三环泡湿地保护区浮游植物功能群季节演替及影响因子1)
2016-12-19马成学尹子龙于洪贤
马成学 尹子龙 于洪贤
(东北林业大学,哈尔滨,150040)
三环泡湿地保护区浮游植物功能群季节演替及影响因子1)
马成学 尹子龙 于洪贤
(东北林业大学,哈尔滨,150040)
为了研究寒区湿地保护区浮游植物功能群的季节变化以及影响因子,于2014年春季(5月份)、夏季(7月份)和秋季(9月份)对三环泡湿地保护区15个采样点进行了调查分析。结果表明:三环泡湿地保护区共划分为20个功能群,其中重要功能群为X2、W1、MP、H1、Y和C,呈现X2+W1→X2+MP→Y+X2+C季节变化特征。通过RDA多元统计分析表明,水温、总氮质量浓度、透明度、浊度、总无机碳质量浓度是影响寒区浮游植物功能群的主要环境因子。
浮游植物;功能群;三环泡湿地保护区
We studied the relationship between the phytoplankton functional groups and associated environmental factors in cold region of China. We investigated the spatial and temporal distribution characteristics of phytoplankton functional groups in Sanhuanpao Wetland Reserve from May to September 2014. There were 20 functional groups in Sanhuanpao Wetland Reserve during the study periods and the dominant groups included functional groups H1, MP, C, Y, X2 and W1. Group W1 and X2 were predominant in the spring associated with high water temperature (WT) and nutrients availability. Group X2, MP and H1 were associated with high WT, nutrients availability in summer. Group Y, X2 and C were associated with low WT and nutrients the most important factors, WT was negatively related with groups Y and C, and positively related availability in the autumn. The redundancy analysis (RDA) showed that WT, TN, SD, and IC were with other groups.
水生态系统中浮游植物、浮游动物、底栖动物和鱼类等同水环境关系密切,其中浮游植物同水环境的关系极为密切,水污染通常导致水体中浮游植物群落结构变化显著[1-3]。浮游植物是水生态系统中的主要初级生产力,其群落结构的变化直接或间接影响浮游动物、底栖动物和鱼类等其他水生生物的群落结构,进而影响水生态系统的功能。湿地水生态系统是湿地生态系统的重要组成部分,湿地水生态系统的健康直接影响整个湿地生态系统的健康。湿地保护区通常远离城市,紧邻乡村,但近年来常常受到农田废水、城市污水和旅游等人为干扰,水质明显下降,水体富营养化严重,蓝藻爆发的现象时有发生[4],因此,对浮游植物的监测显得尤为重要。传统的监测方法往往注重群落结构和多样性的分析,难以体现浮游植物的生态功能[5],因此,Reynolds et al.[6]和Padisak et al.[7]提出了浮游植物功能群的概念,并将浮游植物划分为39个功能群。该方法在国外研究较多,在国内主要应用于南方的河流、水库和高原湖泊中[8-11],在北方寒冷地区,尤其是湿地保护区研究极少[12]。三环泡国家级湿地保护区保护力度较大,严格控制人为活动干扰,没有进行任何旅游的开发利用,但是近年来由于上游煤矿废水、周围农田废水以及挠力河、七星河污水的影响,三环泡湿地保护区部分水域水质明显恶化,水体趋于富营养化。文中根据浮游植物功能群理论,针对三环泡湿地保护区,探讨寒区湿地保护区浮游植物功能群季节变化以及对保护区的指导意义,为北方寒区湿地保护区综合管理提供依据。
1 研究地概况
三环泡湿地保护区为三江平原典型湿地分布区之一,位于黑龙江省三江平原腹地,七星河中下游,富锦市境内南部,距富锦市区75 km。保护区南与宝清县、西与友谊县相邻,西南部与七星河国家级自然保护区隔七星河相望,东南部隔挠力河与挠力河国家级自然保护区相接。保护区的主要保护对象是内陆湿地生态系统及其栖息的珍稀水禽,共有高等植物78科236属415种,鱼类6目11科42种,两栖类2目4科6种,爬行类2目3科5种,鸟类17目40科217种,兽类6目11科30种。其担负着湿地生物多样性保护、实现湿地生态系统的保护和可持续发展的重要任务。
2 材料与方法
采样点设置:在2014年春季(5月份)、夏季(7月份)和秋季(9月份)对三环泡湿地保护区进行水样采集,根据三环泡湿地保护区核心区、缓冲区以及试验区规划以及周围的环境影响共设置了15个采样点(图1),其中S1位于友谊县、宝清县和富锦县三县交界处,S2~S7位于保护区东侧芦苇管护站核心区,S8位于保护区中部缓冲区,S9~S15位于保护区西侧大兴管护站核心区和缓冲区。
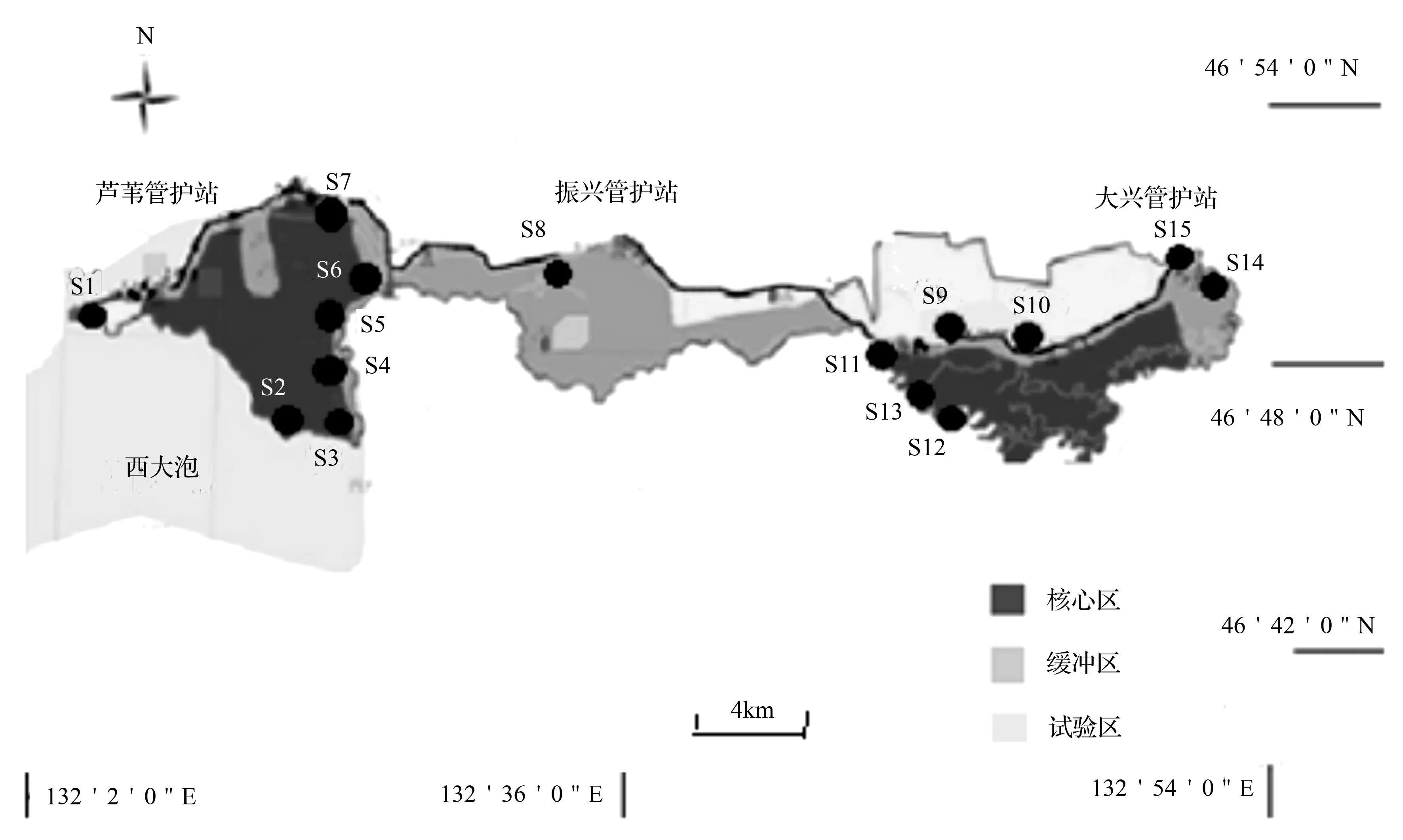
图1 三环泡湿地保护区浮游动物采样点

数据分析:浮游植物功能群各采样点生物量数据和水化学因子数据通过lg(x+1)进行处理使之更趋于正态分布后,首先采用Canoco for windows 4.5软件进行DCA分析,发现SD值<3,因此,对物种数据和相关环境因子数据进行RDA分析[17]。
3 结果与分析
3.1 三环泡湿地保护区水环境特征
三环泡湿地保护区水环境因子情况见表1,方差分析结果表明,除了电导率外,其他环境因子均呈现明显的季节变化,春季透明度、氨氮质量浓度、硝酸盐质量浓度、生物耗氧量和总有机碳质量浓度明显高于其他季节的,而氯离子、浊度、总氮质量浓度、总磷质量浓度、总碳质量浓度和总无机碳质量浓度则明显低于其他季节的;夏季氯离子质量浓度和总氮质量浓度较高;秋季总磷质量浓度和化学耗氧量明显高于其他季节,而水温(TE,P)和生物耗氧量明显低于其他季节。
3.2 三环泡浮游植物功能群的时空分布
三环泡湿地保护区通过采样分析发现浮游植物7门66种,可划分出20个不同的功能群(表2)。将各采样点相对生物量>20%的功能群定义为重要功能群,共有6个功能群,分别为MP、C、H1、Y、X2和W1。
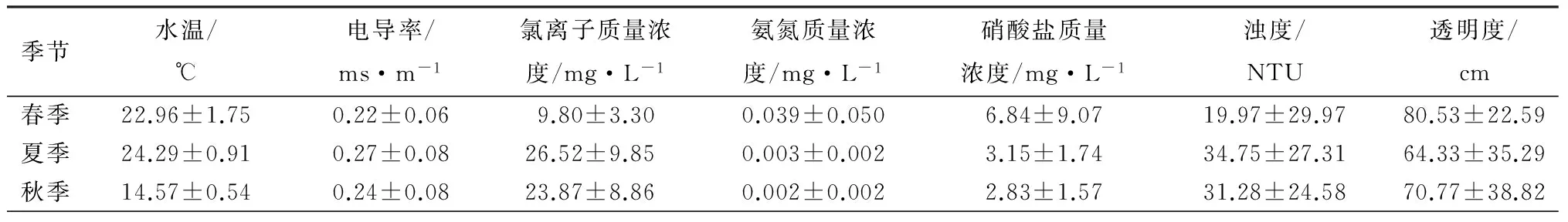
表1 三环泡湿地保护区各季节环境因子
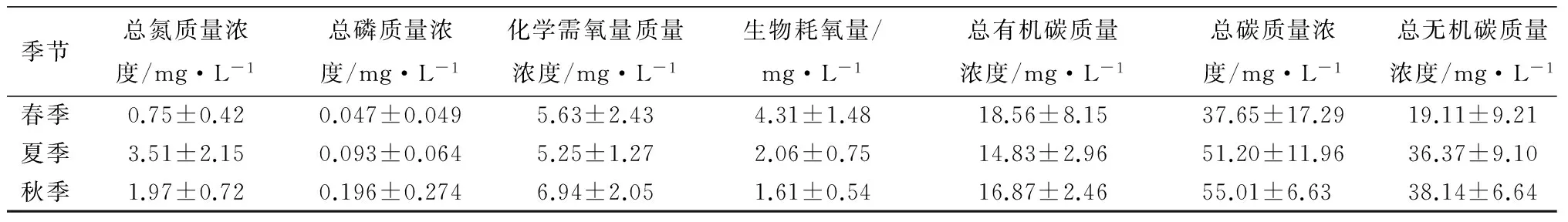
季节总氮质量浓度/mg·L-1总磷质量浓度/mg·L-1化学需氧量质量浓度/mg·L-1生物耗氧量/mg·L-1总有机碳质量浓度/mg·L-1总碳质量浓度/mg·L-1总无机碳质量浓度/mg·L-1春季0.75±0.420.047±0.0495.63±2.434.31±1.4818.56±8.1537.65±17.2919.11±9.21夏季3.51±2.150.093±0.0645.25±1.272.06±0.7514.83±2.9651.20±11.9636.37±9.10秋季1.97±0.720.196±0.2746.94±2.051.61±0.5416.87±2.4655.01±6.6338.14±6.64
注:表中数据为平均值±标准差。
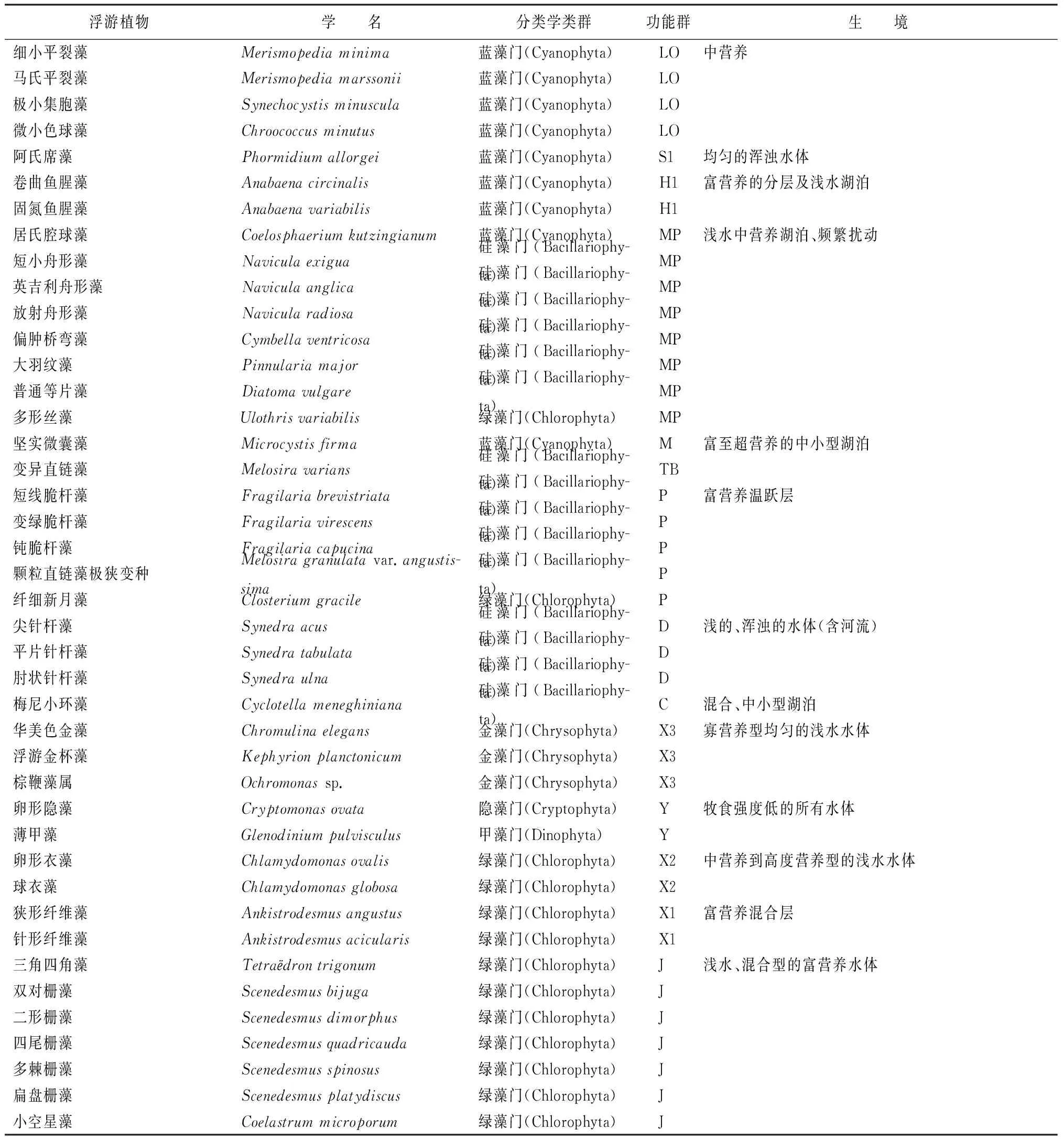
表2 三环泡湿地保护区浮游植物功能群

续(表2)
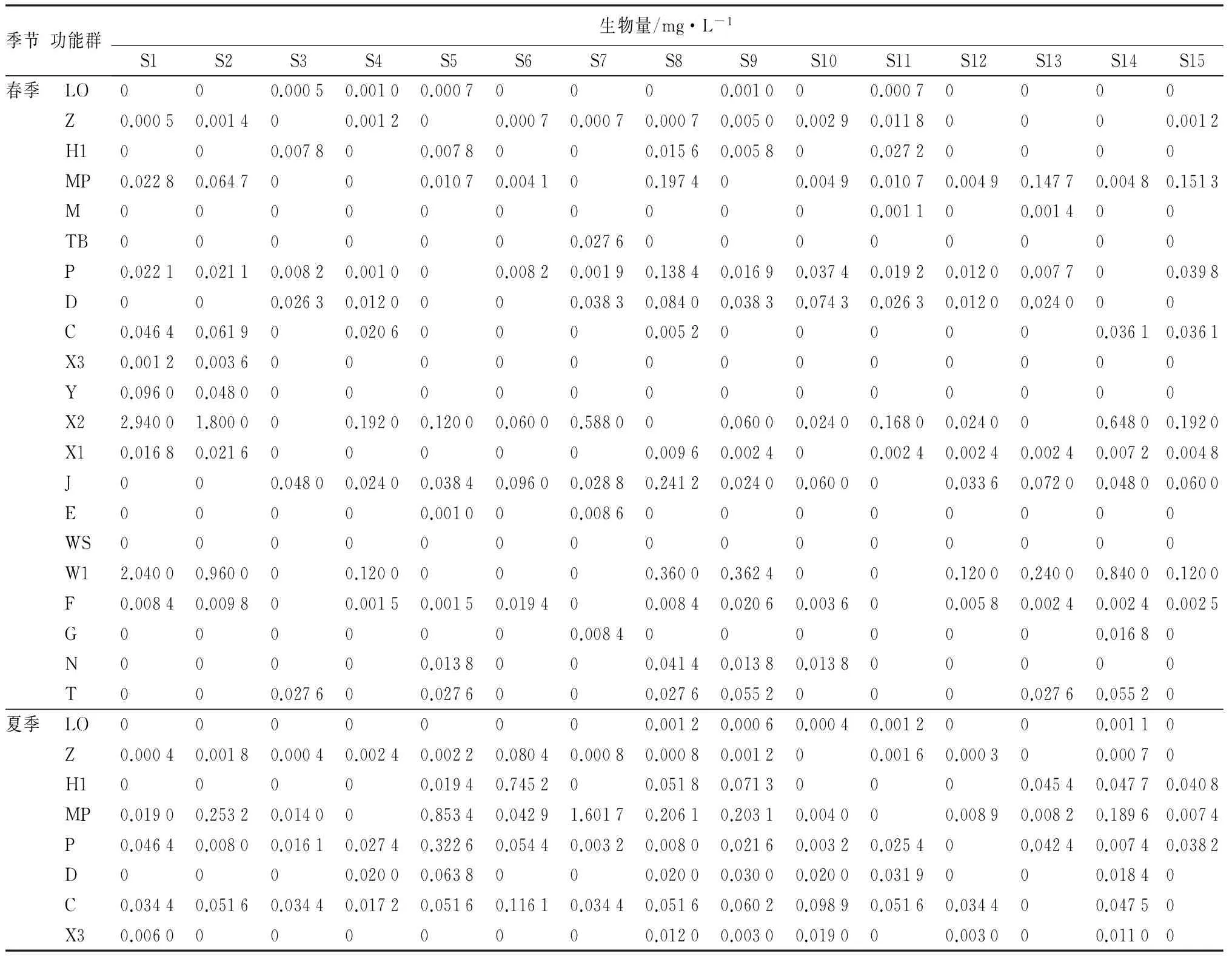
表3 各季节浮游植物功能群生物量水平分布
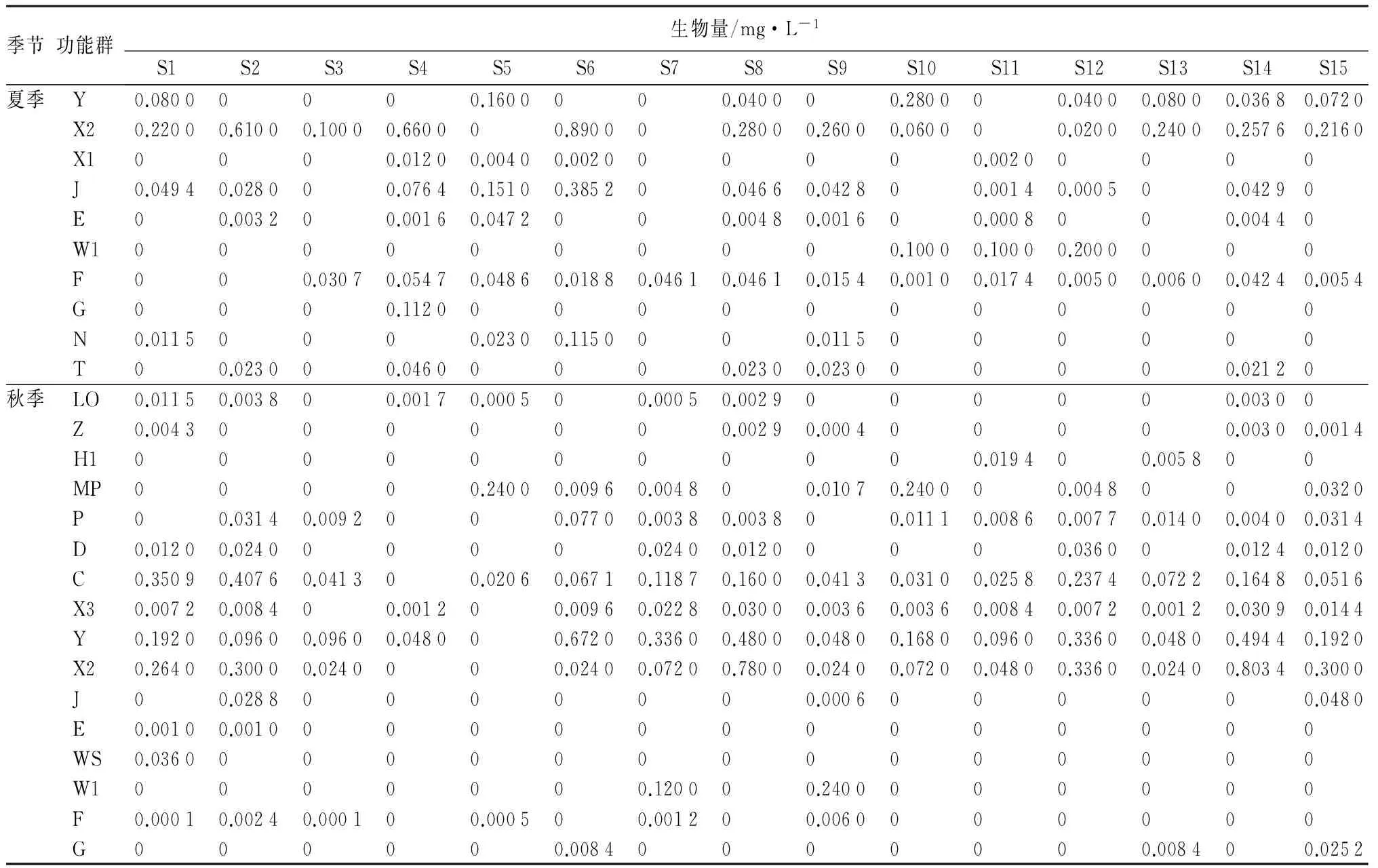
续(表3)
从表3中可以看出,春季三环泡浮游植物生物量主要以功能群X2和W1占优势,尤其是生物量最高的S1和S2采样点,功能群X2所占比例超过50%;夏季三环泡浮游植物生物量主要以功能群X2、MP和H1占优势,S5和S7以功能群MP占优势,S2和S4以功能群X2占优势,生物量最高的S6以功能群X2和H1占优势;秋季三环泡浮游植物生物量主要以功能群X2、Y和C占优势,S1和S2以功能群C占优势,S6和S7以功能群Y占优势,生物量最高的S8以功能群X2和Y占优势。根据各季节功能群平均相对生物量可以看出(表4),春季以功能群X2和W1占优势,分别为45.38%和34.37%;夏季以功能群X2和MP占优势,分别为29.90%和26.75%;秋季以功能群Y、X2和C占优势,分别为33.78%、31.41%和18.31%。
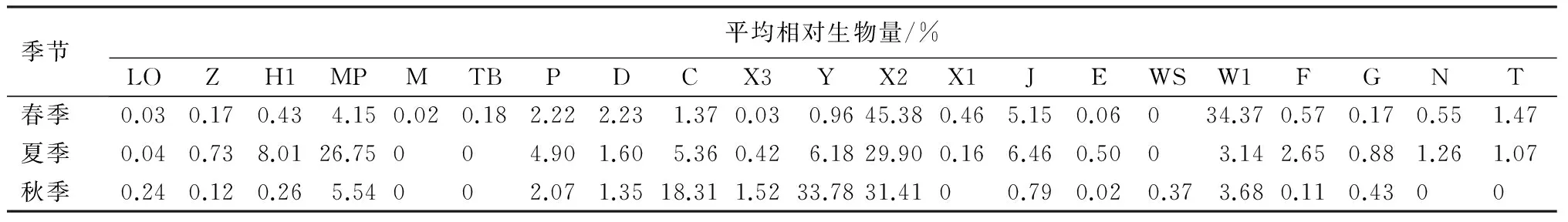
表4 三环泡湿地保护区浮游植物功能群季节平均相对生物量
3.3 浮游植物功能群同水环境因子的关系
不同水环境因子对浮游植物功能群影响不同。从图2可以看出,RDA结果表明,第一轴、第二轴共解释了属种累计方差值的21.6%,在第一轴,TN为最主要正相关因子(0.530),其次负相关因子为透明度(-0.452);在第二轴,水温为最主要负相关因子(-0.647),其次负相关因子为BOD5(-0.488),为最主要影响因子,最主要正相关因子为总无机碳质量浓度(0.470)。总氮质量浓度同绝大多数功能群呈正相关,而透明度恰恰相反,同绝大多数功能群呈负相关。重要浮游植物功能群Y、C同总无机碳质量浓度、浊度呈显著正相关,同水温、透明度呈显著负相关,而重要功能群MP、W1恰恰相反。水环境因子对浮游植物功能群E、P等分布影响不大。
4 结论与讨论
三环泡湿地保护区浮游植物功能群季节变化差异显著,呈现X2+W1→X2+MP→Y+X2+C转变的特征。说明功能群X2是最为重要、最有代表性的浮游植物功能群,与小兴凯湖浮游植物功能群研究结果一致,而同为寒区的安邦河湿地保护区未发现这一特点[12]。其主要原因是浮游动物牧食压力不同所致。安邦河湿地保护区水体中生长有大量的滤食性枝角类大型溞,浮游动物对浮游植物的牧食压力较大,其浮游植物功能群主要以J和P占优势。而小兴凯湖和三环泡湿地保护区浮游动物牧食压力较小,有利于功能群X2的生长[6],浮游植物均以功能群X2占优势。功能群Y由卵形隐藻和薄甲藻组成,主要代表牧食强度低的静水水体[18],三环泡湿地保护区秋季各采样点以功能群Y和功能群X2占优势,同样也说明了三环泡湿地保护区浮游动物牧食压力较小。功能群W1主要由尖尾裸藻组成,功能群W1主要代表从农田或污水中获得有机质的水体[18],说明功能群W1占优势的春季S1和S14采样点受到了农田废水和上游污水污染。S1采样点位于三县交界处,主要受到上游煤矿废水芳香烃类有机物的污染,S14采样点位于大兴农场附近,受农田废水影响较大。功能群MP主要由舟形藻属种类和多形丝藻组成,功能群MP主要代表经常受到扰动的、无机的、浑浊的水体[18],夏季是丰水期,水体扰动频繁,浊度明显增加,有利于功能群MP生长。功能群H1是由蓝藻门卷曲鱼腥藻和固氮鱼腥藻组成,是寒区湿地保护区蓝藻爆发的主要种类[19],仅在夏季S6占优势,S6采样点位于七星河河道上,可能受到上游七星河河水的污染,有潜在的蓝藻爆发趋势。除此之外,功能群C在秋季,尤其是S1和S2中占优势,功能群C代表富营养型的中小型水体,能够耐受低光照的影响[18],从表1可以看出,秋季透明度显著降低,浊度明显增加,光照明显降低,硅藻种类主要由能够耐受低光照的功能群C组成。
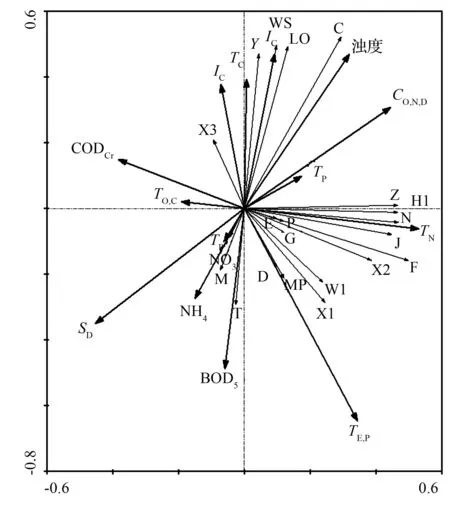
图2 三环泡浮游植物功能群RDA分析
温度、营养盐、水动力学和生物之间的相互作用通常是影响浮游植物功能群的主要影响因子[20]。热带、亚热带地区水温变化不如温带、寒带地区显著,但由于近年来气候变暖使水温上升积温增加也会导致亚热带地区出现水华提前爆发并且加剧的现象[21]。同热带、亚热带地区明显不同,水温变化通常是影响寒区湿地浮游植物功能群的最主要的环境因子[12],同时气候变暖引起的水温上升和积温增加也可能是近年来寒区湿地保护区蓝藻爆发的原因之一,因此,水温对寒区浮游植物功能群的影响变得越来越重要。在RDA图中,第二轴同水温呈显著负相关,温度由水温较低的秋季到水温较高的夏季逐渐升高,功能群也由秋季重要的功能群Y和C向夏季重要功能群MP和X2变化。功能群Y和C主要由耐低温的隐藻门、甲藻门和硅藻门种类组成,功能群MP和X2由喜高温的绿藻门种类组成[22],因此,功能群Y、C和MP、X2分别在秋、夏季节成为优势种。同武安泉等[12]对安邦河湿地保护区浮游植物功能群的研究结果较为一致。营养盐是影响浮游植物功能群的又一个主要影响因子,同其他寒区湿地保护不同,总氮是影响三环泡湿地保护区浮游植物功能群的主要影响因子。所有重要浮游植物功能区均与总氮呈正相关,尤其是功能群H1和X2,功能群H1是由蓝藻门卷曲鱼腥藻和固氮鱼腥藻组成,是寒区湿地保护区蓝藻爆发的主要种类,夏季三环泡湿地保护区N质量浓度/P质量浓度明显大于16,说明总氮是限制因子[23],因此,总氮是控制三环泡湿地保护区夏季蓝藻爆发的主要途径之一。三环泡湿地保护区透明度较高,泥沙和悬浮物含量较少,透明度主要是由浮游植物引起的,因此,同浮游植物功能群呈明显负相关,而功能群C同浊度呈显著性正相关,说明根据温带湖泊划分的功能群C同样适用于寒区湿地。碳是构成浮游植物的主要元素之一,浮游植物是碳循环的主要环节,以往学者普遍认为蓝藻是浮游植物固碳的主要功能类群,而英国学者最新研究结果表明,其他藻类的固碳作用同样不可小觑[24],约占浮游植物固碳总量的一半。无机碳对不同浮游植物的影响明显不同,杨威[25]等对天津新开湖的浮游植物和水中无机碳研究表明,无机碳与浮游植物没有明显的相关性。但在文中RDA图中可以看出,在第二轴,总无机碳质量浓度同浮游植物功能群Y和C呈显著正相关,同功能群MP和W1呈显著负相关。说明功能群Y和功能群C种类能够有效利用水体中的二氧化碳和碳酸盐类,固碳作用更强。过去学者普遍认为有机碳同浮游植物呈正相关,但相关不显著[26],在文中总有机碳也不是主要的影响因子。但中国科学院海洋研究所最新研究结果表明,有机碳明显有利于绿藻门小球藻的生长,对于其他门类浮游植物的影响还有待于进一步研究。
[1] 于洪贤,宋俊霖,马成学,等.小鹤立河水库及上游河流水域春季浮游植物特征[J].东北林业大学学报,2013,41(9):98-102.
[2] 于洪贤,张力文,马成学,等.小兴凯湖5月份浮游植物群落及其与环境因子的典范对应[J].东北林业大学学报,2014,42(11):95-97.
[3] 崔安琪,翟昊,于洪贤.宁夏沙湖浮游动物群落结构及多样性[J].东北林业大学学报,2015,43(9):121-124.
[4] 厉威池,马成学,郭志伟,等.虎林市月牙湖微囊藻水华爆发时及其后期的藻类丰度和毒素含量[J].水产学杂志,2014,27(3):12-17.
[5] YU H X, WU J H, MA C X, et al. Seasonal dynamics of phytoplankton functional groups and its relationship with the environment in river: a case study in northeast China[J]. Journal of Freshwater Ecology,2012,27(3):429-441.
[6] REYNOLDS C S, HUSZAR V, KRUK C, et al. Towards a functional classification of the freshwater phytoplankton[J]. Journal of Plankton Research,2002,24(5):417-428.
[7] PADISAK J, CROSSETTI L O, NASELLI-FLORES L. Use and misuse in the application of the phytoplankton functional classification: a critical view with updates[J]. Hydrobiologia,2009,621(1):1-19.
[8] 董静,李艳晖,李根保,等.东江水系浮游植物功能群季节动态特征及影响因子[J].水生生物学报,2013,37(5):836-843.
[9] 黄享辉,胡韧,雷腊梅,等.南亚热带典型中小型水库浮游植物功能类群季节演替特征[J].生态环境学报,2013,22(2):311-318.
[10] 高国敬,肖丽娟,林秋奇,等.海南省典型水库浮游植物功能类群的结构特征与水质评价[J].生态科学,2013,32(2):144-150.
[11] 黄国佳,李秋华,陈橼,等.贵州高原三板溪水库浮游植物功能群时空分布特征[J].环境科学学报,2015,35(2):418-428.
[12] 武安泉,郭宁,覃雪波.寒区典型湿地浮游植物功能群季节变化及其与环境因子关系[J].环境科学学报,2015,35(5):1341-1349.
[13] 黄翔飞,陈伟民,蔡启铭.湖泊生态调查观测与分析[M].北京:中国标准出版社,1999:27-105.
[14] 胡鸿钧,魏印心.中国淡水藻类:系统、分类及生态[M].北京:科学出版社,2006.
[15] 齐雨藻,李家英,谢淑琦,等.中国淡水藻志:第10卷:硅藻门羽纹纲[M].北京:科学出版社,2004.
[16] 国家环境保护总局.水和废水监测分析方法[M].北京:中国环境科学出版社,1997.
[17] XIAO L J, WANG T, HU R, et al. Succession of phytoplankton functional groups regulated by monsoonal hydrology in a large canyon-shaped reservoir in southern China[M]. Water Research,2011,45(16):5009-5019.
[18] 杨文,朱津永,陆开宏,等.淡水浮游植物功能类群分类法的提出、发展及应用[J].应用生态学报,2014, 25(6):1833-1840.
[19] 于洪贤,王尚,马成学.夏季大兴凯湖(中国一侧)沿岸带浮游植物群落的动态变化[J].水产学杂志,2014,27(3):1-5.
[20] MA C X, YU H X. Phytoplankton community structure in reservoirs of different trophic status, Northeast China[J]. Chinese Journal of Oceanology and Limnology,2013,31(3):1-11.
[21] DENG J M, QIN B Q, PAERL H, et al. Earlie rand warmer springs increase cyanobacterial (Microcystisspp.) blooms in subtropical Lake Taihu,China[J]. Freshwater Biology,2014,59(5):1076-1085.
[22] CHEN Q W, ZHANG C C, RECKNAGEL F, et al. Adaptation and multiple parameter optimization of the simulation model SALMO as prerequisite for scenario analysis on shallow eutrophic Lake[J]. Ecological Modelling,2014,273(10):109-116.
[23] REYNOLDS C S. The ecology of freshwater phytoplankton[M]. Camridge: Cambridge University Press,1984.
[24] LUDWIG J, MIKHAIL V Z, JOHN P, et al. Significant CO2fixation by small prymnesiophytes in the subtropical and tropical northeast Atlantic Ocean[J]. The ISME Jouranl,2010,4:1180-1192.
[25] 杨威.水中无机碳对浮游植物生长的影响研究[D].天津:南开大学,2007.
[26] GRANELI E, CARLSSON P, LEGRAND C. The role of C, N and P in dissolved and particulate matter as a nutritional source for phytoplankton growth,including toxic species[J]. Aquatic Ecology,1999,33:17-27.
Seasonal Dynamics Characteristics and Physical Factors of Phytoplankton Functional Groups in Sanhuanpao Wetland Reserve//
Ma Chengxue, Yin Zilong, Yu Hongxian
(Northeast Forestry University, Harbin 150040, P. R. China)//Journal of Northeast Forestry University,2016,44(11):45-51.
Phytoplankton; Functional groups; Sanhuanpao Wetland Reserve
1)中央高校基本科研业务费专项资金(2572015CA19、2572014EA07-04)、国家自然科学基金项目(41271106)。
马成学,男,1979年9月生,东北林业大学野生动物资源学院,讲师。E-mail:mch007@163.com。
于洪贤,东北林业大学野生动物资源学院,教授。E-mail:china.yhx@163.com。
2016年5月10日。
X385
责任编辑:任 俐。
