飞播马尾松林林分空间结构对林下植被多样性的影响1)
2016-12-19黎芳潘萍宁金魁赖国桢欧阳勋志徐辉郭丽玲吴自荣易祖滨
黎芳 潘萍 宁金魁 赖国桢 欧阳勋志 徐辉 郭丽玲 吴自荣 易祖滨
(中国林业科学研究院 亚热带林业实验中心,江西省分宜县,336600)江西农业大学)(江西省兴国县林业局)
飞播马尾松林林分空间结构对林下植被多样性的影响1)
黎芳 潘萍 宁金魁 赖国桢 欧阳勋志 徐辉 郭丽玲 吴自荣 易祖滨
(中国林业科学研究院 亚热带林业实验中心,江西省分宜县,336600)江西农业大学)(江西省兴国县林业局)
以飞播马尾松中龄林为研究对象,通过典型样地调查,运用单因素方差及灰色关联分析方法,分析林分空间结构对林下植被多样性的影响。结果表明:(1)大小比数对灌木及草本层多样性及盖度影响均不显著(P>0.05);角尺度、混交度对灌木及草本Shannon-Wiener指数影响显著(P<0.05),且混交度对草本Patrick指数影响达到显著水平(P<0.05);空间密度、开敞度对草本Shannon-Wiener指数影响显著(P<0.05),且开敞度对灌木Patrick指数的影响达显著水平(P<0.05),空间密度对灌木盖度的影响达显著水平(P<0.05);各种空间结构指数对Pielou指数的影响均不显著(P>0.05)。总体上,林下植被多样性表现为林木随机分布的林分高于聚集分布的林分,混交林高于纯林,中等密度林分高于高密度林分,开敞度基本充足的林分高于开敞度严重不足的林分。(2)受林下植被物种多样性影响最大的是空间密度,最小的是大小比数。因此,以调整林分密度为主,综合调整林木分布格局、树种结构及其开敞度,将有利于提高林下植被的多样性。
飞播造林;马尾松林;林下植被;林分空间结构;植被多样性
With the understory vegetation of aerial seedingPinusmassonianaplantations in the middle age, we explored the effect of spatial structure on the understory vegetation diversity by investigating the typical plots by one-way ANOVA method and grey correlation analysis method. The results showed that: (1)Both uniform angle index and mingling degree had significant influence on Shannon-Wiener index of shrub and herb; the influence of mingling degree to Patrick index of herb also reached a significant level, while non-significant influence of neighborhood comparison was found on all diversity indexes of shrub and herb; both the effects of spatial density and opening degree on Shannon-Wiener indexes of herb were significant. The opening degree had a significant impact on the Patrick index of shrub and spatial density had a significant impact on the cover of shrub; non-significant relationship was found between all spatial structure indexes and Pielou index. Overall, the diversity indexes of understory vegetation were the mixed stands>the pure stands, the stands of random distribution>the stands of aggregated distribution, middle density stands>high density stand, and the stands of basically adequate opening degree>the stands of seriously short opening degree. (2)Through the grey correlation analysis between the stand spatial structure indexes and understory vegetation diversity indexes, spatial density had the greatest influence on the species diversity of understory vegetation, and the minimum was the neighborhood comparison. Therefore, after giving priority to adjust the stand density integrated with regulating the spatial pattern of the tree, species structure and opening degree will be conducive to improve the diversity of the understory vegetation.
林下植被是森林生态系统的重要组成部分,在维持系统稳定、防止水土流失、促进养分循环等方面发挥着重要作用。物种多样性是维持整个森林生态系统稳定的关键因子之一[1],林下植被盖度的改变可影响土壤的径流和侵蚀[2]。林下植被的生长主要受到光照、土壤、地形、海拔、林分结构等因素的影响和制约,这些因素影响着林下植被群落的分布及物种多样性。乔木层作为森林生态系统的主体,其林木的空间结构特征将对林分空间异质性产生明显的影响,导致林下植被生长的微环境发生变化,从而影响林下植被的多样性。Basken et al.[3]认为,如果忽略空间细节特征,维持环境和生态条件(如生物多样性)是不可能的。当前对影响林下植被多样性因子的研究,大多数集中在纬度、海拔、坡度、坡向、土壤厚度等环境因子以及树高、直径结构、林龄、森林冠层、林分密度、郁闭度等林分非空间结构因子[4-7],针对林分空间结构对林下植被多样性影响的研究较少。
马尾松(Pinusmassomiana)是我国南方地区飞播造林的主要树种,适应性强、耐干旱与贫瘠[8],是亚热带退化生态系统恢复中的优良先锋树种,飞播造林是快速恢复植被的有效手段[8-10]。赣南曾是水土流失较严重的地区之一,尤其是该区的兴国县,水土流失范围广、强度大,曾被称为“江南沙漠”。20世纪70—90年代,进行了大面积的马尾松飞播造林,目前飞播马尾松林保存面积6.4万hm2,占全县有林地面积的29.5%[11]。然而,整个飞播区不少林分林下植被稀少,“远看绿油油,近看水土流”的现象仍然普遍存在[12],而一个地区的植物多样性是该地区植被恢复和保护的基础[13]。因此,本文以兴国县为研究区,探讨飞播马尾松林空间结构与林下植被特征的关联,挖掘影响林下植被物种多样性的空间结构因子,为合理制定飞播马尾松林林下植被恢复措施以及土壤严重退化区林下植被恢复机理提供科学依据。
1 研究区概况
兴国县位于江西省赣州市的北部(26°03′~26°42′N,115°01′~115°51′E),地貌以低山、丘陵为主,海拔在130~1 200 m。该区属亚热带东南季风气候区,气候温和,雨量充沛,光照充足,四季分明;年平均气温18.9 ℃,无霜期280~300 d,平均日照时间1 861 h,年平均降水量为1 539 mm。母岩主要为花岗岩、第四纪红色粘土等,土壤类型以酸性红壤为主。主要植被类型有针叶林、常绿阔叶林、针阔混交林和竹林等。
2 研究方法
2.1 样地设置与调查
由于受人为不合理的利用方式及松毛虫危害等因素的影响,飞播的马尾松林保留下来的甚少,现有飞播林绝大部分为中龄林(20~30 a),且飞播区为中丘地貌,土壤主要为花岗岩发育的红壤。同时,为控制坡向、坡位及人为干扰对林下植被的影响,在飞播区选择人为干扰程度低、阳坡、下坡位的中龄林中设置典型样地,共设置16个典型样地,样地面积为900 m2(30 m×30 m),并采用相邻网格调查方法,将样地划分为9个10 m×10 m调查单元。样地调查包括其林木位置(坐标)、树种组成、胸径、树高等林分因子。同时,在每个样地的上、中、下分别设置4 m×4 m的灌木样方,并在所选的灌木样方中各设置1个2 m×2 m的草本样方,调查记录植被的种类、数量和盖度等因子。
2.2 分析方法
林分空间结构指数:为了消除处于林分边缘树木系统的影响,本研究设置5 m的缓冲区,根据前人研究经验[14],采用时,构成的空间结构单元来分析林分结构。本文分别选取大小比数、角尺度、混交度、空间密度、开敞度为空间结构指数[15-18]。

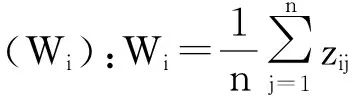
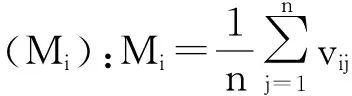
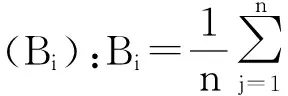

林下植被多样性指数及盖度:在参考相关文献[19-22]的基础上,林下植被的多样性研究从多样性、均匀度、丰富度3个方面分别选用Shannon-Wiener指数、Pielou指数和Patrick指数。
Pielou指数(Js):Js=H/lnS;
Patrick指数(P):P=S;
林下植被盖度(C):C=样地内灌木(或草本)植被垂直投影面积/样地面积。
式中:S为物种总数;Pi是第i种的个体数(ni)占总个体数(N)的比例,即Pi=ni/N;i=1,2,3,…,S。
2.3 数据处理
数据分析主要运用SPSS17.0和DPS9.5统计软件,运用聚类分析将空间密度分类,运用单因素方差分析林下植被不同林分空间结构类型多样性特征,运用灰色关联分析法分析各个物种多样性指标与林分空间结构的关联度及排序。数据处理与图表绘制在MicrosoftExcel2003和Origin7.5中完成。
3 结果与分析
3.1 空间结构类型的划分
为便于统计分析,根据16个典型样地实际情况,将空间结构指数(大小比数、混交度、角尺度、开敞度)计算结果进行分类,分类参照相关文献[17,23-24];而空间密度前人未有分组情况,根据计算结果采用聚类分析分成3种类型。具体分类结果见表1。
3.2 单因素方差分析
由表2可知,5个指数的不同空间结构类型对Pielou指数均无显著影响(P>0.05);不同大小比数类型对灌木层各项多样性指标的影响均未达到显著水平(P>0.05);不同混交度类型对灌木Shannon-Wiener指数的影响达显著性水平(P<0.05),且该指数表现为弱度、中度混交类型时显著高于零度混交类型(P<0.05),呈零度混交<强度混交<弱度混交<中度混交,换言之,适当的混交度有利于灌木多样性的增加;不同角尺度类型对灌木Shannon-Wiener指数的影响也达显著水平(P<0.05),随机分布时的灌木Shannon-Wiener指数显著高于其它两种类型(P<0.05),呈现聚集分布<均匀分布<随机分布,这说明林木的分布格局越趋于随机分布,灌木Shannon-Wiener指数越大;不同开敞度类型对灌木Patrick指数的影响达到显著差异水平(P<0.05),各个多样性指数随着开敞度增加呈现增长的趋势,Patrick指数由开敞度严重不足至基本充足显著升高(P<0.05),表现为严重不足<不足<基本充足,即开敞度越大,越有利于灌木植物的生长;不同空间密度类型对灌木层多样性的影响除Shannon-Wiener指数外,盖度也达到显著水平(P<0.05),均为中等密度的指标显著高于较高密度的指标(P<0.05),表现为较高密度<低密度<中等密度,即空间密度的差异会影响灌木植物的多样性以及盖度。

表2 不同空间结构类型灌木层多样性及其盖度特征

空间结构类型开敞度Shannon-Wiener指数Pielou指数Patrick指数盖度空间密度Shannon-Wiener指数Pielou指数Patrick指数盖度I1.07a0.85a4.60b13.4a 0.84b0.86a5.80a9.00bII1.24a0.90a7.14ab19.71a1.59a0.99a7.75a32.50aIII1.54a0.93a8.25a24.50a1.38ab0.88a6.57a22.00abIV
注:同列不同小写字母表示差异显著(P<0.05)。
由表3可知,草本Pielou指数也对空间结构类型的响应不显著(P>0.05);不同大小比数类型对草本层的多样性影响未达到显著水平(P>0.05);不同混交度类型对草本Shannon-Wiener指数和Patrick指数影响显著(P<0.05),Shannon-Wiener指数和Patrick指数均表现为中度混交,显著高于零度混交(P<0.05),呈零度混交<强度混交<弱度混交<中度混交,这反映出草本层多样性指数在不同混交度类型的差异较灌木层表现的更为明显;不同角尺度类型对草本Shannon-Wiener指数和盖度的影响均达到显著水平(P<0.05),Shannon-Wiener指数与盖度在随机分布类型时显著高于其它类型(P<0.05),呈聚集分布<均匀分布<随机分布;不同开敞度类型和空间密度类型对草本Shannon-Wiener指数的影响显著(P<0.05),开敞度表现为类型III显著高于类型I(P<0.05),随着空间密度减小,各项多样性指数呈较高密度<低密度<中等密度,这说明开敞度与空间密度均对林下草本的生长条件有一定的影响,从而导致多样性的变化。
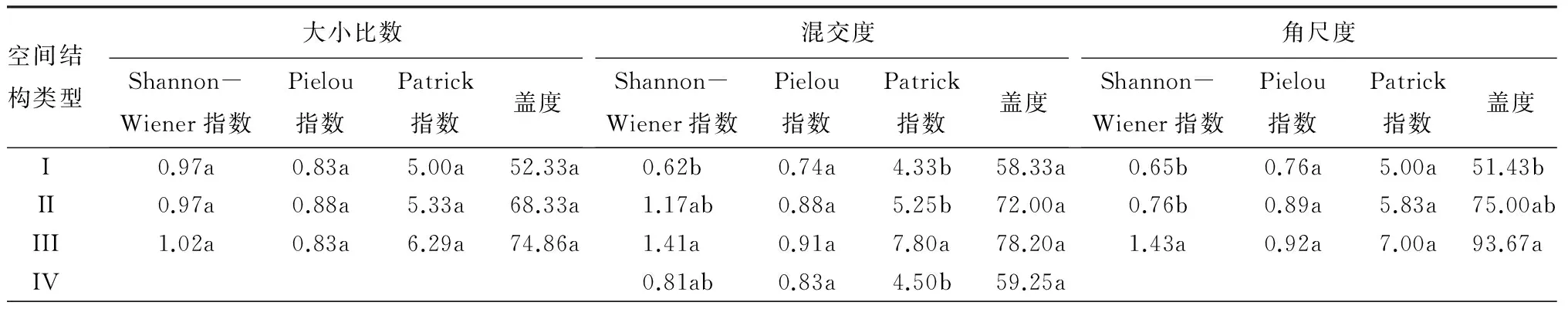
表3 不同空间结构类型草本层多样性及其盖度特征

空间结构类型开敞度Shannon-Wiener指数Pielou指数Patrick指数盖度空间密度Shannon-Wiener指数Pielou指数Patrick指数盖度I0.71b 0.84a5.00a56.86a0.71b0.79a4.57a51.71aII1.03ab0.78a5.80a69.60a1.44a0.92a7.00a88.25aIII1.42a0.96a6.75a86.25a0.93b0.85a6.20a75.20aIV
注:同列不同小写字母表示差异显著(P<0.05)。
3.3 灰色关联分析
为进一步探讨林分空间结构对林下植被多样性的影响,将16个样地林下灌木层和草本层多样性指数和空间结构指数作为一个灰色系统,分别以灌木层、草本层植被多样性指数(Shannon-Wiener指数、Pielou指数、Patrick指数)和盖度为参考序列,以空间结构指数(角尺度、空间密度、混角度、开敞度、大小比数)为子序列,运用DPS 9.5进行灰色关联分析,结果见表4。由表4可知,影响灌木层Shannon-Wiener指数关联序由大到小的顺序为:空间密度、角尺度、混交度、开敞度、大小比数;影响灌木层Pielou指数关联序由大到小的顺序为:角尺度、空间密度、混交度、开敞度、大小比数;影响灌木层Patrick指数关联序由大到小的顺序为:空间密度、开敞度、角尺度、混交度、大小比数;影响灌木层盖度关联序由大到小的顺序为:空间密度、开敞度、角尺度、混交度、大小比数。可以看出,灌木层物种多样性与空间结构指数的灰色关联度均较高,对于不同多样性指数,同一空间结构指数与它们的关联序却不一样,即不存在某一空间结构指数对物种多样性指数起着决定性的作用,说明灌木层特征不只是由于一个空间结构指数的影响,它受多重指数共同影响。总体而言,对灌木多样性影响较大的空间结构指数主要为空间密度、角尺度,其次是开敞度、混交度,大小比数对灌木多样性的指数的影响最小。
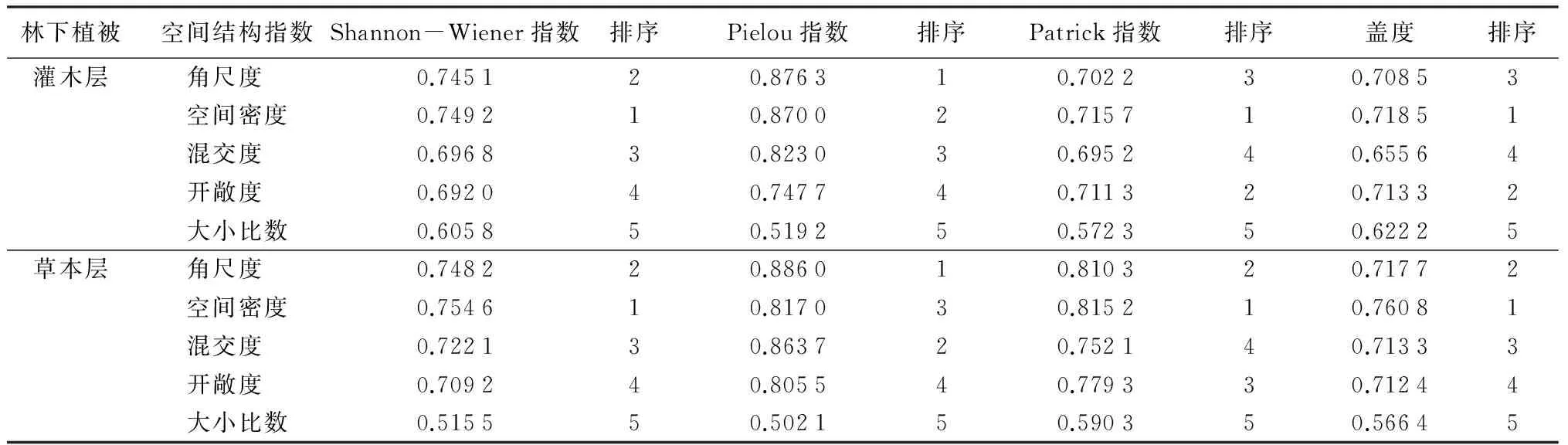
表4 林下植被物种多样性与空间结构关联度及其排序
影响灌木层Shannon-Wiener指数关联序由大到小的顺序为:空间密度、角尺度、混交度、开敞度、大小比数;影响灌木层Pielou指数关联序由大到小的顺序为:角尺度、混交度、空间密度、开敞度、大小比数;影响灌木层Patrick指数关联序由大到小的顺序为:空间密度、角尺度、开敞度、混交度、大小比数;影响灌木层盖度关联序由大到小的顺序为:空间密度、角尺度、混交度、开敞度、大小比数。可以看出,空间密度、角尺度、混交度、开敞度对草本层多样性影响较大,与影响灌木层的主要因子较为一致,大小比数影响最小。
总体而言,对林下植被物种多样性影响最大的是空间密度,角尺度、混交度、开敞度紧随其后,最小为大小比数,但两层次各个空间结构指数与多样性指数的关联序也存在一些差异,这说明林下灌木、草本受林内光照、湿度等微环境的影响还是有些不同,其影响机制较为复杂。
4 讨论与结论
研究发现,对林下植被物种多样性及盖度影响最大的空间结构指数是空间密度。不同空间密度对林下植被Shannon-Wiener指数、灌木盖度影响均达显著水平(P<0.05)。乔木层影响草本层的机理主要是乔木密度通过影响光照、降水以及枯落物量,从而影响到草本层[25],乔木层影响灌木的机理可能一致。密度较高的林分林内光照不足、林木对水分的竞争增加,抑制了林下植被的生长发育,低密度林分可能由于枯落物相对较少,影响土壤养分,从而不利于林下植被的生长,即飞播马尾松林林分密度过高或过低均不利于林下植被的生长[26]。
对林下植被多样性及盖度影响较大的空间结构指数是角尺度、混交度、开敞度。不同角尺度类型对林下植被Shannon-Wiener指数、草本盖度影响均达显著水平(P<0.05),这可能是因为上层林木由于其分布格局的不一样,影响枯落物分布以及光斑分布,导致林下植被多样性特征出现差异。不同混交度类型对林下植被Shannon-Wiener指数、草本Patrick指数影响均达显著水平(P<0.05),从关联序可知,混交度对草本层的作用较灌木层更为明显,究其原因,可能是在光照条件不及灌木层的草本层,营养主要来源于土壤,在马尾松纯林增加阔叶树比例能显著增加林分枯落物的现存量[27],影响土壤的营养;混交林较纯林而言,阔叶树的生态位与马尾松的生态位在一定程度上相互抑制,为其林下植被的生长释放了一定的营养空间,这可能就是导致混交度与林下植被物种多样性关联密切的重要原因。不同开敞度类型对灌木Patrick指数、草本Shannon-Wiener指数影响均达显著水平(P<0.05),光照被认为是限制森林植被的盖度和(或)物种丰富度的主要因素[28],作为林下光照条件的测度指标,开敞度不同导致林分的透光度有差异,从而影响林下植被的生长;而其对草本盖度或丰富度影响不显著可能说明草本层由于乔木层和灌木层的遮蔽作用,光照条件不及灌木层,因而对光照变化的反应不及灌木层敏感。
对林下植被多样性及盖度影响最小的空间结构指数是大小比数,不同大小比数类型对林下植被多样性影响均不显著(P>0.05),导致这一结果的原因可能是大小比数主要反映林木间的竞争,对林下植被多样性的影响较为微弱。此外,5种指数的不同空间结构类型对林下植被Pielou指数影响均不显著(P>0.05),这可能是因为部分植物在环境内的无序分布主要受物种种类影响造成的[29]。
综上所述,林下植被多样性及盖度变化表现为:林木随机分布的林分高于聚集分布的林分、混交林高于纯林、密度中等的林分高于高密度林分、开敞度基本充足的林分高于开敞度严重不足的林分,其混交等方面的结论与燕亚飞等[30]、Barbier et al.[31]的研究结果相似。因此,在实际应用中可以调整林分密度为主,综合调整林木分布格局、树种结构及其开敞度,这将对提高林下植被多样性方面起到重要作用。
运用灰色关联度分析得出飞播马尾松林林分空间结构与林下植被多样性及盖度关系密切,影响林下植被的最主要的因子为空间密度。虽然用回归、相关等统计分析方法也可以做出一定程度的回答,但对数据量和数据的分布特征要求高,很难分析各因子的主次关系,而灰色关联分析作为一种系统分析技术,对数据要求低,适用于部分信息已知、部分信息未知的不确定性系统[29],对上述不足有所克服和弥补。由于本研究样本数量有限,以后可扩大样本数量,建立林下植被多样性与林分空间结构因子之间的关系模型,为进一步探明林分空间结构影响林下植被多样性的机理提供科学的依据。
[1] KERNS B K, OHMANN J L. Evaluation and prediction of shurb cover in coastal oregon forests(USA)[J]. Ecological Indicators,2004,4(2):83-98.
[2] NEARING M A, JWTTEN V, BAFFAUT C, et al. Modeling response of soil erosion and runoff to changes in precipitation and cover[J]. Catena,2005,61(2/3):131-154.
[3] BASKENT E Z, KELES S. Spatial forest planning a review[J]. Ecological Modelling,2005,188(2/3/4):145-173.
[4] 李际平,赵春燕,袁晓红,等.杉阔间物种多样性影响因子灰色关联分析[J].中南林业科技大学学报,2011,31(10):10-14.
[5] 卢训令,丁圣彦,游莉.伏牛山自然保护区森林冠层结构对林下植被特征的影响[J].生态学报,2013,33(15):4715-4723.
[6] TILMAN D. Causes, Consequences and ehics of biodiversity[J]. Nature,2000,405:208-211.
[7] 康冰,刘世荣,蔡道雄,等.马尾松人工林林分密度对林下植被及土壤性质的影响[J].应用生态学报,2009,20(10):2323-2331.
[8] 林惠章,郭孝玉,欧阳勋志,等.飞播马尾松纯林补植木荷后土壤酶活性与土壤养分变化[J].东北林业大学学报,2015,43(7):136-139.
[9] 万杨,付晓莉,杨风亭,等.南方稀疏马尾松老头林形成的种源供给和种子萌发环境限制分析[J].江西农业大学学报,2014,36(4):782-789.
[10] XIAO X, WEI XH, LIU YQ, et al. Aerial seeding: an effective forest restoration method in highly degraded forest landscapes of sub-tropic regions[J]. Forests,2015,6(6):1748-1762.
[11] 丁松,应学亮,吕丹,等.赣南飞播马尾松林林下植被盖度对土壤质量的影响[J].水土保持研究,2014,21(3):31-36.
[12] 赵芳,欧阳勋志.飞播马尾松林林下植被盖度与环境因子的关系[J].应用生态学报,2015,26(4):1071-1076.
[13] 曲红,王百田,王棣,等.黄土区不同配置人工林物种多样性研究[J].生态环境学报,2010,19(4):843-848.
[14] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27.
[15] 惠刚盈,KLAUSVON G, MATTHIAS A.一个新的林分空间结构参数:大小比数[J].林业科学研究,1999,12(1):4-9.
[16] 惠刚盈,胡艳波.角尺度在林分空间结构调整中的应用[J].林业资源管理,2006(2):31-35.
[17] 邓送求,闫家锋,关庆伟.宝华山紫楠风景林林分空间结构分析[J].东北林业大学学报,2010,38(4):29-32.
[18] 李建军,刘帅,张会儒,等.洞庭湖森林生态系统空间结构均质性评价[J].生态学报,2013,33(12):3732-3741.
[19] 于景金,陈丽华,谷建才,等.基于灰色关联分析的环境因子与物种多样性关系[J].林业资源管理,2010(1):68-72.
[20] 马克平,刘玉明.生物群落多样性的测度方法-Iα多样性的测度方法:下[J].生物多样性,1994,2(4):231-239.
[21] 马克平.生物群落多样性的测度方法-Iα多样性的测度方法:上[J].生物多样性,1994,2(3):162-168.
[22] 刘灿然,马克平,于顺利,等.北京东灵山地区植物群落多样性的研究IV:样本大小对多样性测度的影响[J].生态学报,1997,17(6):584-592.
[23] 马洪婧,袁发银,刘中亮,等.北亚热带栎树混交林空间结构特征分析[J].西北林学院学报,2013,28(1):151-156.
[24] 曹小玉,李际平,封尧,等.不同龄组杉木生态公益林空间结构[J].浙江农林大学学报,2015,32(1):84-91.
[25] 夏富才,潘春芳,赵秀海,等.长白山原始阔叶红松林林下草本植物多样性格局及其影响因素[J].西北植物学报,2012,32(2):370-376.
[26] 肖欣,郭孝玉,欧阳勋志,等.不同密度飞播马尾松林林下植被特征及土壤质量评价[J].东北林业大学学报,2015,43(9):62-66.
[27] 殷沙,赵芳,欧阳勋志.马尾松木荷不同比例混交林枯落物和土壤持水性能比较分析[J].江西农业大学学报,2015,37(3):454-460.
[28] JENNINGS S B, BROWN N D, SHEIL D. Assessing forest canopies and understorey illumination:canopy closure,canopy cover and other measures[J]. Forestry,1999,72(1):59-73.
[29] 李倩,梁宗锁,董娟娥,等.丹参品质与主导气候因子的灰色关联度分析[J].生态学报,2010,30(10):2569-2575.
[30] 雷相东,唐守正,李冬兰,等.影响天然林下层植物物种多样性的林分因子的研究[J].生态学杂志,2003,22(3):18-22.
[31] BARBIER S, GOSSELIN F, BALANDIER P. Influence of tree species on understory vegetation diversity and mechanisms involved-A critical review for temperate and boreal forests[J]. Forest Ecology and Management,2008,254(1):1-15.
Effects of Stand Spatial Structure on Understory Vegetation Diversity of Aerial SeedingPinusmassonianaPlantations//
Li FangPan Ping, Ning Jinkui, Lai Guozhen, Ouyang Xunzhi, Xu Hui, Guo Liling, Wu ZirongYi Zubin
(Experimental Center for Subtropical Forestry, Fenyi 336600, P. R. China); (Jiangxi Agricultural University); (Forestry Bureau of Xingguo County)//Journal of Northeast Forestry University,2016,44(11):31-35,40.
Aerial seeding;Pinusmassonianaplantations; Understory vegetation; Stand spatial structure; Diversity
1)国家自然科学基金项目(31360181)。
黎芳,女,1992年5月生,江西农业大学林学院,现工作于中国林业科学研究院亚热带林业实验中心,助理工程师。E-mail:lffnms@163.com。
欧阳勋志,江西农业大学林学院,教授。E-mail:oyxz_2003@hotmail.com。
2016年4月21日。
S718.5
责任编辑:王广建。
