不同年龄橡胶-催吐萝芙木-降香黄檀复合生态系统中植物的生长动态及其生物量
2016-12-20王飞军陈蕾西刘成刚刘长安唐建维
王飞军,陈蕾西,刘成刚,刘长安,唐建维
(1.中国科学院大学,北京 100039;2.中国科学院 西双版纳热带植物园 热带植物资源可持续利用重点实验室,云南 勐腊 666303;3.中国药科大学 生命科学与技术学院,江苏 南京 211198)
不同年龄橡胶-催吐萝芙木-降香黄檀复合生态系统中植物的生长动态及其生物量
王飞军1,2,陈蕾西3,刘成刚2,刘长安2,唐建维2
(1.中国科学院大学,北京 100039;2.中国科学院 西双版纳热带植物园 热带植物资源可持续利用重点实验室,云南 勐腊 666303;3.中国药科大学 生命科学与技术学院,江苏 南京 211198)
根据2个不同年龄橡胶—催吐萝芙木—降香黄檀复合生态系统样地的调查数据,采用所建立的生物量回归方程,分析了2个不同年龄复合生态系统中植物的生长动态、生物量及其分配特征。结果表明:4年生橡胶林下催吐萝芙木和降香黄檀的年均胸径生长量均显著大于7年生橡胶林下催吐萝芙木和降香黄檀的生长量,但年均高度生长量则无显著差异;在种植4.5 a后,前者的生物量分别是后者的2.86倍和2.90倍;4年生橡胶林复合生态系统的总生物量(67.49 t·hm-2)显著大于其单一橡胶林的生物量(47.06 t·hm-2),并超过7年生橡胶林复合生态系统(63.86 t·hm-2)及单一橡胶林的总生物量(60.38 t·hm-2);生物量的器官分配比例除4年生复合系统中的催吐萝芙木为茎>枝>根>叶外,均呈现出茎>根>枝>叶的趋势;随着橡胶年龄的增长,催吐萝芙木和降香黄檀的生长量和生物量虽呈下降的趋势,但橡胶的生物量及胶林复合生态系统的总生物量呈增长的趋势。橡胶林下催吐萝芙木和降香黄檀的种植具有提高生物量积累和碳固存的潜力和优势。
复合生态系统;橡胶;催吐萝芙木;降香黄檀;生长动态;生物量
橡胶Hevea brasiliensis原产于亚马逊热带雨林,其生长需要充足的热量和分配均匀的降水[1],大致分布于10°N~10°S,最高海拔600 m[2]。近半个世纪以来,随着对天然橡胶需求量的增加,橡胶已被引种到了东南亚、南美、非洲的许多高纬、高海拔地区[3]。橡胶现已经是热带地区尤其是东南亚广泛种植的一种经济树种,其种植面积的不断扩大导致了东南亚许多地区的天然林和次生林被取代[4-5]。对西双版纳地区而言,近30年来最显著的土地利用变化就是大面积的热带雨林被结构单一的纯橡胶林所替代,橡胶面积达40万hm2[3],橡胶树也“爬升”到了海拔1 100 m高度,与季节雨林、次生林共同组成西双版纳地区主要的森林类型[1,6-8]。发展橡胶产业不仅能提供橡胶战略资源,促进地方经济社会的发展,还能够适当缓解天然林的保护压力,但单一连片大规模地种植橡胶也带来了一些不利影响,如物种多样性的丧失[6,9],生态环境平衡受到威胁,局部地区水文、气候格局的改变[10-12],雾凉季雾气减少等[13-14]。
农林复合生态系统作为一种可持续的土地利用方式,通过利用不同物种的生态互补功能,能够提高养分的吸收利用效率、减缓水土流失、提高生物多样性和生态系统生产力等[15-20]。在热带地区,作为缓解森林砍伐的最有效手段,能够改善当地农村人口的生活水平[21],在环境脆弱和资源短缺的地区发挥着巨大的作用[20]。基于大面积单一的橡胶种植所出现的问题以及为了提高胶园的经济、生态效益和胶园稳定性的需要,单一橡胶林下的复合种植受到了广泛关注,组建了以橡胶为主的多种多样的农林复合生态系统模式,如橡胶—茶叶、橡胶—咖啡、橡胶—香蕉、橡胶—甘蔗、橡胶—菠萝、橡胶—大叶千斤拔、橡胶—多种植物等。而橡胶—萝芙木—降香黄檀组合模式就是其中之一。
催吐萝芙木Rauvol fi a vomitoria是夹竹桃科Apocynaceae萝芙木属常绿灌木植物,原产于热带非洲,20 世纪60年代被引种到我国。催吐萝芙木具有繁衍方法简单、生长迅速、根中利血平含量较高等优点[22]。目前在我国云南、海南、广东、广西等地均已有人工栽培[23]。从萝芙木属植物根中分离得到了萝芙木碱、利血平等15种生物碱[24],利血平(Reserpine)是其中起降压作用的主要药效成分[25]。目前催吐萝芙木的根在国内主要作为提取利血平的原料,并将其总碱制成的降压灵(Verticil)和降压平(Resemine)应用于临床[26]。因国内对降压药的需求不断增大,野生萝芙木资源出现枯竭,因此应加强萝芙木属植物的人工种植,以满足国内对萝芙木资源的需求。
降香黄檀Dalbergia odorifera是蝶形花科黄檀属植物,为常绿半落叶乔木树种。原产于我国海南省,主要分布于海南西部、西南部和南部,其木材质地坚硬沉重,纹理细密,是制作高档家具的上等用材。降香黄檀的木材含有芳香油,以心材蒸馏得到的降香油是香料的定香剂,也是高级镇痛药材,有抗血凝、氧化、扩冠脉等作用[27-28]。降香黄檀因其具有较高的经济价值而导致大量砍伐,其天然林现已残存不多,濒临灭绝,亟需进行拯救、保存这一濒危植物[29]。
基于目前西双版纳地区橡胶大面积单一种植、土地利用效率低及其对生态环境产生了严重影响的问题[30],根据橡胶、催吐萝芙木和降香黄檀的生态学特性,于2010年在西双版纳2个不同林龄的橡胶林下种植了催吐萝芙木和降香黄檀,进行了橡胶—催吐萝芙木—降香黄檀的组合模式试验,即在橡胶的行距间种植药用植物——催吐萝芙木及珍贵用材树种——降香黄檀,并以橡胶纯林作对照。其试验目的就是改变目前橡胶单一种植的土地利用格局,提高单一种植橡胶林的土地利用效率及经济与生态效益,以改善橡胶种植区域的生态环境。本研究仅就新组建的橡胶—催吐萝芙木—降香黄檀的组合模式中的植物生长动态及其生物量进行初步分析,以期为探索环境友好型橡胶园的新模式建设、热带山地的退化恢复及可持续利用、开发和种植药用及珍贵树种新途径提供科学依据。
1 研究地概况
研究地点位于西双版纳傣族自治州勐腊县境内,该地区属于西南热带季风气候,干、湿季变化明显,一年可分为干热、湿热和雾凉3季,3~5月为干热季,气温较高,雨量较少;6~10 月为雨季(湿热季),气候湿热,此时期集中了全年降水的85%;11月至次年2月为雾凉季,降雨较少,但早晚有浓雾,空气湿度较大,可以弥补此时期降雨量的不足。年平均气温21.5 ℃,≥10 ℃积温7 860 ℃,平均最低气温7.5 ℃,年日照时数1 828 h,年均降水量达1 539 mm。土壤为由白垩纪砂岩发育而成的砖红壤,pH值在5.0左右[30],各样地基本特征见表1。

表1 不同年龄橡胶—催吐萝芙木—降香黄檀复合生态系统样地的基本特征Table 1 Characteristics of plots in mixed rubber-Rauvolfia vomitoria-Dalbergia odorifera systems with two different stand ages
2 材料与方法
2.1 组合模式的试验设计
样地主要分布在海拔885 m左右的山地丘陵。橡胶的种植规格为8 m×2.5 m,分别为2003、2006年6月种植。2010年6月在2个不同年龄的一部分橡胶林的行距间种植2行催吐萝芙木和1行降香黄檀,另一部分作为对照;催吐萝芙木的种植规格为2 m×3 m,离橡胶2.0 m,每公顷为1 200株;在2行催吐萝芙木的中间种植1行降香黄檀,株距为3 m,每公顷为600株。在上述2个不同年龄的组合模式及橡胶纯林(对照)中分别建立3块25 m × 20 m的样地。
2.2 植物生长调查及生物量的测定
自2010年至2014年,每年12月对样地里所有橡胶个体的胸径、高度分别进行一次测量和估测;催吐萝芙木和降香黄檀则在每块样地的上、中、下分别选取1行依次对其中的10 株个体进行基径、高度的调查。所有个体均进行了标号并在胸径或基径处用红油漆作标记。橡胶生物量的测定采用已经建立的生物量回归模型[31]进行计算(见表2)。

表2 西双版纳橡胶的生物量回归模型†Table 2 Regression models for biomass of rubber in Xishuangbanna area
为建立催吐萝芙木和降香黄檀的生物量模型,在样地外分别选取了29株不同大小的催吐萝芙木和20株降香黄檀个体,采用收获法对其生物量进行测定。利用样本各器官(茎、枝、叶、根)的干质量及总干质量建立各器官生物量及总生物量与样木的基径和高度的生物量回归方程。根据样地中催吐萝芙木和降香黄檀的每木调查结果,采用所建立的生物量回归方程,分别计算样地催吐萝芙木和降香黄檀各器官的生物量,其总生物量则是各器官的生物量之和。在本实验中,因对林下进行了抚育管理,故未对草本层生物量及凋落物量等进行估算。
2.3 数据处理及统计分析方法
将数据录入Excel 2007表进行基本统计,然后采用SPSS17.0进行统计分析,采用单因素方差分析和t检验进行显著性检验,显著水平设定为α=0.05。
3 结果与分析
3.1 不同年龄组合模式中的植物生长动态
对2个年龄不同种植模式中植物生长动态的调查结果表明,种植在4年生橡胶林下的催吐萝芙木和降香黄檀不论是年均基径生长还是高度生长都要大于7年生橡胶林下的催吐萝芙木和降香黄檀(见表3),尤其是2个树种的年均基径生长,4年生橡胶林下的催吐萝芙木(2.06 cm)和降香黄檀(0.91 cm)分别是7年生的1.67倍和1.54倍(p<0.01);而2个树种的年均高度生长则稍大之,并无显著差异(p>0.05)。就2个年龄不同种植模式中的橡胶生长而言,不论是4年生组合模式还是7年生组合模式,橡胶的年均胸径生长和高度生长都要稍大于单一种植的橡胶(对照)(p>0.05)。同样,4年生组合模式中的橡胶及单一种植的橡胶(对照)的年均胸径生长和高度生长也稍大于7年生组合模式的橡胶及单一种植的橡胶(对照)(p>0.05)。这表明在2个年龄不同的种植模式中,虽然3种植物的年均基径生长和高度生长量各不相同,但4年生的橡胶及组合模式中植物的生长量均要大于7年生的橡胶及组合模式中的植物生长量。同时也表明随着橡胶年龄的增长,各种植物的生长呈逐渐减慢的趋势。
3.2 催吐萝芙木和降香黄檀生物量回归模型的建立
根据野外对催吐萝芙木和降香黄檀不同大小个体各器官生物量的调查数据,分别以催吐萝芙木和降香黄檀基径的平方(D2)和高度(H)的乘积(DB2H)为自变量,采用多种线性和非线性的回归方程对两个树种各个体器官的生物量(W)逐一加以拟合,从中选取相关系数最高者作为回归模型。结果显示,以幂函数拟合的生物量模型的相关系数最高,且均达到了极显著水平(见表4)。表明拟合的生物量模型较好地反映了催吐萝芙木降香黄檀个体各器官的生物量。
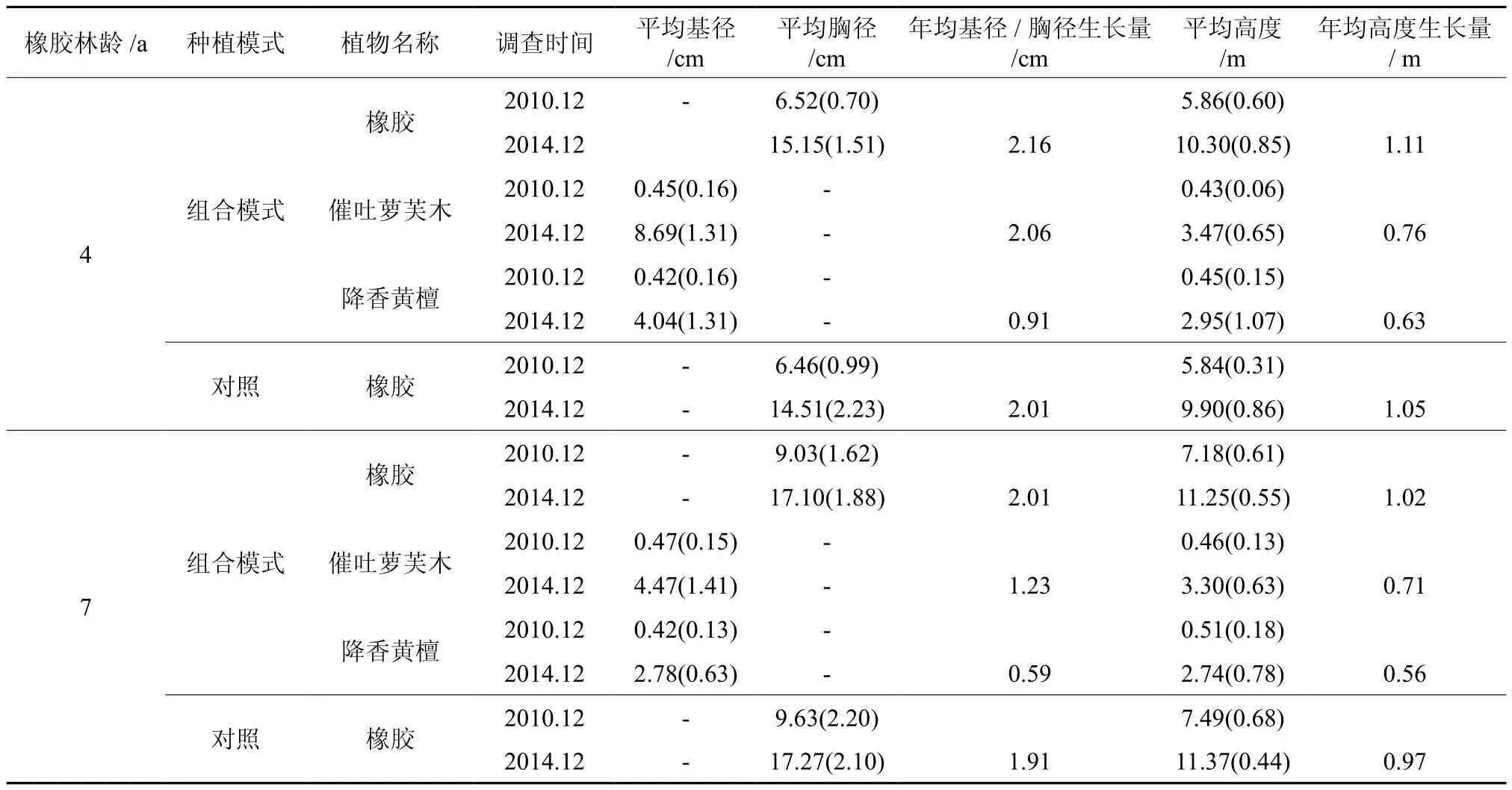
表3 不同年龄橡胶—催吐萝芙木—降香黄檀复合生态系统及橡胶纯林中植物的生长动态†Table 3 Dynamics of plant growth in mixed rubber-R.vomitoria-D.odoriferasystems and rubber pute plantations with two different stand ages
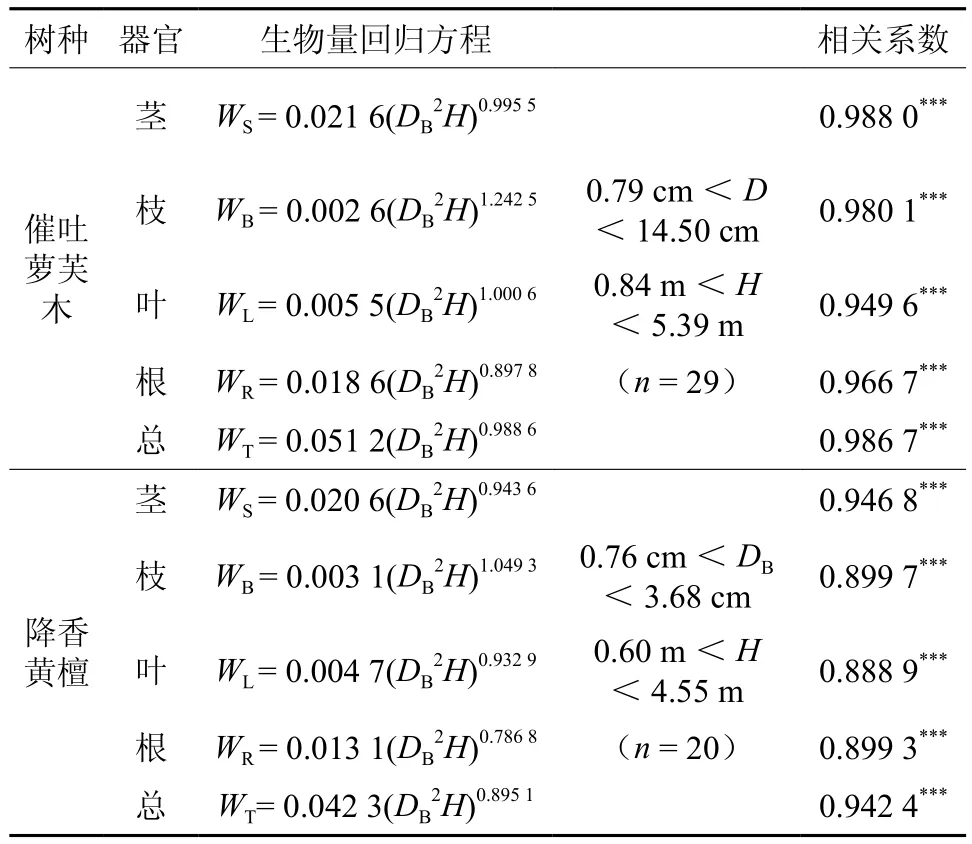
表4 萝芙木和降香黄檀各器官的生物量回归方程†Table 4 Biomass regression equations fororgans of R. vomitoria and D. odorifera
3.3 不同种植模式的生物量及其器官分配
根据2个年龄不同种植模式各样地的植物生长调查数据,采用已建立的生物量回归模型,对其植物个体的生物量进行估算,得到2个年龄不同种植模式中各样地的生物量(见表5)。在种植催吐萝芙木和降香黄檀4.5 a后, 4年生的组合模式的总生物量主要以橡胶的生物量最大,占总生物量(67.49 t·hm2)的73.4%;催吐萝芙木占24.3%,而降香黄檀的生物量仅占总量的2.2%。与单一橡胶林(对照)相比,4年生组合模式中的橡胶生物量(49.63 t·hm2)稍大于单一的橡胶林(对照)(47.06 t·hm2)(p>0.05)。但4年生组合模式的总生物量则显著大于单一的橡胶林(对照)的生物量(p<0.05),约是单一橡胶林的1.43倍。在7年生的组合模式中,橡胶的生物量是其最主要的组成部分,占其总生物量(63.86 t·hm2)的90.24%;催吐萝芙木占8.97%,降香黄檀的生物量仅占总量的0.8%。与4年生组合模式及单一橡胶林生物量不同的是:7年生组合模式中的橡胶生物量(57.63 t·hm2)略小于单一的橡胶林(对照)(60.38 t·hm2)(p>0.05)。但7年生组合模式的总生物量则稍大于单一的橡胶林(对照)的生物量(p>0.05),说明单一橡胶林中植物种类的增加可以提高其生态系统的生物量。就2个年龄不同种植模式中橡胶的生物量及总生物量来看,不论是4年生组合模式中的橡胶生物量还是单一橡胶林(对照)的生物量均显著小于7年生组合模式中的橡胶生物量和单一橡胶林的生物量(p<0.05),但4年生组合模式的总生物量则大于7年生组合模式的总生物量(p>0.05)和单一橡胶林的生物量(p<0.05)。这主要是4年生组合模式中的催吐萝芙木和降香黄檀的生物量显著大于7年生组合模式中的催吐萝芙木和降香黄檀的生物量所致(p<0.05)。前者2个树种的生物量分别是后者2个树种生物量的2.86倍和2.90倍。
在2个年龄不同种植模式生物量的器官分配中,4年生组合模式的生物量主要以茎所占比例最大,约占总生物量的3/5;根次之,占1/5以上;枝约占1/6;叶的生物量均最少,呈现出茎>根>枝>叶的分配特点。而4年生单一橡胶林(对照)生物量的器官分配与4年生组合模式的生物量的器官分配特点基本一致,即茎约占总生物量的3/5;根约占1/5;枝约占1/6;叶的生物量均最少。就4年生组合模式中3个树种生物量各器官的分配比例而言,橡胶生物量各器官的分配比例基本与4年生单一橡胶林(对照)生物量的器官分配比例相同。降香黄檀生物量的器官分配特点虽与橡胶的器官分配一样,即茎>根>枝>叶,但茎所占比例不足总生物量的3/5;根占1/6以上;枝约占1/7;叶所占的比例则超过1/8。而催吐萝芙木生物量的器官分配则体现出与橡胶和降香黄檀不一样的特点,虽然仍以茎所占的比例最高,但约占总量的2/5;而枝所占的比例(22.87%)则稍大于根(21.84%),叶的生物量约占1/9,其器官的生物量分配比例呈现出茎>枝>根>叶的趋势。
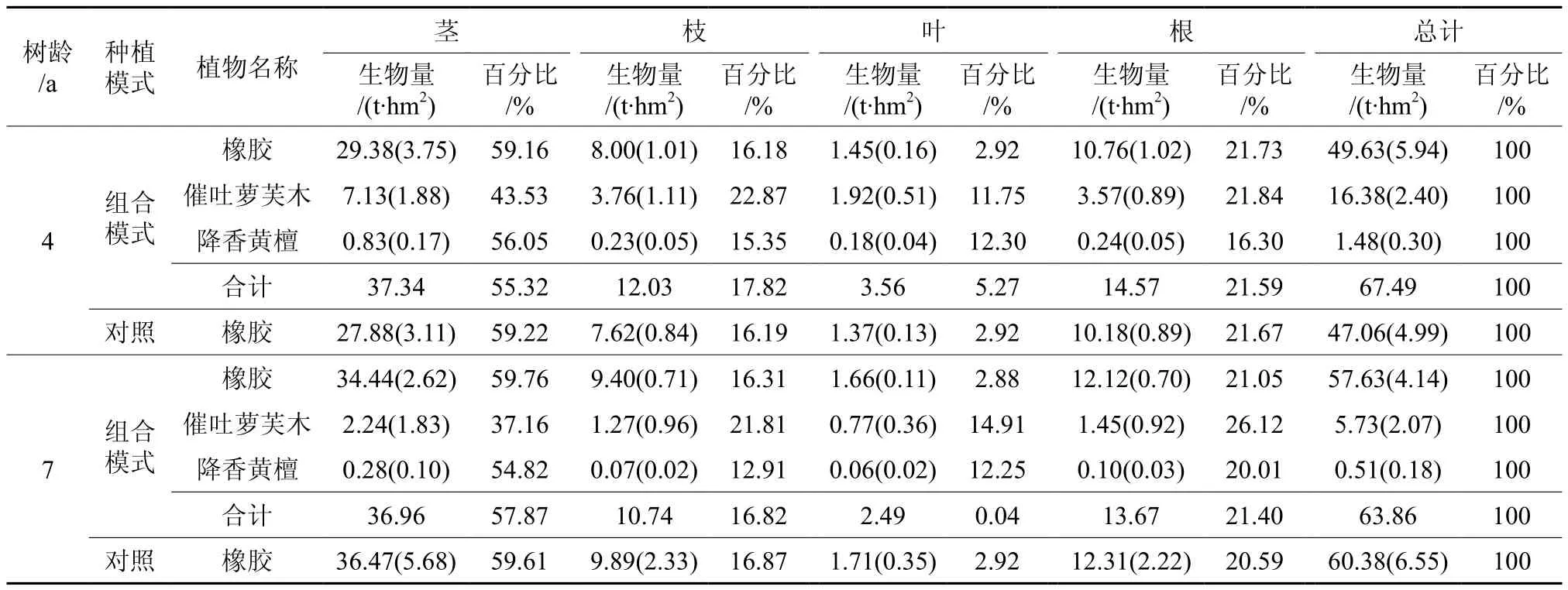
表5 不同年龄橡胶—催吐萝芙木—降香黄檀复合生态系统及橡胶纯林生物量和器官分配†Table 5 Biomass (t·hm2) and biomass allocation among various plant organs in mixed rubber-R.vomitoria-D.odoriferasystems and rubber pure plantationswithtwo different stand ages
在7年生组合模式生物量的器官分配中,虽与4年生组合模式生物量的器官分配特点一致,但其分配比例则有所不同,茎所占比例低于3/5;根次之,占1/5以上;枝占1/6;叶所占的比例最少。而7年生单一橡胶林(对照)生物量的器官分配特点与7年生组合模式一致,仅茎和叶所占的比例分别提高了1.74% 和2.88%,根下降了0.81%,枝所占比例则无变化。在7年生组合模式3个树种生物量的器官分配中,橡胶生物量的器官分配比例同样与其对照的生物量分配比例基本相同。催吐萝芙木生物量的器官分配以茎所占比例最高,占1/3以上;根次之,超过1/4;枝约占1/5 ;叶的生物量约占1/7。降香黄檀生物量的器官分配以茎所占比例最高,占1/2以上;根为1/5,枝所占的比例则稍大于叶,约占1/8。这表明7年生组合模式和单一橡胶林(对照)的总生物量及每种植物生物量的器官分配均呈现出如下趋势:茎>根>枝>叶。
4 讨 论
4.1 不同年龄的橡胶对林下植物生长的影响
在农林复合生态系统中,植物之间的互作存在着光照、热量、水分和养分等空间资源的竞争。上层树种不同的遮荫度所造成的光照强弱直接影响着植物的光合作用、生长速率及其产量[32]。在本试验中,虽然没有测定 2 个不同年龄橡胶林的光照强度,但在相同的种植规格下7 年生橡胶林的遮荫度大于 4 年生的橡胶林,以致种植在 4 年生橡胶林中的催吐萝芙木和降香黄檀的年均基径生长及高度生长均大于 7 年生橡胶林中的 2 个树种的生长。王兴祥等[33]对 5 年生和 9 年生南酸枣—花生复合系统的研究表明,随着南酸枣年龄的增长,上层遮荫度增大,导致了林下花生种植区域光合有效辐射的降低,致使花生产量下降;而且复合系统中的花生产量、生物量与相对光合有效辐射值呈显著正相关。裴保华等[34]对杨农间作的研究也证实光照不足是抑制花生生长、产量降低的主要原因之一。这也说明了复合生态系统中光照对林下植物生长的重要性。这主要是对喜光的植物(如花生等)所产生的影响,但对某些耐荫植物或作物而言则具有促进作用,如茶园中上层遮荫树种的种植则有利于提高茶叶产量和品质[35-36]。这也说明在组建上层以乔木树种为主的复合生态系统时,林下种植的植物应优先选择较耐荫的植物或作物为佳。本研究中 2 个不同年龄橡胶林下种植的 2 种植物目前在生长方面虽有所差异,但均能正常健康地生长。随着橡胶年龄的增长,上层遮荫度的增加,其生长动态如何?对群落结构、功能等方面又会产生怎样的影响?仍有待于进一步长期的观测和研究。
4.2 林下植物种类的增加对群落生物量及生产力的影响
植物种类、层次结构单一的人工橡胶林,不仅造成土壤的有机质和氮含量下降,使土壤贫瘠、退化,且易导致水土流失严重及土壤中的营养元素流失,同时系统的稳定性较差[30,37-39]。而农林复合生态系统通过植物对地上部分光、热资源和对地下水分和养分资源的充分有效的利用及其互补效应,达到充分利用资源及可持续发展的目的[40]。已有的研究表明,不同植物的间作不仅可改善系统中的小气候环境,调节土层温度,减少土壤水分蒸发,改善土壤养分循环,促进植物的生长[40]。目前对橡胶与其他植物间作的复合生态系统模式已进行了大量的研究。曹建华等[41]对橡胶和唐鬼桑、黑籽雀稗和柱花草间作的复合生态系统研究表明,适宜的间作能够改善胶园的环境小气候,在夏秋高温季节能明显地降低地表空气和土壤的温度,减少土壤水分的蒸发等作用,以达到减少高温和干旱对橡胶树的伤害,从而提高了胶树的光合效率和光能利用率,促进橡胶树特别是幼龄树的胸径生长。梁艳丽等[42]对橡胶与魔芋进行间作的研究发现,魔芋对橡胶的生长有极显著地促进作用。橡胶与茶叶的间作,不仅减少了水土流失,改善了土壤结构及土壤水分和养分,而且提高了橡胶的产量[37]。橡胶林下大叶千斤拔的种植,使土壤含水量的季节性变化较为缓慢且旱季含水量较高,并有效地提高了土壤肥力,促进了橡胶生长[30]。而橡胶林下种植茶叶、咖啡及多种植物的复合生态系统的生物量是橡胶纯林的1.15~1.25 倍[43]。杨桂明[44]对橡胶与玉桂的间作试验表明,胶桂间作不仅增强了橡胶的抗风、抗寒能力,减少了胶园杂草丛生和水土流失,且促进了胶树生长和干胶产量的增加。范鸿雁等[45]对幼龄橡胶园间作香蕉高效栽培模式进行了研究,发现间种香蕉三茬后,间作双行橡胶为最佳种植模式,不仅香蕉生长和果品质量未受到影响,而且还促进了橡胶的胸径和高度的生长。本试验4年生组合模式中催吐萝芙木和降香黄檀的基径和高度生长量以及生物量均大于7年生组合模式,而且4年生和7年生组合模式的生物量均高于单一的橡胶林,这也说明了橡胶林下其他植物种类的种植,使复合生态系统具有更大的生物量积累优势和碳储存潜力。此外,有关其他树种的混交试验表明,杉木林下砂仁的种植不仅能提高土壤肥力,而且能促进杉木的生长[46]。沙棘与杨树混交后均能显著地增加杨树生产量,林分平均胸径增加6% ~ 38%,林分平均高度增加8% ~ 23%,地上部分净生产力混交林比纯林高27% ~ 113%[47];华南马尾松纯林的生产力要明显低于由马尾松、藜蒴栲组成的混交林[48];而福建的杉木、马尾松、木荷混交林的生产力明显高于单一的纯林[49]。在澳大利亚,桉树和固氮树种的混交林不仅可维持土壤肥力,同时也能增加林分的生产力[50]。这表明单一人工林中植物种类的增加可提高林分的生物量和生产力。
4.3 不同种植模式的生物量与其他人工林的比较
森林群落的生物量和生产力是森林生态系统重要的功能特征之一。不同森林群落生物量的比较,将有助于了解各种森林群落类型在结构和功能上的变化[51]。本研究中,4年生单一橡胶林及组合模式的生物量分别为 47.06 t·hm-2和 67.49 t·hm-2,而7年生单一橡胶林及组合模式的生物量分别为60.48 t·hm-2和 63.86 t·hm-2。说明随着橡胶年龄的增大以及林下植物种类的增加,系统生物量的积累迅速增加。与我国热带及亚热带地区的其他人工林相比,4年生单一橡胶林及组合模式的生物量分别是云南6~10年生思茅松人工林(27.24 t·hm-2)[52]的 1.73倍和 2.48倍,是广西 6年生马尾松人工林生物量(15.03 t·hm-2)和桉树人工林生物量(12.49 t·hm-2)[53]的 3.13倍、4.49倍和 3.77倍、5.40倍。而7年生单一橡胶林及组合模式的生物量是福建7年生千年桐生物量(14.01 t·hm-2)[54]的4.32倍和4.56倍以及9年千年桐生物量(14.89 t·hm-2)[54]的 4.06倍和 4.29倍,是 7年生不同坡位乐昌含笑人工林生物量(23.04 t·hm-2)[55]的2.63倍和2.77倍,是7年生秋茄人工林生物量(25.36 t·hm-2)[56]的 2.38倍和 2.52倍,是湖南 12年生闽楠人工林生物量(52.25 t·hm-2)[57]的 1.16倍和1.22倍。这说明西双版纳的橡胶林具有较大的生物量积累速率,而其橡胶林复合生态系统则具有更大的生物量积累优势和碳储存潜力[43]。
[1]Vogel A W,Wang M Z,Huang X Q. People’s Republic of China: reference soil (lotosal) of tropical southern Yunnan province[C]. Institute of soil science-Academica Sinica,Nanjing,and International soil reference and information center,Wageningen.1995.
[2]Guardiola-Claramonte M,Troch P A,Ziegler A D,et al.Hydrologic effects of the expansion of rubber (Hevea brasIliensis) in a tropical catchment[J]. Ecohydrolgy,2010,3(3):306-314.
[3]Liu W J,Li J T, Lu H J,et al. Vertical patterns of soil water acquisition by non-native rubber trees (Hevea brasiliensis) in Xishuangbanna,southwest China[J]. Ecohydrology,2013,6(4):1234-1244.
[4]Zhang H,Zhang G L,Zhao Y G,et al. Chemical degradation of a Ferralsol (Oxisol) under intensive rubber (Hevea brasiliensis)farming in tropical China[J]. Soil & Tillage Research,2007,93(1): 109-116.
[5]Werner C,Zheng X H,Tang J W,et al. N2O,CH4and CO2emissions from seasonal tropical rain forests and a rubber plantation in Southwest China[J]. Plant and Soil,2006,289(1-2):335-353.
[6]Li H M,Aide T M,Ma Y X,et al. Demand for rubber is causing the loss of high diversity rain forest in SW China [J]. Biodiversity and Conservation,2007,16(6): 1731-1745.
[7]Li H M,Thomas M A,Ma Y X,et al. Past,present,and future land-use in Xishuangbanna,China and the implications for carbon dynamics[J]. Forest Ecology and Management,2008,255(1): 16-24.
[8]Ziegler A D,Fox J M,Xu J C. The rubber juggernaut[J]. Science,2009,324(5930): 1024-1025.
[9]Li H M,Ma Y X,Liu W J,et al. Clearance and fragmentation of tropical rain forest in Xishuangbanna,SW,China[J]. Biodiversity and conservation,2009,18(13): 3421-3440.
[10]周 宗,胡绍云. 橡胶产业对西双版纳生态环境影响初探[J].环境科学导刊,2008,27(3): 73-75.
[11]周 外,吴兆录,何謦成,等. 橡胶种植与饮水短缺: 西双版纳戈牛村的案例[J]. 生态学杂志,2011,30(7): 1570-1574.
[12]Tan Z H,Zhang Y P,Song Q H,et al. Rubber plantations act as water pumps in tropical China[J]. Geophysical Research Letters,2011,38:24406.
[13]喻 彦,蒙桂云,张利才.西双版纳地区近45年来气候变化及对热带作物的影响[J].热带农业科技,2007,30(3): 48-52.
[14]喻 彦,蒙桂云,张利才. 西双版纳地区近45年来气候变化特征[J]. 气象科技,2008,36(4): 410-413.
[15]陈 斌,张桃林.低丘红壤区农林间作系统的水分生态特征及生产力 [J].生态学杂志,1997,16(4): 1-5.
[16]张劲松,崔国栋. 农林复合系统的水分生态特征研究述评 [J].世界林业研究,2003,6(1): 10-14.
[17]Nair V D,Graetz D A. Agroforestry as an approach to minimizing nutrient loss from heavily fertilized soils: The Florida experience [J].Agroforestry Systems,2004,61(1): 269-279.
[18]Mcneely J A,Schroth G. Agroforestry and Biodiversity Conservation-Traditional Practices,Present Dynamics,and Lessons for the Future [J]. Biodiversity and Conservation,2006,15(2): 549-554.
[19]Roshetko J M,Lasco R D,Angeles M S. Smallholder agroforestry systems for carbon storage[J]. Mitigation and Adaptation Strategies for Global Change, 2007,12(2): 219-242.
[20]Dossa E L,Fernandes E C,Reid W S,et al. Above and below ground biomass,nutrient and carbon stocks contrasting an opengrown and a shaded coffee plantation [J]. Agroforestry Systems,2008,72(2): 103-115.
[21]Ashley R,Russell D,Swallow B. The policy terrain in protected area landscapes: challenges for agroforestry in integrated landscape conservation [J]. Biodiversity and Conservation,2006,15(2): 663-689.
[22]黎 蕾,蔡传涛,刘贵周. 光强和施氮量对催吐萝芙木生长及生物量的影响[J]. 中国生态农业学报,2010,9(1): 57-61.
[23]张丽霞,郭绍荣,李学兰,等. 催吐萝芙木的主要虫害及其防治[J]. 中药材,2006,(12): 1276-1278.
[24]李 琳,何红平,周 华,等. 催吐萝芙木中生物碱的研究(英文)[J]. 天然产物研究与开发,2007, (2): 235-239.
[25]Dutta P,Chopra I,Kapoor L. Cultivation of Rauvol fi a serpentine in India [J]. Economic Botany,1963,17(4): 243-251.
[26]赵永生,周亚兴,查云盛.云南萝芙木引种驯化栽培及发展前景[J]. 中国民族民间医药杂志,2006,(3): 179-180.
[27]符智建.珍贵树种——花梨木[J]. 农村新技术,2004,(8): 30.
[28]邱治军,周光益,陈升华. 海南特有珍贵红木树种——降香黄檀[J]. 林业实用技术,2004,(6): 41-42.
[29]倪 臻,王凌晖,吴国欣,等. 降香黄檀引种栽培技术研究概述[J]. 福建林业科技,2008,( 2): 265-268.
[30]庞家平,陈明勇,唐建维,等. 橡胶-大叶千斤拔复合生态系统中的植物生长与土壤水分养分动态[J]. 山地学报,2009,(4):433-441.
[31]唐建维,庞家平,陈明勇,等. 西双版纳橡胶林的生物量及其模型[J]. 生态学杂志,2009,28(10): 1942-1948.
[32]高国治,王明珠,张 斌.低丘红壤南酸枣-花生复合系统物种间水肥光竞争的研究——南酸枣与花生利用光能分析[J].中国生态农业学报,2004,12(2):92-94.
[33]王兴祥,张 斌,王明珠,等. 低丘红壤复合农林系统光能竞争与生产力—以花生南酸枣间作为例[J].生态学杂志,2002,21(4): 1-5.
[34]裴保华,贾渝彬,王文全,等. 杨农间作田的光强和土壤水分状况及其对农作物的影响[J]. 河北农业大学学报,1998,21(2):28-33.
[35]丁瑞兴. 茶园间作乌桕的气候生态效应[J]. 应用生态学报,1992,3(2): 131-137.
[36]王丽娟,朱兴正,毛加梅,等.不同遮荫树种对茶园土壤和茶叶品质的影响[J].中南林业科技大学学报,2011,31(8):66-73.
[37]汪汇海,李德厚.胶茶人工群落在改善山地土壤生态环境上的作用[J]. 山地学报,2003,21(3): 318-323.
[38]张墨谦,周可新,薛达元. 种植橡胶林对西双版纳热带雨林的影响及影响的消除[J]. 生态经济:学术版,2007,(02):377-378,439.
[39]张 敏,邹晓明. 热带季节雨林与人工橡胶林土壤碳氮比较[J].应用生态学报,2009,(05):1013-1019.
[40]王玉娟,雷小林,周文才,等.间种对林地土壤理化性质及树体生长的影响研究概述[J]. 湖北林业科技,2011,(4): 40-43.
[41]曹建华,梁玉斯,蒋菊生. 胶—农复合生态系统对橡胶园小环境的影响[J]. 热带农业科学,2008,(1): 1-8.
[42]梁艳丽,李 建,岩 对,等. 橡胶树与魔芋间作对魔芋净光合速率及产量的影响[J]. 长江蔬菜,2010,24: 35-38.
[43]萧自位,王秀华,郑 丽,等. 西双版纳不同胶林复合生态系统的生物量及其分配特征[J]. 中南林业科技大学学报,2014,34(2): 108-116.
[44]杨桂明. 推行胶桂间作提高橡胶树防灾减灾能力[J]. 中国热带农业,2009,(5): 41-42.
[45]范鸿雁,何 舒,罗志文,等. 幼龄橡胶园间作香蕉高效栽培模式研究[J]. 中国南方果树,2014,(2): 68-70.
[46]Yang Yusheng,Chen Guangshui,Yu Xintuo. Soil fertility in agroforestry system of Chinese fir and villous amomum in subtropical China [J]. Pedosphere,2001,11(4): 341-348.
[47]刘世荣. 沙棘对中国亚湿润干旱区杨树人工林生长与生产力的影响[J]. 植物生态学报,2000,24(2): 169-174.
[48]陈红跃,徐英宝.马尾松、藜蒴栲混交林生产力的研究[J].华南农业大学学报,1993,14(1): 144- 148.
[49]曹汉洋,陈金林. 杉木马尾松木荷混交林生产力研究[J]. 福建林学院学报,2000,20(2): 97-100.
[50]Forrester D I,Bauhus J,Cowie A L,et al. Mixed-species plantations of Eucalyptus with nitrogen- fi xing trees: A review[J].Forest Ecology and Management,2006,233: 211-230.
[51]Cairns M A,Olmsted I,Granados J,et al. Composition and aboveground tree biomass of a dry semi-evergreen forest on Mexico’s Yucatan Peninsula[J]. Forest Ecology and Management,2003,186:125-132.
[52]李 江,邱 琼,朱宏涛,等. 思茅松中幼龄人工林的生物量碳密度及其动态变化[J]. 中国水土保持科学,2011,(2):106-111.
[53]杜 虎,曾馥平,王克林,等. 中国南方3种主要人工林生物量和生产力的动态变化[J]. 生态学报,2014,(10):2712-2724.
[54]洪 滔,吴承祯,林勇明,等. 千年桐人工林乔木层的生物量特征[J]. 山地学报,2012,(6):648-654.
[55]林 文. 不同坡位7年生乐昌含笑人工林生物量分布研究[J].安徽农学通报(上半月刊),2012,(7):149-151,207.
[56]王 韧,李晓景,蔡金标,等. 闽东沿海秋茄天然林与人工林生物量比较[J]. 西南林学院学报,2010,(1):16-20.
[57]邵明晓,文仕知,何功秀,等. 不同林龄闽楠人工林生物量结构特征[J]. 中南林业科技大学学报,2014,34(6):44-48.
Dynamics of growth and biomass of two mixed rubber+Rauvol fi a vomitoria+Dalbergia odorifera systems with different stand ages
WANG Fei-jun1,2,CHEN Lei-xi3,LIU Cheng-gang2,LIU Chang-an2,TANG Jian-wei2
(1.University of Chinese Academy of Sciences,Beijing 100039,China; 2.Key Lab. of Tropical Plant Resources and Sustainable Use and Xishuangbanna Tropical Botanical Garden,Chinese Academy of Sciences,Mengla 666303,Yunnan,China; 3. College of Life Seience and Technology,Chinese Pharmaceutical University,Nanjing 211198,Jiangsu,China)
According to the survey data of two mixed rubber (with different tree ages)+Rauvol fi a vomitoria+Dalbergia odoriferain the agroforestry systems with 4-year-old and 7-year-old and three 20 m × 25 m plots,by adopting the author established biomass regression equations,the plant growth dynamics,biomass and distribution in two agroforestry systems with different stand ages were analyzed and calculated. The results show that the annual meanbasal diameter growth of R. vomitoria and D.odorifera in 4-year-old mixed rubber-tree system was signi fi cantly greater than that of growing in 7-year-old mixed rubber-tree system,while the annual mean height growth of R.vomitoria and D.odorifera in two mixed rubber-tree systems were no signi fi cant differences; After R.vomitoria and D.odorifera were planted for 4.5 years in 4-year-old and 7-year-old rubber plantations,the biomass of the former was 2.86 and 2.90 times as much as that of the latter,respectively; The total biomass (67.49 t·hm-2) of 4-year-old mixed rubber-tree system was signi fi cantly greater than that of the same years old rubber plantation (47.06 t·hm-2),as well as greater than that both of 7-year-old mixed rubber-tree system (63.86 t·hm-2) and rubber plantation (60.38 t·hm-2); Organ allocation pattern of biomass in two mixed rubber-tree systems and rubber plantations and trees was ranked as stem > root > branch > leaf,except the pattern of stem > branch > root > leaf for R.vomitoria in 4-year-old mixed rubber-tree system; The growth and biomass of R.vomitoria and D. odorifera declined while the biomass of rubber and the total biomass of mixed rubber-tree systems increased with the stand age,as well as revealed the advantage and potential for enhancing biomass accumulation and carbon sequestration of R. vomitoria and D. odorifera intercropped with rubber plantation.
agroforestry system; rubber plantation; Rauvol fi a vomitoria; Dalbergia odorifera; growth dynamics; biomass
S718.55+6;S794.1
A
1673-923X(2016)01-0086-08
10.14067/j.cnki.1673-923x.2016.01.015
2015-03-20
中国科学院农业项目办公室项目(KSCX2-EW-B-12;KSCX2-YW-N-44-09);西双版纳州科学技术局项目(200915;YX201102);中国科学院“一三五”专项(XTBG-T03)
王飞军,硕士研究生 通讯作者:唐建维,研究员,博士;E-mail:tangjw@xtbg.org.cn
王飞军,刘成刚,刘长安,等.不同年龄橡胶-催吐萝芙木-降香黄檀复合生态系统中植物的生长动态及其生物量[J].中南林业科技大学学报,2016,36(1): 86-93.
[本文编校:谢荣秀]
