柞栎象成虫触角感受器超微结构1)
2016-12-19王重舒李燕陈玉宝高文韬孟庆繁
王重舒 李燕 陈玉宝 高文韬 孟庆繁
(吉林市林木种苗管理站,吉林,132013) (北华大学) (吉林市林业科学研究院) (北华大学)
柞栎象成虫触角感受器超微结构1)
王重舒 李燕 陈玉宝 高文韬 孟庆繁
(吉林市林木种苗管理站,吉林,132013) (北华大学) (吉林市林业科学研究院) (北华大学)
为研究柞栎象(Curculiodentipes)寄主定位机制,利用环境扫描电镜对柞栎象触角形态、感受器的类型及分布特点等进行了观察。结果表明:柞栎象触角上有感受器5种类型10个亚型,分别为:刺形感受器2个亚型、毛形感受器4个亚型、锥形感受器、齿形感受器2个亚型和腔形感受器。柞栎象雌雄成虫触角性二型不明显,很难作为形态上性别鉴定的可靠依据。雄虫触角感受器数量多于雌虫,但差异不显著。柞栎象触角感受器从基部至端部(柄节至棒节第4亚节)有明显的分化趋势。
柞栎象;触角;感受器;环境扫描电镜
For the biological characteristics ofCurculiodentipes, we used environmental scanning electron microscopy (ESEM) to observe and describe the morphology of antennal sensilla on adults of both sexes. We observed ten different types of sensilla, two types of sensillachaetica, four types of sensillatrichodea, one type of sensillabasiconica, two types of sensilladentiform and one type of sensilla cavity. The differentiation trend of antennal sensilla was not obvious, and it tended to more refined from the base to the end of antennas. Antennae of males had more sensilla than those of females, but it is not significantly. There were obvious differentiation from base to top of sensilla.
昆虫触角感受器由昆虫体壁细胞特化而来,主要由毛原细胞、神经细胞和各种鞘细胞组成,是昆虫感知外界环境与内部反应,接收各类物理、化学信息的器官[1-9]。昆虫的寄主定位、取食、交配和产卵定位等行为都与触角感受器有着密切的联系。由于触角通常是昆虫感受器最为集中最为密集的着生器官。由此,大量学者对其进行了研究并取得了一定的成果。杨燕等[10]发现云南木蠹象(Pissodesyunnanensis)触角存在双芽形感器、耳形感器、毛形感器、锥形感器、脚形感器、芽形感器、端指形感器和腔形感器8种感器。常立儒等[11]在杨干象(Cryptorhynchuslapathi)触角上发现刺形感器、毛形感器、Böhm氏鬃毛、锥形感器、钟形感器5种感器类型。Yu et al.[12]对沟眶象(Eucryptorrhynchuschinensis)和臭椿沟眶象(E.brandti)触角感器种类和分布差异进行了研究,推测2个物种对寄主挥发物感知的差异与他们触角感受器的种类、分布差异有关。
柞栎象(Curculiodentipes)是蒙古栎(Quercusmongolica)种实的重要害虫,果实被害率为45%~55%,最高可达80%,严重时种仁被害一空[13]。由于柞栎象幼虫寄生于蒙古栎果实内部,危害极为隐蔽,严重影响了蒙古栎果实生产淀粉的经济效益。目前对柞栎象触角感受器的研究十分有限,利用环境扫描电镜对柞栎象触角上的感受器类型和分布进行细致统计,为下一步研究柞栎象寄主定位机制、针对柞栎象虫害制定综合防治手段提供基础研究。
1 材料与方法
1.1 试验材料
供试昆虫:选择触角完整的柞栎象雌雄各10只,摘取触角备用,供试柞栎象取自北华大学林学院昆虫实验室2013年室内饲养获取的羽化成虫。室内饲养的幼虫采自吉林市林业科学研究院松花湖实验基地。
1.2 样品制备
将雌雄成虫干制标本编号,分别放入培养皿中,加入4%的Triton X-100溶液浸泡24 h,用75%和95%的乙醇溶液逐级脱水各5 min,然后将样品转移至正己烷溶液中,浸泡2 h,并超声降解60 s,取出样品,自然风干24 h,待用。
取成虫触角,并进一步编号。每只成虫的触角按腹、背面分别粘在粘有导电胶布的铝片载物台上(圆形,Φ=4 cm),用SC7620离子溅射仪(Quorum,英国)进行喷镀,然后在20.0 kV加速电压下,利用环境扫描电镜(FEI,荷兰,Quanta200-D7972)进行表面观察。
1.3 统计方法
以单位面积内各类型感受器数量来反映触角感受器的总体情况。根据电子显微镜照片的比例尺,利用Photoshop CS6软件截取2 500 μm2(50 μm×50 μm)的正方格,进行统计分析。
2 结果与分析
2.1 触角的基本形态和分化趋势
柞栎象触角成膝状,表面光滑,从基部向端部依次分为柄节(Sc)、索节(Fu)和棒节(Cl)共3节(12亚节)组成,各节成圆锥形,连接处明显膨大。索节由7个亚节组成并向端部逐渐变粗变短,末端棒节共4个膨大亚节(图1)。在触角着生处的后下方,喙的表面具有一条触角沟,当触角缩回时,柄节位于触角沟内。触角基部呈球状,着生在一个臼形触角窝中。触角背面、腹面及各节连接处皆有感受器分布。感器从基部至端部(柄节至棒节第4亚节)有着明显的分化趋势。感器类型由粗糙逐渐趋于精细,由庞大逐渐趋于微小,由稀疏逐渐趋于致密。

图1 柞栎象触角
柞栎象雌雄成虫触角各亚节长度也存在着一些细微差异,除Cl.2、Cl.3外,雄虫触角各亚节长度均大于雌虫(表1)。

表1 柞栎象雌雄成虫触角各亚节长度 μm

性别Fu.5Fu.4Fu.3Fu.2Fu.1Sc雌333.75±0.96333.78±0.34431.52±6.23577.85±43.92324.48±21.702521.99±19.96雄416.17±86.97424.13±48.30511.17±73.77617.93±91.18750.19±97.242220.96±332.97
注:表中数据为平均值±标准误。
2.2 触角感受器类型
根据扫描电镜观察发现触角上的感受器共有5种类型10个亚型,分别为:
(1)刺形感器(Ch)。柞栎象成虫触角上分布最多的一种感器。外形为毛刺状,基部有基窝,外壁光滑无孔,顶端形态存在分化,有的顶端尖利,有的顶端分叉。刺形感器分布在触角的每一节,有2个亚型。
刺形感器Ⅰ(Ch.1):感器表面光滑,分布在触角棒节,且较均匀(图2A)。
刺形感器Ⅱ(Ch.2):基部到端部逐渐变细直至尖利,端部分叉或不分,略向触角表面弯曲,几乎贴附于触角表面。主要分布在柄节(图2E)。
(2)毛形感器(Tr)。基部到端部逐渐变细,端部钝圆。表面形态存在分化,从触角端部至基部纵脊从无到有逐渐增多。
毛形感器Ⅰ(Tr.1):柞栎象触角上最长的一种感器,表面光滑(图2A)。着生于触角棒节每个亚节靠近端部位置。
毛形感器Ⅱ(Tr.2):分布较多的感器类型之一,表面光滑,无纵脊,从基部至端部逐渐变细,端部急剧变细至钝圆(图2A),主要分布在Cl.2、Cl.3和Cl.4。
毛形感器Ⅲ(Tr.3):表面光滑,无纵脊。端部急剧变细,500倍下观察似尖刺,20 000倍下观察可见端部为明显的钝圆状(图2B),主要分布在Fu.5、Fu.6、Fu.7,Fu.1和Fu.2少量分布。
毛形感器Ⅳ(Tr.4):表面光滑,有明显纵脊,中部略膨大成纺锤形,端部钝圆(图2C),主要分布在Fu.1、Fu.2、Fu.3和Fu.4。
(3)锥形感器(Ba)。成圆锥形,基部到端部逐渐变细,端部钝圆,基部有突起的基座(图2A)。参考杨贵军等的研究,将3种形态类似,功能相同的Ba.合并为一个类型[15-16],笔者将这些类似的Ba.作为一个类型统计。主要分布在Cl.2、Cl.3和Cl.4,Cl.1少量分布。
(4)齿形感器(De)。
齿形感器Ⅰ(De.1):表面光滑,有明显纵脊,端部分叉为两个或更多(图2B),分布在索节各亚节靠近端部的位置。
齿形感器Ⅱ(De.2):表面光滑,有明显纵脊(图2C),少量分布在Fu.1、Fu.2、Fu.3、Fu.4和Sc中部。
(5)腔形感器(Co)。圆孔形凹腔(图2D),仅在Sc观察到。
2.3 雌雄成虫触角感受器的分布差异
通过扫描电镜观察,柞栎象雌雄成虫触角感器总体数量分布具有细微差异,其中单位面积上雄虫触角各亚节的感器数量均略高于雌虫(表2)。

Tr.1.毛形感受器Ⅰ;Tr.2.毛形感器Ⅱ;Ch.1.刺形感器Ⅰ;Ba.锥形感器;Tr.3.毛形感器Ⅲ;De.1.齿形感器Ⅰ;Tr.4.毛形感器Ⅳ;De.2.齿形感器Ⅱ;Co.腔形感器;Ch.2.刺形感器Ⅱ。
图2 柞栎象触角感受器类型
柞栎象雌雄成虫触角上感器种类分布差异较为明显的有:索节的Fu.2,雌虫主要分布齿形感器Ⅱ(De.2),毛形感器Ⅲ(Tr.3)和毛形感器Ⅳ(Tr.4)极少分布,雄虫则3种感器分布比较平均,仅为不同类型感器数量比例上的差异;Fu.3,雌虫只分布毛形感器Ⅳ(Tr.4),而雄虫上除主要分布毛形感器Ⅳ(Tr.4)外还分布少量齿形感器Ⅱ(De.2);Fu.4雌雄虫的差异同Fu.3;Fu.7则是雌虫的齿形感器Ⅰ(De.1)端部为2个齿形凸起,而雄虫的齿形感器Ⅰ(De.1)端部为多个齿状凸起。
棒节上较明显的差异存在于Cl.3和Cl.4。Cl.3雌虫刺形感器Ⅰ(Ch.1)和毛形感器Ⅱ(Tr.2)分布较为平均,锥形感器(Ba)分布较雄虫少切不均匀,而雄虫则是以刺形感器Ⅰ(Ch.1)为主,锥形感器(Ba)较为均匀地着生于刺形感器Ⅰ(Ch.1)之下;Cl.4雌虫为毛形感器Ⅱ(Tr.2)多于刺形感器Ⅰ(Ch.1),而雄虫则是刺形感器Ⅰ(Ch.1)多于毛形感器Ⅱ(Tr.2),仅是不同感器分布比例的细微差别。
由此可见,虽然在棒节(Cl)雌雄虫感器分布的差异仅仅是不同种类之间比例的差异,但这种微小的差异反映出的是雌雄虫触角端部精细感器比例多少的差异。

表2 柞栎象雌雄成虫触角感器数量与分布 个
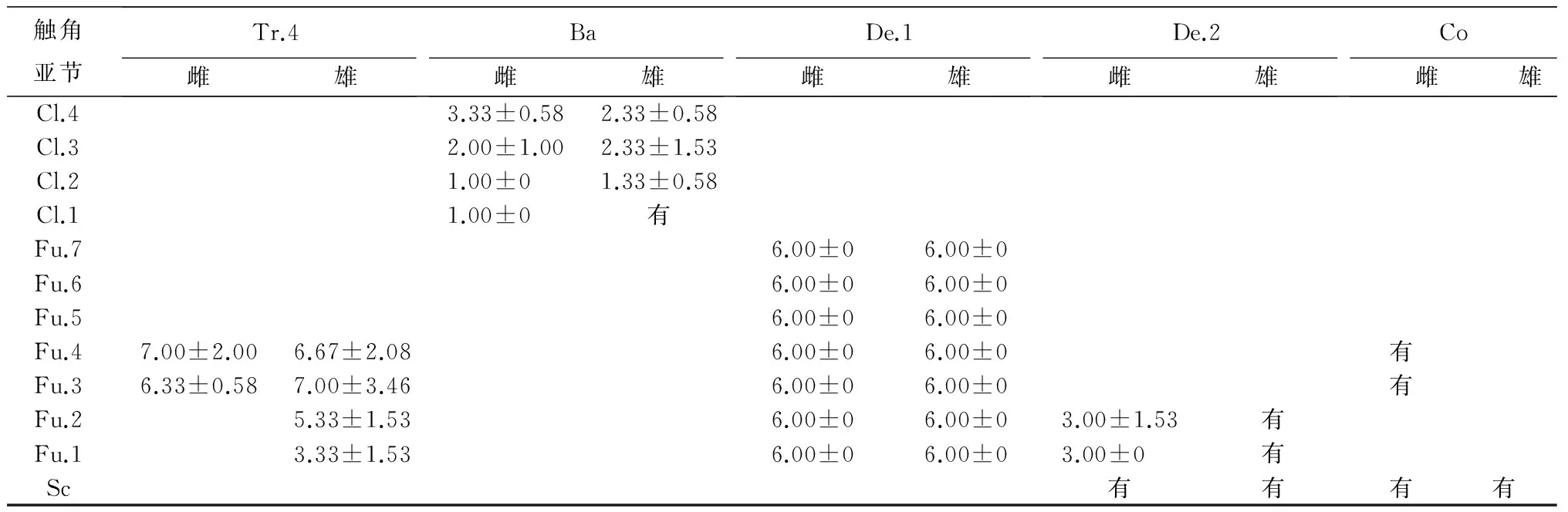
触角亚节Tr.4雌雄Ba雌雄De.1雌雄De.2雌雄Co雌雄Cl.43.33±0.582.33±0.58Cl.32.00±1.002.33±1.53Cl.21.00±01.33±0.58Cl.11.00±0有Fu.76.00±06.00±0Fu.66.00±06.00±0Fu.56.00±06.00±0Fu.47.00±2.006.67±2.086.00±06.00±0有Fu.36.33±0.587.00±3.466.00±06.00±0有Fu.25.33±1.536.00±06.00±03.00±1.53有Fu.13.33±1.536.00±06.00±03.00±0有Sc有有有有
注:统计面积为2 500 μm2。表中数据为平均值±标准误。
3 结论与讨论
柞栎象触角感受器与之前学者们研究象甲科其他昆虫触角感受器形态相似,由此推断其功能也相似,其中毛形感器(Tr)表面光滑,没有腺孔结构,之前的学者研究猜测此类感器只是一种机械感受器,或者仅仅是起到保护作用的毛状结构[13]。De.1是柞栎象触角上最长、最粗壮的感器,是触角与物体发生接触行为时首先触碰物体的感受器。以前的研究表明,De.1可能具有机械触感和化学感应双重功能[14-15]。本研究发现的腔形感器(Co)中未观察到内部结构(如突起),由此判断此类型感器为触角表皮腺孔[16],Zacharuk提出此类感器为嗅觉感器的假设[17],并在2002年被Lopes et al.证实[18]。
柞栎象雌雄成虫触角性二型不明显,只有在感器分化程度上有细微差别,因此很难作为形态学上性别鉴定的可靠依据。
柞栎象触角上着生的感器从基部至端部(柄节至棒节第4亚节)有着明显的分化趋势。感器类型由粗糙逐渐趋于精细,由庞大逐渐趋于微小,由稀疏逐渐趋于致密。
虽然在棒节(Cl)雌雄虫感器分布的差异仅仅是不同种类之间比例的差异,但这种微小的差异反映出的是雌雄虫触角端部精细感器比例多少的差异。由以上分析可以推测:雄虫触角端部精细感器的比例高于雌虫,这可能使柞栎象雌虫活跃度低于雄虫。但这个推测还需进一步证实。
柞栎象雌雄成虫触角感器分布的种类与本科其他属昆虫有着明显的差异。杨燕等发现云南木蠹象触角存在双芽形感器、耳形感器、毛形感器、锥形感器、脚形感器、芽形感器、端指形感器和腔形感器[10],其中耳形感器和芽形感器未在柞栎象触角上发现,但柞栎象触角着生的齿形感器则未在云南木蠹象触角上发现。杨干象具有刺形感器、毛形感器、Böhm氏鬃毛、锥形感器、钟形感器5种感器类型[11],而柞栎象上无Böhm氏鬃毛分布,而有齿形感器、腔形感器感器分布。
感器的种类和功能在柞栎象对寄主定位和寻找配偶获得生殖机会的过程中具有重要意义,本研究也为柞栎象干扰性和聚集性新型制剂的研制提供了科学依据。
[1] 余海忠.昆虫触角感受器研究进展[J].安徽农业科学,2007,35(14):4238-4240,4243.
[2] 马瑞燕,杜家纬.昆虫的触角感器[J].昆虫知识,2000,37(3):179-183.
[3] SCHNEIDER D. Insect antennae[J]. Annu Rev Entomol,1964,9:103-122.
[5] HALLBERG E, HANSSON B S, STEINBRECHT R A. Morphological characteristics of antennal sensilla in the European cornborerOstrinianubilalis(Lepidoptera: Pyralidae)[J]. Tissue Cell,1994,26(4):489-502.
[6] HUNGER T, STEINBRECHT R A. Functional morphology of adouble-walled multiporous olfactory sensillum: the sensillum coeloconicum ofBombyxmori(Insecta: Lepidoptera)[J]. Tissue and Cell,1998,30(1):14-29.
[7] SHIELDS V D C, HILDEBRAND J G. Fine structure of antennal sensilla of the female sphinx moth,Manducasexta(Lepidoptera: Sphingidae). II. Auriculate, coeloconic, and styliform complex sensilla[J]. Canadian Journal of Zoology,1999,77(2):302-313.
[8] ZIMMERMANN B. Differentiation of the thermo-/hygrosensitive (no-pore) sensilla on the antenna ofAntheraeapernyi(Lepidoptera, Saturniidae): a study of cryofixed material[J]. Cell Tissue Res,1991,266:427-440.
[9] WALTERS B D, ZACHARUK R Y. Morphology and ultrastructure of sensilla on the proboscis of the adult spruce budworm,Choristoneurafumiferana(Clem.)(Lepidoptera: Tortricidae)[J]. Candian Journal of Zoology,1998,76(3):466-479.
[10] 杨燕,杨茂发,杨再华,等.云南木蠹象触角感器的扫描电镜观察[J].林业科学,2009,45(2):72-78.
[11] 常立儒,张璐,王涛,等.杨干象触角感器的扫描电镜观察[J].植物检疫,2014,28(3):23-29.
[12] YU Q Q, LIU Z K, CHEN C, et al. Antennal sensilla ofEucryptorrhynchuschinensis(Olivier) andEucryptorrhynchusbrandti(Harold) (Coleoptera: Curculionidae)[J]. Microscopy Research and Technique,2013,76:968-978.
[13] 裴建国.栎实象生物学特性及防治措施研究[J].安徽农学通报,2011,17(8):121-122.
[14] 杨贵军,雍惠莉,王新谱.沟眶象的生物学特性及行为观察[J].昆虫知识,2008,45(1):65-69.
[15] 杨贵军,张大治,孙晶莹.沟眶象触角感器的扫描电镜观察[J].昆虫知识,2008,45(6):926-931.
[16] BLAND R G. Antennal sensilla of the adult alfalfa weevilHyperapostica(Gyllenhal) (Coleoptera: Curculionidae)[J]. Int J Insect Morphol Embryol,1981,10(3):265-274.
[17] ZACHARUK R Y. Ultrastructure and function of insect chemosensilla[J]. Annu Rev Entomol,1980,25(1):27-47.
[18] LOPES O, BARATA E H, ARAUJO J, et al. Fine structure of antennal sensilla basiconica and their detection of plant volatiles in the eucalyptus woodborer,PlcoracanthasemipunctataFabricius (Coleoptera: Cerambycidae)[J]. Arthropod Struct Dev,2002,31(1):1-13.
Ultrastructure in Antennal Sensilla ofCurculiodentipes//
Wang Chongshu; Li Yan; Chen Yubao; Gao Wentao, Meng Qingfan
(Seeding management station of Jilin, Jilin 132013, P. R. China)(Beihua University)(Forestry Science Research Institute of Jilin)(Beihua University)//Journal of Northeast Forestry University,2016,44(11):92-95.
Curculiodentipes; Antennae; Sensilla; Environmental scanning electron microscopy (ESEM)
王重舒,男,1986年12月生,吉林市林木种苗管理站,工程师。E-mail:35409763@qq.com。
孟庆繁,北华大学林学院,教授。E-mail:qingfanmeng@sina.com。
2016年1月27日。
S763.301
1)吉林省科技支撑项目(20130206056NY)。
责任编辑:程 红。
