担。寻找具有合成与分泌胰岛素的细胞进行细胞替代治疗一直是研究的热点。近年来研究证实间充质干细胞具有多向分化的潜能且无免疫原性,是一类极具应用潜能的干细胞[4-5]。胰十二指肠同源框基因1 (pancreatic and duodenal homeobox factor 1,PDX 1)在胚胎发育过程中具有促进胚胎干细胞向胰腺的早期发育和晚期胰岛素分泌细胞分化的功能[6];此外,还具有维持胰岛β细胞合成与分泌胰岛素的功能[7-8];然而,单独的PDX1转录因子并不能有效诱导干细胞定向向胰岛β细胞分化,在胰腺前体分化与发育过程中,成对盒基因4(paired box gene 4,PAX4)表达将有利于胰腺前体细胞定向向β细胞分化,并参与调控β细胞功能的成熟[8-10]。因此,PDX1与PAX4联合作用对诱导MSCs定向向具有合成与分泌胰岛素功能的β样细胞可能具有促进作用,基于这一假设,在本研究中,选用对MSCs具有较高感染效率的Ⅴ型腺病毒载体,构建携带目的基因PDX1与PAX4的重组腺病毒ADxsi-CMV-PDX1/CMV-PAX4,感染MSCs细胞,以观测所携带的目的基因表达情况,为进一步研究PDX1与PAX4在诱导MSCs向具有合成与分泌胰岛素功能的β样细胞分化的可能及其分子机制奠定实验基础。
1材料和方法
1.1材料与试剂
Bgl II等多种限制性内切酶、Klenow及T4 DNAligase均购自美国Sigma 公司;CIP(Alkaline Phosphatase,Calf Intestinal)酶、质粒提取纯化试剂盒和凝胶回收试剂盒购自北京天恩泽公司;腺病毒pADxsi载体系统由上海汉恒生物有限公司提供;293细胞及DH5a菌株由深圳清华研究院郑义博士惠赠,pEGFP-N1-PAX4与pEGFP-N1-PDX1质粒为前期本实验室所构建。Anti-PAX4 antibody (ab42450)/IgG、Anti-PDX1 antibody (ab47383)/IgG购自美国Abcam公司,HRP标羊抗鼠IgG、HRP标羊抗兔IgG、FITC标羊抗兔IgG、CY5标羊抗鼠IgG购自eBioscience公司。Lipofectamine 2000购自Invitrogen公司;引物由北京赛百盛基因技术有限公司合成;RT-PCR试剂盒为北京天恩泽公司; DMEM/F12培养基购自武汉博士德生物工程有限公司;胎牛血清(fetal bovine serum,FBS)购自Hyclone公司。原代人脐带间充质干细胞购自北京医科利昊生物科技有限公司。
1.2方法
1) pADxsi-CMV- PDX1/CMV- PAX4腺病毒载体的构建。 首先构建pShuttle-GFP-CMV- PDX1穿梭质粒:已知PDX1序列上游有Nhe I和Bgl II酶切位点,下游有Sal I和EcoR I酶切位点,首先用Sal I酶切pEGFP-N1- PDX1,再用Klenow平端处理,最后用Nhe I酶切,回收0.66 kb片段;其次用Nhe I和Pme I双酶切pShuttle-GFP-CMV载体,替换GFP, CIP去磷酸化处理,回收载体片段5.2 kb;最后,酶连切好的载体片段和插入片段,获得pShuttle-CMV-PDX1。再构建pShuttle-CMV-PDX1/CMV-PAX4穿梭质粒载体:从pEGFP-N1-PAX4上用BamH I和Sal I双酶切切下PAX4(PAX4序列上游存在BamH I和Bgl II酶切位点,下游存在Xba I和Sal I酶切位点);同时对pShuttle-CMV- PDX1载体用BamH I和 Sal I双酶切,CIP去磷酸化处理,胶分别回收与纯化载体与酶切PAX4目的基因片段并用T4 DNA 连接酶酶连得到pADxsi-CMV-PDX1/CMV-PAX4穿梭质粒。最后用T4 DNA 连接酶酶连目的片段pShuttle-CMV-PDX1/CMV-PAX4和pADxsi骨架质粒;转化产物并转染DHSa,扩增pADxsi -CMV-PDX1/CMV- PAX4质粒,对质粒产物进行提取并进行电泳签定,产物由上海丰恒生物科技有限公司进行测序鉴定。
2) 重组腺病毒的包装、扩增和滴度测定。 Pac I酶切线性化重组腺病毒质粒,用Lipofectamine 2000脂质体转染细胞密度约80%的293细胞。3~5 d后,开始出现明显噬斑,待大部分细胞病变(cyto-pathic effect,CPE)时,收集细胞混悬液,于-80℃/25℃反复冻融3次,再离心收集上清,继续感染293细胞扩增病毒以提高腺病毒(ADxsi-CMV-PDX1/CMV-PAX4)滴度,TCID 50法测定病毒滴度。
3) ADxsi-CMV-PDX1/CMV-PAX4感染MSCs与目的基因表达。 于六孔板接种MSCs细胞5.0×105/孔,细胞密度达80%时,按感染指数为10PFU/细胞、50PFU/细胞、100PFU/细胞加入相应量的腺病毒液;同时设ADxsi-CMV-GFP感染的空病毒对照组。当实验进行到相应阶段时,即培养到24h、48h和72h等阶段分别收集细胞行WB、免疫细胞化学与间接荧光检测;同时RT-PCR检测目的基因表达,引物分别如下PDX1 的F:5′-AAGCTAGCCCGCAGCCATGA-3′、R:5′-TCCTCGAGTCATCGTGGTTCCTG-3′,Tm为61.5℃,扩增目的片段为882bp;PAX4:F:5′-TCCCAGTGTCTCCTCCATC-3′、R:5′-ACCTTTCCGGTGCTGTTGC-3′,Tm为60 ℃,扩增目的片段为515 bp;内参照分子GAPDH:F:5′-GTCAGTGGTGGACCTGACCT-3′、R: 5′-TGAGGAGGGGAGATTCAGTG-3′。
(4)Western blotting检测
胰酶消化,收集目的细胞,裂解细胞并离心取上清液,应用15%的SDS-PAGE胶电泳2h后,湿转移法至PVDF膜,脱脂牛奶TBST液封闭1h,再分别加抗人PAX4、PDX1抗体IgG液振荡过夜,HRP标记相应二抗亲合反应后ECL显色。
2结果
2.1鉴定重组腺病毒质粒
Xho I酶切pADxsi-CMV- PDX1/CMV-PAX4病毒质粒,阳性克隆pADxsi-CMV-PDX1/CMV-PAX4由以下6条带组成即:14 kb、11.8 kb、5.9 kb、2.47 kb、1.45 kb、0.6 kb;而阴性克隆(pADxsi骨架质粒)只有以下6条带组成:14 kb、11.8 kb、4.0 kb、2.47 kb、1.45 kb、0.6 kb;酶切结果阳性质粒均与理论预期一致,具体酶切结果(见图1)。
2.2重组腺病毒的包装和滴度测定
Pac I酶切线性化的pADxsi-CMV- PDX1/CMV-PAX4重组腺病毒质粒经脂质体Lipofectamine2000转染293细胞后3 d,开始出现病变效应(见图3)。120 h后,此时约70%细胞悬浮,收集细胞与上清液体,冻融后,再应用293细胞重复扩增、收集病毒液。空斑计数法(PFU)测定病毒滴度为1.0×108 PFU/mL;使用同样方法测定对照组。空载腺病毒ADxsi的滴度为2.0×108 PFU/mL。
2.3 目的基因表达
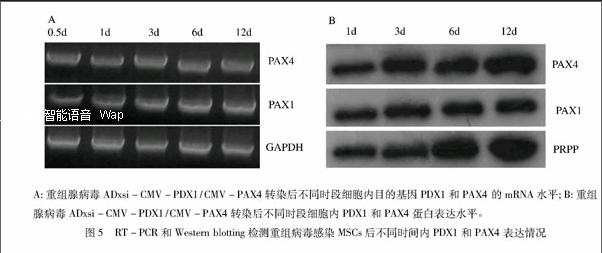
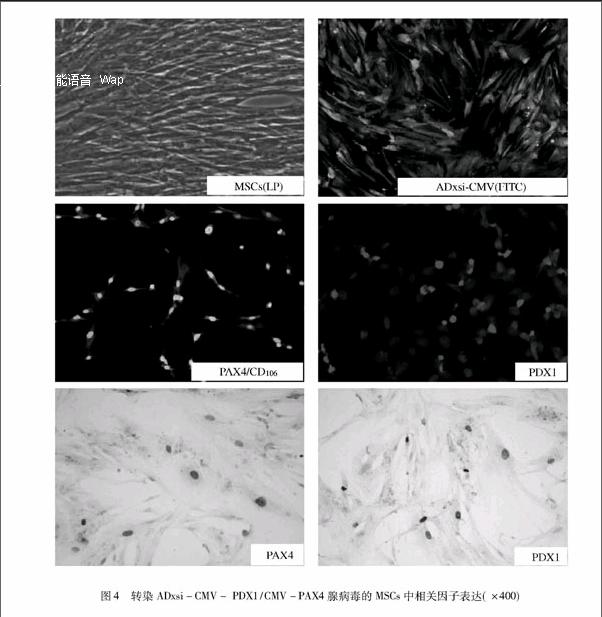
MSCs在光学显微镜下呈典型的长梭形,呈漩涡状或放射状平行排列(见图4); ADxsi-CMV-GFP空病毒感染后24 h即可在荧光镜下观察到GFP表达。免疫细胞化学染色与间接荧光分别证实ADxsi-CMV-PDX1/CMV-PAX4腺病毒以感染复数(MOI)=100感染目的细胞 (MSCs)24 h后,实验组MSCs的细胞核中即可检测到PDX1与PAX4 mRNA;细胞化学检测显示,PDX1与PAX4 主要定位于细胞核内,阳性率高于75%。间接荧光同样证实细胞核中PAX4(FITC)与PDX1 (CY5)均稳定表达(见图4)。
转染后光镜下的细胞形态相同于正常的MSCs细胞形态,呈梭形排列,应用ADxsi-CMV-EGFP空病毒感染细胞后,可见到细胞内绿色荧光表达,分别应用抗体检测PDX1与PAX4表达与定位,荧光结果显示PDX1与PAX4均定位于细胞核内,且免疫细胞化学检测也证实两转录因子定位于细胞核内,且稳定表达。
2.4 目的基因转录与表达
RT-PCR法与WB分别检测重组腺病毒ADxsi-CMV- PDX1/CMV-PAX4转染后不同时段细胞内目的基因mRNA和蛋白表达结果显示:MSCs胞质内PDX1与PAX4 mRNA水平稳定;进一步Western blotting(WB)法检测感染后12 d的细胞核内PDX1与PAX4蛋白一直稳定表达(见图5),提示重组腺病毒ADxsi-CMV-PDX1/CMV-PAX4可以有效感染MSCs,且目的基因均能稳定表达,这为后续研究目的基因在MSCs转分化过程中的功能奠定实验基础。
3讨论
MSCs具有多向分化潜能,为探讨MSCs分化为合成与分泌胰岛素功能的胰腺β细胞,选用了在胰腺前体细胞向β细胞分化过程中起关键作用的两个转录因子PDX1与PAX4[8-11],并构建对MSCs具有稳定转染能力的Ⅴ型腺病毒载体[12-13],用PDX1换去示踪基因EGFP,把PAX4基因插入到多克隆位点,构建并包装成带PDX1与PAX4双目的基因的活性重组腺病毒(ADxsi-CMV-PDX1/CMV-PAX4),酶切结果与测序结果证实重组腺病毒构建成功,目的基因连接正确。
应用ADxsi-CMV-PDX1/CMV-PAX4感染MSCs结果显示,重组病毒可以稳定高效感染MSCs,间接荧光检测结果显示,转染的MSCs在细胞核稳定表达PDX1与PAX4分子,细胞化学染色结果同样证实PDX1与PAX4分子定位于在MSCs细胞核内,这提示带PDX1与PAX4转录因子基因的重组腺病毒不仅可以高效感染目的细胞,并且稳定表达目的转录因子,而且所带的转录因子均具有核定位功能。
另一方面应用RT-PCR技术检测转录重组腺病毒的MSCs细胞内目的基因的转录水平表明目的基因在重组腺病毒感染细胞后仍可以检测到转录目的基因的mRNA水平稳定,提示在腺病毒的CMV启动子的作用下,目的基因在细胞内稳定转录。进一步抽提感染腺病毒的MSCs的细胞核蛋白,并行WB检测PDX1和PAX4转录因子蛋白水平,发现两种转录因子在细胞核水平稳定,这一方面提示腺病毒的CMV启动子具有强的启动目的基因转录功能,另一方面,目的转录因子稳定定位于细胞核,说明表达的转录因子具有核定位能力。的并且能稳定转录与表达PDX1与PAX4分子。
综上检测结果证实, PDX1与PAX4双带目的基因的Ⅴ型重组腺病毒被成功构建,重组腺病毒所带的目的基因均能稳定转录与翻译成功能转录因子PDX1与PAX4,且两功能转录因子均具有核定位功能,这为下一步研究2功能转录因子在诱导MSCs定向向胰腺β细胞分化过程中的功能与分子机制奠定了实验基础。
参考文献:
[1]GAO T, MCKENNA B,Li C, REICHERT M, et al. Pdx1 maintains β cell identity and function by repressing an α cell program [J]. Cell Metab, 2014, 19(2):259-271.
[2]GAGE BK, WEBBER TD, KIEFFER TJ. Initial cell seeding density influences pancreatic endocrine development during in vitro differentiation of human embryonic stem cells[J]. PLoS One , 2013, 8(12):e82076.
[3]RAIKWAR SP, ZAVAZAVA N. PDX1-engineered embryonic stem cell-derived insulin producing cells regulate hyperglycemia in diabetic mice[J]. Transplant Res , 2012, 1(1):19.
[4]LIMA MJ, DOCHERTY HM, CHEN Y, et al. Pancreatic transcription factors containing protein transduction domains drive mouse embryonic stem cells towards endocrine pancreas[J]. PLoS One, 2012, 7(5):36 481.
[5]NAIR GG, VINCENT RK, ODORICO JS. Ectopic Ptf1a expression in murine ESCs potentiates endocrine differentiation and models pancreas development in vitro[J]. Stem Cells , 2014 ,32(5):1 195-1 207.
[6]DHANASEKARAN M, INDUMATHI S, HARIKRISHNAN R, et al. Human omentum fat-derived mesenchymal stem cells transdifferentiates into pancreatic islet-like cluster[J]. Cell Biochem Funct , 2013, 31(7):612-619.
[7]TANG XIAOLONG, WAN ZEMING, GAO YUNLONG, et al. Generation of insulin-producing cells from bone marrow mesenchymal stem cells with modified expression of PDX1 and NKX6.1[J]. Asian Journal of Chemistry, 2014, 26(11):3 417-3 424.
[8]JAFARIAN A, TAGHIKHANI M, ABROUN S, et al.Generation of high-yield insulin producing cells from human bone marrow mesenchymal stem cells[J]. Mol Biol Rep, 2014, 41(7)
:4 783-4 794.
[9]TALAVERA-ADAME D, DAFOE DC. Endothelium-derived essential signals involved in pancreas organogenesis[J]. World J Exp Med , 2015,5(2):40-49.
[10]PEDICA F, BECCARI S, PEDRON S, et al. PDX-1 (pancreatic/duodenal homeobox-1 protein 1) [J]. Pathologica,2014,106(4):315-321.
[11]PONDUGALA PK, SASIKALA M, GUDURU VR, et al. Interferon-γ Decreases Nuclear Localization of Pdx-1 and Triggers β-Cell Dysfunction in Chronic Pancreatitis[J]. J Interferon Cytokine Res,2015, 35(7):523-529.
[12]SEMACHE M, GHISLAIN J, ZARROUKI B, et al. Pancreatic and duodenal homeobox-1 nuclear localization is regulated by glucose in dispersed rat islets but not in insulin-secreting cell lines[J]. Islets, 2014, 6(4):982 376.
[13]JOHNSON JS, KONO T, TONG X, et al. Pancreatic and duodenal homeobox protein 1 (Pdx-1) maintains endoplasmic reticulum calcium levels through transcriptional regulation of sarco-endoplasmic reticulum calcium ATPase 2b (SERCA2b) in the islet β cell[J]. J Biol Chem , 2014, 289(47):32 798-32 810.
(责任编辑:何学华吴晓红)
