柚木无性系光合作用的日季节变化及其主要影响因素
2016-12-21黄桂华梁坤南周再知马华明
黄桂华,梁坤南,周再知,马华明
(中国林业科学研究院热带林业研究所,广东 广州 510520)
柚木无性系光合作用的日季节变化及其主要影响因素
黄桂华,梁坤南,周再知,马华明
(中国林业科学研究院热带林业研究所,广东 广州 510520)
以柚木无性系苗为材料,研究了柚木光合作用及其相关生态因子的日季节变化,分析了柚木光合作用的主要影响因子。结果显示,在广州,柚木6月净光合速率(Pn)的日变化为单峰曲线,高峰值(9.826 μmol·m-2s-1)出现在上午11时;8月和10月净光合速率的日变化则为双峰曲线,出现“午休”现象,第一高峰均出现在上午9时,分别为8.317和11.915 μmol·m-2s-1,8月第二小高峰出现在15:00~16:00之间,10月第二小高峰出现在13:00。不同月份(季节)的柚木净光合速率大小排序为10月>6月>8月。影响柚木光合作用的生态因子主要是光合有效辐射,二者存在极显著正相关;其次是温度,6月和8月中午高温对柚木的光合作用有极显著的抑制作用。
柚木;净光合速率;日变化;季节变化;生态因子
柚木Tectona grandisL. f.天然分布于印度、缅甸、泰国和老挝,为世界名贵的用材树种,素有“万木之王”的美称,在珍贵用材树种中属于生长迅速的类型,由于其材质优良,价格昂贵,柚木在亚洲、非洲、美洲中部和南部国家被广泛引种[1-2]。无论是在天然分布区,还是广泛的引种区,柚木的生长生态环境都有非常复杂和巨大的差异,这使得柚木生长特点以及木材性质都存在明显的差异[3-5]。众所周知,树木生长主要受到光合作用的影响,而光合作用是一个非常复杂的树木生理现象,要受树木生长环境中许多生态因子(光照、温度、CO2浓度和空气湿度等)的影响[6]。目前,黄桂华等(数据未发表)研究了来自不同国家不同种源的柚木无性系苗期生长指标、气体交换和叶绿素荧光参数,分析了柚木不同基因型资源光合生理特征及其与生长的关系,发现净光合速率(Pn)和苗高呈显著正相关。另外,郑飞翔等[7]研究了柚木无性系苗期对模拟酸雨胁迫的光合响应,陈德祥等[8]研究了柚木人工林中不同邻体干扰强度下林木CO2气体交换特征。但这些研究主要集中在柚木基因型或在特定条件下的柚木光合作用的瞬时情况,而对其光合作用季节日变化及各生态因子对其光合作用影响的研究未见报道。研究分析柚木在我国生长环境中光合作用的季节日变化,找出影响柚木生长及光合速率的生态因子,将有利于我国柚木栽培区的划分,剖析柚木在不同生态区的生长特点和木材材性等差异在光合方面的生理机理,从而对我国柚木育种和栽培技术研究具有重要的意义。
1 材料与方法
1.1 材料与试验设计
本研究柚木材料选取了印度3个无性系(7122、7146、7210)、缅甸3个无性系(7559、7509、7531)和1个尼日利亚无性系(3078-5),2011年对各无性系分别进行组培苗轻基质移栽培育,2012年4月挑选各无性系中生长一致的1年生苗30株,盆栽于中国林科院热带林业研究所苗圃(113°18′E,23°06′N),5 株小区,6 次重复,盆间距40 cm。
1.2 指标测定
根据柚木在广州5月初开始萌芽生长,6~10月生长旺盛,11月生长速度减慢,12月叶片变黄,开始脱落的生长规律,2012年分别选择天气晴朗的6月20日、8月20日和10月21日从7:00~19:00,每隔2 h测定1次光合作用指标。每个无性系随机选择3株,且每株从上往下选择第2和第3轮的3片同龄向阳功能叶片,采用美国LI-COR公司Li-6400型便携式光合仪测定包括净光合速率Pn(μmol·m-2s-1)、光合有效辐射PARo(μmol·m-2s-1)、叶室 CO2浓度Ca(μmol·mol-1)、叶室温度 Tair(℃)和叶室相对湿度RH(%)等光合作用指标。测定时,每次按相同的顺序测量每个无性系,每叶片测定3次稳定值。
1.3 数据分析
采用SAS软件程序统计各指标的平均值,采用DPS数据处理软件对光合速率和生态因子进行了逐步多元回归分析、偏相关和通径分析。
2 结果与分析
2.1 柚木净光合速率和生态因子的日变化
对测定的柚木净光合速率及主要生态因子数据进行日变化和季节变化作图分析,从图1可知,在高温高湿的6月,柚木净光合速率的日变化为单峰曲线。7:00~11:00柚木净光合速率在适宜的温度和CO2供应条件下,随光合有效辐射的增强而快速上升,并在11:00左右达到高锋,峰值为 9.826±1.525 μmol·m-2s-1。之后随光合有效辐射的减弱,柚木净光合速率逐渐下降到19:00的-1.564±0.246 μmol·m-2s-1。而在 9:00 ~ 12:00 时间段的净光合速率都维持在较高水平。
在高温的8月,柚木净光合速率的日变化出现双峰曲线。9:00左右出现第一高峰,峰植为8.317±1.138 μmol·m-2s-1,该时段柚木净光合速率主要随光合有效辐射的增强而增强;之后柚木净光合速率快速降低并在12:00左右降至低谷,谷值仅有 2.582±0.736 μmol·m-2s-1,出现大“午休”现象。其原因一方面是该时段温度迅速上升,当叶室温度超过40℃,叶片蒸腾作用强,为防止过量失水,叶片气孔大量关闭,导致CO2供应不足,最终导致了柚木净光合速率迅速下降;另一方面,12:00~13:00光合有效辐射达到顶峰,这时可能出现了光抑制现象。柚木净光合速率在15:00~16:00之间出现当日第二个小高峰,该时段属于温度回落后,气孔逐渐恢复打开状态,CO2供应恢复,净光合速率迅速上升,但此时光合有效辐射下降了,相应净光合速率只小幅上升。最后,柚木净光合速率随光合有效辐射的减弱而快速下降。
温度适中的10月,柚木净光合速率的日变化也为双峰曲线。同样9:00左右出现第一高峰,净光合速率达到 11.915±1.363 μmol·m-2s-1,之后在 11:00左右柚木净光合速率小幅下降,出现小“午休”现象,中午最低值保持有 8.278±1.937 μmol·m-2s-1。数据表明,此时秋季温度不高,CO2供应属合适范围,而净光合速率也出现低谷,说明此时光合有效辐射依然偏高,出现了光抑制现象。随后柚木净光合速率小幅上升并在13:00左右出现第二个小高峰,该时段属于温度和CO2供应属适中,光合有效辐射有所下降,光抑制现象得到解除,因此柚木净光合速率迅速上升,但也因光合有效辐射下降了,净光合速率只能小幅上升。最后,柚木净光合速率随光合有效辐射的减弱而逐渐下降到最低值。测得10月温度相对温和适中(27.93~34.54℃),柚木净光合速率在8:00~16:00都维持在较高水平。

图1 柚木净光合速率及主要生态因子的季节日变化(a-e)Fig. 1 The diurnal and seasonal changes of net photosynthetic and ecological factors on teak
2.2 柚木净光合速率和生态因子的季节变化
植物光合作用是个复杂的生理过程,受多种生态因子的控制。不同月份(季节)的光合强度随着影响因子的变化而不同。从表1可以看出,柚木净光合速率10月>6月>8月,说明柚木在广州的10月叶片光合能力最强,生长最旺盛,其次为6月和8月。而光合有效辐射和叶室温度正好与净光合速率相反,是8月>6月>10月光合有效辐射和叶室温度存在极显著正相关系,说明在特定时空区域,光照越强,叶室温度越高。叶室CO2浓度的变化则与净光合速率一致,是10月>6月>8月,叶室相对湿度是8月>10月>6月。
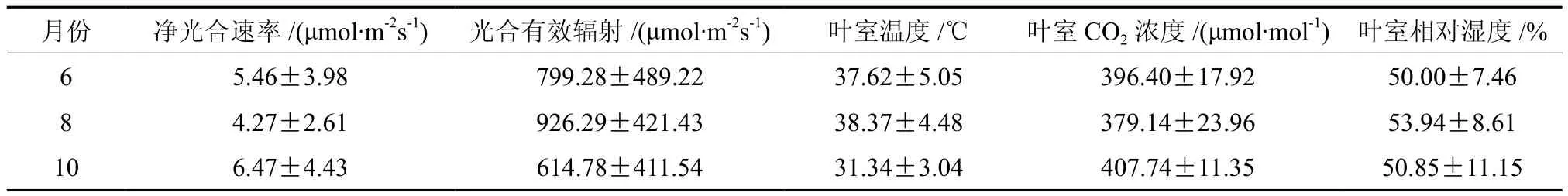
表1 不同月份柚木净光合速率及生态因子的比较†Table 1 The net photosynthetic and ecological factors of teak in different seasons
2.3 柚木净光合速率与主要生态因子间的关系
分别对光合有效辐射PARo(X1)、叶室温度Tair(X2)、叶室CO2浓度Ca(X3)、叶室相对湿度RH(X4)与净光合速率Pn(Y)进行逐步多元回归分析,建立最优回归方程。得到6月回归方程Y=33.488 561 1+0.008 150 162 685X1-0.662 462 854 9X2-0.192 431 764 17X4(复相关系数R=0.822 599,F=31.392 1,P=0.000 0)。回归方程、偏相关分析(表2)和通径分析(表3)的结果一致显示,影响6月柚木净光合速率的主要生态因子是PARo(X1)>Tair(X2)>RH(X4),其中柚木净光合速率与PARo(X1)极显著正相关;与Tair(X2) 极显著负相关,说明6月进入了高温季节,过高的温度对柚木净光合速率有极显著的抑制作用;柚木6月净光合速率与RH(X4)呈负相关,但该影响不显著。

表2 柚木净光合速率与生态因子的偏相关分析Table 2 The partial correlations analysis between net photosynthetic and ecological factors on teak

表3 柚木净光合速率与生态因子的通径分析Table 3 The path analysis between net photosynthetic and ecological factors on teak
8月回归方程Y=87.197 610 8+0.002 650 435 567 7X1-0.915 612 956 2X2-0.132 546 612 86X3(复相关系数R=0.810 029,F=28.623 3,P=0.000 0)。从回归方程可知,对8月柚木净光合速率的影响因素是PARo(X1)、Tair(X2)和Ca(X3)。表2、3显示,各因素与8月柚木净光合速率的相关程度是Tair(X2)>Ca(X3)>PARo(X1),其中与PARo(X1)极显著正相关,与Tair(X2)和Ca(X3)极显著负相关。说明8月柚木净光合速率受到多因子的影响,有的因子是正影响,有的因子是负影响,且这些影响是以交叉互作影响柚木净光合速率的。显然,高温对8月柚木净光合速率有极显著抑制作用。而CO2浓度对柚木净光合速率的极显著负影响,笔者认为是高温下柚木为防止蒸腾旺盛失水,气孔大量关闭,胞间CO2浓度下降,致使柚木净光合速率下降,因此,在高温条件下,即使测得的空气CO2浓度高,柚木净光合速率也可能较低,即呈现出测定的空气CO2浓度高而柚木净光合速率低的负相关的“假象”。
10月回归方程Y=54.117 681 8+0.005 514 817 309X1-0.134 911 368 29X3+0.078 147 963 04X4(复相关系数R=0.801 206,F=26.891 3,P=0.000 0)。从回归方程和表3得出,对10月柚木净光合速率的影响因素是PARo(X1) 、Ca(X3)和RH(X4)。但从表2可知,主要影响柚木10月净光合速率的是PARo(X1),二者存在极显著正相关,与RH(X4)和Ca(X3)相关性不显著。
3 结论与讨论
光合作用是一个受植物自身因子和环境因子综合作用的过程。不同月份(季节)的光照强度、温度等生态因子对净光合速率的影响有很大差别。本研究结果表明,柚木苗期6月净光合速率日变化为单峰曲线,8月和10月净光合速率日变化出现“午休”现象,因而为双峰曲线。不同季节出现第一高峰、“午休”和第二高峰的时间及对应净光合速率值差异较大。柚木净光合速率最大值(11.915±1.363 μmol·m-2s-1)出现在 10 月上午9:00左右,说明广州10月相对适中的气候及其他因子有利于柚木光合作用。另外,关德新等[9]研究认为晴天植物对CO2的吸收最大强度发生在上午9:00时左右,这在本文中得到了印证,8月和10月柚木净光合速率都是在9:00左右出现第一高峰,除了这时温度和光照等生态因子适合外,估计还与此时是吸收CO2的最强时刻有关,而CO2浓度的增加,可以显著促进柚木的光合作用[8]。当然,不同树种日季节光合峰值和“午休”也各有不同,李树日变化最高峰出现在10:00左右,次高峰出现在15:00前后[10]。
“午休”现象是植物在长期的进化和适应过程中形成的抵御和减轻强光和高温胁迫的防御机制[11-14]。植物“午休”的原因可能是多方面的。陈德祥等[8]测得弱邻体干扰强度下的21年生柚木的光饱和点为 822 μmol·m-2s-1,说明在光照强度大于该值时,柚木光合作用不再增强,甚至会出现光抑制现象,而本研究柚木“午休”现象发生时都是处在一天中光照最强的时间段。另本试验苗在夏季蒸发量大和秋季进入干旱季节的条件下,很可能出现了土壤水分不足的情况。因此,8月柚木大“午休”现象的出现,是由强光、高温和土壤干旱引起气孔大量关闭和光合作用光抑制共同导致。10月柚木小“午休”,主要是因为强光和土壤干旱引起气孔少量关闭和光抑制共同导致。这和姜英等[15]研究红椎幼苗的光合作用所获得的结论比较一致,红椎在全光照的30%时,叶绿素含量最高,光合作用最强。
柚木日季节光合和生态因子变化曲线显示,广州6月温度接近8月,而柚木6月没有出现“午休”现象,这点和美洲黑杨[16]净光合速率日进程一样。对比而已,广州6月为雨季水分充足,光合有效辐射也不是特别高,气孔不关闭,柚木光合作用顺利进行。8月温度最高、光合有效辐射最强,蒸腾作用很强,气孔容易关闭,柚木光合作用受阻,严重时可能导致叶片在形态功能上发生改变[17]。10月温度和光合有效辐射适中,但进入干旱季节,土壤水分也可能供应不足,强光下气孔容易关闭。另10月柚木净光合速率白天维持在较高水平,光合作用的产物来不及运走,积累在叶肉细胞的细胞质中,也可能阻碍细胞内二氧化碳的运输,这些原因都将使柚木光合作用受阻。
柚木在广州的生长规律为:5月初开始萌芽生长,6~10月叶片浓绿生长旺盛,11月叶片开始衰老生长速度减慢,12月叶片变黄,开始脱落。根据本研究得出的柚木净光合速率10月>6月>8月。班小泉[18]研究认为,柚木旱季的光合速率高于雨季的光合速率近36%。广州10月开始进入旱季,而4~8月为雨季,因此,二者在不同地点研究的不同树龄的柚木光合速率随旱季和雨季的变化规律是一致的。根据本研究结果,推测柚木在广州光合速率季节变化应为双峰曲线,即柚木5月萌芽嫩叶期光合作用偏慢,6月生长变快,然后8月光合有所降低,10月光合作用又得到加强,最后柚木光合作用减弱直到落叶。姜小文等[19]测得四季柚净光台速率大小依次为10月、6月、11月、7月、8月。可见,柚木光合速率的季节特征和四季柚相近。
本试验结果表明,对柚木净光合速率影响的主要生态因子是光合有效辐射,其次是温度,特别是在6月和8月的中午前后,过高的气温对柚木净光合速率有极显著的抑制作用。由此看来,柚木光合作用的主要影响因子与红椎[15]、樟树[20]基本相同。
在开展本试验过程中,8月同时测了栽种于同苗圃地块的2.5年生柚木小树(高3.5 m,胸径3.3 cm)的净光合速率,最高值为 18.1 μmol·m-2s-1,中午 12:00 光合速率也达到了 8.12 μmol·m-2s-1。据此推测,柚木大树的光合作用要强于苗期。另一方面,班小泉[18]研究海南省尖峰岭21年生柚木人工林不同林体干扰程度下的光合速率,发现旱季日平均光合速率为 6.36 μmol·m-2s-1和雨季为 3.88 μmol·m-2s-1,分别对应接近本研究结果的10月(旱季)6.47±4.43 μmol·m-2s-1和 8 月(雨季)4.27±2.61 μmol·m-2s-1、6 月(雨季)5.46±3.98 μmol·m-2s-11。本试验柚木苗期盆栽在一定程度上限制了柚木的生长,且8月高温和10月旱季有一定程度的水分胁迫发生。因此,本试验测得的光合速率整体都低于自然生长条件下柚木的光合速率,但本试验条件下,也真实可靠地反映了柚木光合速率的日变化和季节变化规律。植物光合作用的测定受天气条件影响很大,本研究每个季节只测定一天,今后这方面的研究需要连续重复测定3天 更具有科学性。本研究结果还表明,6月上午8:30~11:30柚木光合速率相对8月和10月比较平稳,适合测定比较不同无性系的瞬时光合速率。
[1]Kadambi K. Silviculture and management of teak [M]. Bulletin 24 School of Forestry, Stephen F. Austin State University,Nacogdoches, Texas, 1972: 137.
[2]White K J. Teak: some aspects of research and development [M].Yangon: San Thain Kha Press, 1991: 70.
[3]梁坤南, 赖 猛, 黄桂华, 等. 10个柚木种源27年生长与适应性[J]. 中南林业科技大学学报, 2011, 31(4): 8-12.
[4]赖 猛, 梁坤南, 黄桂华, 等. 柚木种源生长和与材质有关特性的遗传变异及综合评价[J]. 林业科学研究, 2011, 24(2):234-238.
[5]Bendale V W, Naik R Y, Mehta J L,et al. Variability studies in teak [J]. Journal of Ecobiology, 2005, (17): 29-34.
[6]Turnbull M H, Murthy R, Grif fi n K L. The relative impacts of day-time and night-time warming on photosynthetic capacity in Populus deltoids [J]. Plant, Cell and Environment, 2002, 25:1729-1737.
[7]郑飞翔, 温达志, 旷远文. 模拟酸雨对柚木幼苗生长、光合与水分利用的影响[J]. 热带亚热带植物学报, 2006, 14(2): 93-99.
[8]陈德祥, 班小泉, 李意德, 等. 热带柚木(Tectona grandisL.f.)人工林中不同邻体干扰强度下林木CO2气体交换特征[J].生态学报, 2008, 28(9): 4059-4069.
[9]关德新, 吴家兵, 金昌杰, 等. 长白山红松针阔混交林CO2通量的日变化与季节变化[J]. 林业科学, 2006, 42(10): 123-128..
[10]于泽源, 许姣卉, 霍俊伟. 李光合特性的研究[J]. 东北农业大学学报, 2004, 35(3): 315-317.
[11]张有福, 陈春艳, 陈 拓, 等. 2种圆柏属植物叶绿素荧光对季节变化的响应特征[J]. 植物研究, 2010, 30(3): 289-293.
[12]许大全. 植物光胁迫研究中的几个问题[J]. 植物生理学通讯,2003, 39(5): 493-495.
[13]王 强, 温晓刚, 张其德. 光合作用光抑制的研究进展[J]. 植物学通报, 2003, 20(5): 539-548.
[14]Demmig-Adams B, Adams W W I, Logan B A,et al.Xanthophyll cycle-dependent energy dissipation and flexible photosystemⅡef fi ciency in plants acclimated to light stress [J]. Functional Plant Biology, 1995, 22(2): 249-260.
[15]姜 英, 黄志玲, 郝海坤. 不同光照强度下红锥幼苗叶绿素荧光参数的季节变化[J]. 西南林业大学学报, 2014, 34(2): 8-12..
[16]李文文, 黄秦军, 丁昌俊, 等. 南方型和北方型美洲黑杨幼苗光合作用的日季节变化[J].林业科学研究,2010,23(2):227-233.
[17]Maseyk K S, Lin T B, Rotenberg E,et al. PhysiologyI-phenology interaction in a productive semi-arid pine forest [J]. New Phytologist, 2008, 178: 603-616.
[18]班小泉. 海南岛尖峰岭柚木光合生理生态和树干液流特征研究[D]. 北京:中国林业科学研究院,2005.
[19]姜小文, 张秋踞, 易干军, 等. 四季柚的光合特性研究[J]. 湖南农业大学学报:自然科学版, 2003, 29(5): 397-401.
[20]黄志宏, 田大伦, 闫文德, 等. 城市樟树人工林冠层光合作用的时空特征[J]. 中南林业科技大学学报, 2011, 31(1): 39-46.
Diurnal and seasonal photosynthetic characteristics and in fluencing factors in teak clones
HUANG Gui-hua, LIANG Kun-nan, ZHOU Zai-zhi, MA Hua-ming
(Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou 510520, Guangdong, China)
The photosynthetic characteristics and in fluencing factors have been studied on clones younglings ofTectona grandisby Li-6400 photosynthetic system, the results showed that the curves of diurnal variations of net photosynthetic rate had one single peak curve in June at Guangzhou, the peak value 9.826 μmol·m-2s-1was at 11:00, while it showed two-peak curves with “photosynthetic midday depression” in August and October, the highest peak values 8.317 and 11.915 μmol·m-2s-1respectively were at 9:00, the second peak value was around 15:00 to 16:00 in August and at 13:00 in October. The average net photosynthetic rate of teak was October>June>August. Photosynthetic available radiation (PARo) is the most important ecological factor in fluencing the net photosynthetic rate of teak, and then is air temperature. PAR has great signi fi cant positive correlation with net photosynthetic rate, high temperature in June and August has great signi fi cant depressing effect to net photosynthetic rate.
teak; net photosynthesis rate; diurnal changes; seasonal changes; ecological factors
S796
A
1673-923X(2016)04-0011-06
10.14067/j.cnki.1673-923x.2016.04.003
http: //qks.csuft.edu.cn
2015-03-11
“十二五”国家科技支撑计划“柚木和西南桦珍贵用材林定向培育技术研究与示范”课题(2012BAD21B01)
黄桂华,博士,副研究员;E-mail:hghcom@sina.com
黄桂华,梁坤南,周再知,等. 柚木无性系光合作用的日季节变化及其主要影响因素[J].中南林业科技大学学报,2016,36(4): 11-16.
[本文编校:吴 彬]
