汕头3种人工红树林湿地大型底栖动物群落的比较
2016-12-13唐以杰陈思敏方展强罗丽芬陈晓芬黄慧娟
唐以杰, 陈思敏, 方展强, 罗丽芬, 陈晓芬, 黄慧娟, 陆 棣
汕头3种人工红树林湿地大型底栖动物群落的比较
唐以杰1, 陈思敏1, 方展强2, 罗丽芬1, 陈晓芬1, 黄慧娟1, 陆 棣1
(1. 广东第二师范学院生物与食品工程学院, 广东广州 510303; 2. 华南师范大学生命科学学院, 广东省高等学校生态与环境科学重点实验室, 广东广州 510631)
对广东省汕头市莲阳河口区恢复背景相同的拉关木()、无瓣海桑()、海桑()3种人工林和光滩湿地中大型底栖动物群落进行比较研究。结果显示, 4种生境中大型底栖动物优势种(>0.02)存在差异; 相似性分析检验(One-Way ANOSIM)表明4种生境间大型底栖动物群落结构差异显著。等级聚类和非参数多变量标序也显示4种生境间的大型底栖动物群落结构差异显著。3种人工林中, 无瓣海桑、海桑人工林中大型底栖动物群落结构较为相似; 3种人工林中大型底栖动物物种多样性和生物量从大到小的排列次序为: 拉关木>海桑>无瓣海桑。BIOENV分析表明大型底栖动物分布与林地土壤pH、有机质、砂粒和粉粒含量、红树植物平均株高、平均胸径、平均基径和冠幅(南北×东西)这些理化因子显著相关, 这进一步证实了红树植物种类对林地大型底栖动物多样性和分布的影响。
拉关木(); 无瓣海桑(); 海桑(); 大型底栖动物
随着对红树林生态功能的进一步认识, 恢复红树林以保护海岸湿地生物多样性的工作受到广泛重视。许多地方开展了红树林的人工种植, 以改善当地的生态环境。这些用于种植的红树植物除了秋茄、桐花树、木榄、海桑和红海榄等乡土树种外, 还从国外引进了一些生长迅速、结实率高、定居容易、适应性广的树种, 比如来自孟加拉国的无瓣海桑 ()[1]以及引种墨西哥的拉关木 ()[2], 这些树种的引进, 扩大了红树种质资源, 改善了滩涂环境, 但是, 与乡土红树植物相比, 这些树种的种植, 对林下湿地底栖动物的影响如何, 研究甚少, 尤其是拉关木, 未见报道; 而且, 对于同为海桑属()的无瓣海桑与海桑(), 对林下湿地底栖动物的影响有没差别也未见报道。大型底栖动物作为红树林生态系统中物质循环、能量流动积极的消费者和转移者, 对红树林生态系统的生态功能有重要意义。大型底栖动物的多样性和丰度对天然和人工恢复红树林的生境变化均具有潜在的生物、生态指示作用[3]。鉴于此, 本研究通过比较恢复背景和林龄相同的拉关木、无瓣海桑和海桑人工林湿地中大型底栖动物群落的差异, 以及影响大型底栖动物群落结构和物种多样性的主要环境因子, 以期为无瓣海桑、拉关木的科学引种和滩涂红树林造林提供决策依据。
1 材料与方法
1.1 区域概况
调查区位于广东省东南部, 地理位置处于23°02′33″~23°38′58″N, 116°14′40″~117°19′36″E, 北回归线从市区穿过, 属南亚热带海洋性季风气候, 温暖湿润, 阳光充足, 雨水充沛, 无霜期长, 冬无严寒, 夏无酷暑。年日照2 000~2 500 h, 日照最短为每年3月。年降雨量1 300~1 800 mm, 多集中在4~9月。年平均气温21~22℃; 最低气温在0℃以上, 冬季偶有霜冻; 最高气温36~40℃, 多出现在7月中旬至8月初受太平洋副热带高压控制期间[4]。
1.2 采样方法及样品处理
分别于2013年4月、7月和10月, 2014年1月, 在汕头市澄海区莲阳河红树林区, 选择恢复背景相同的6年林龄拉关木、无瓣海桑、海桑人工林和周围20 m内无植被的光滩; 每个研究样地从沿岸端向近海端选择4个较为典型的地方布设站点(分别标记为A1~A4, B1~B4, C1~C4, D1~D4)。大型底栖动物的调查, 采用25 cm × 25 cm的定量样框, 每个站点取4个样框, 先捡出框内底表动物, 再挖至30 cm深的沉积物, 用0.5 mm孔径筛子分选出底内动物, 所获样品用5%的甲醛溶液现场固定, 于实验室内分析鉴定。在进行大型底栖动物调查的同时, 每个站点布设一个10 m × 5 m的样方, 对样方内所有植物进行全面调查, 测定各样方内红树植物的平均树高、胸径、基茎和树冠幅, 同时, 每个样方按对角线随机选5个样点, 挖取0~30 cm层的底泥, 均匀混合, 取1 kg左右的混合沉积物作为测试样品。沉积物粒径组成、盐分和有机质含量分别用比重计法、重量法和重铬酸钾法测定, pH值用电位法, 每个样品3次重复, 各生境理化因子水平取全年的平均值。
1.3 大型底栖动物数据分析
1.3.1 物种优势度指数(
=P×f(1)
式中,P为研究区种的个体数占总个体数的比例,f为种在研究区内各站位出现的频率, 当>0.02时, 该种为优势种。物种数和物种优势度指数计算的数据取各生境4个站点的全年平均值。
1.3.2 多样性测度
采用Margalef物种丰富度指数()、Shannon- Wiener多样性指数(′) 以及Pielou均匀度指数(′)。
Margalef物种丰富度指数:(2)
Shannon-Wiener指数:(3)
Pielou均匀度指数:;(4)
1.3.3 物种数、栖息密度和生物量差异显著性分析
对不同人工林和光滩湿地中大型底栖动物群落的平均栖息密度和平均生物量等用SPSS19.0软件包进行单因素方差分析(One-Way ANOVA)。
1.3.4 动物群落结构的分析
大型底栖动物群落结构的多变量分析采用非参数多变量群落结构分析方法, 为了减少机会种对群落结构的干扰, 删除在总体中相对丰度<1%的种类, 但保留其中任一站点相对丰度>3%的种类。首先以相似性系数为基础构建相似性矩阵, 进一步进行类平均法站点等级CLUSTER聚类和非参数多变量排序(MDS), 然后利用PRIMER软件包中的图形分析和多变量分析等方法对群落结构进行研究[5]。
2 结果与分析
2.1 各生境环境因子的理化性状
对各生境的统计分析(表1)表明, 3种人工林红树植物平均树高、平均胸径、平均基径和冠幅(南北×东西)从大到小的排列次序为: 无瓣海桑>海桑>拉关木, 林地土壤盐分、有机质和粉粒含量从大到小的排列次序也为: 无瓣海桑>海桑>拉关木, 而林地土壤pH、砂粒含量从大到小的排列次序刚好相反: 拉关木>海桑>无瓣海桑。3种林地土壤pH、砂粒含量均低于光滩, 而土壤盐分、有机质、黏粒和粉粒含量均高于光滩。
表1 各生境理化因子水平(平均值±标准差)
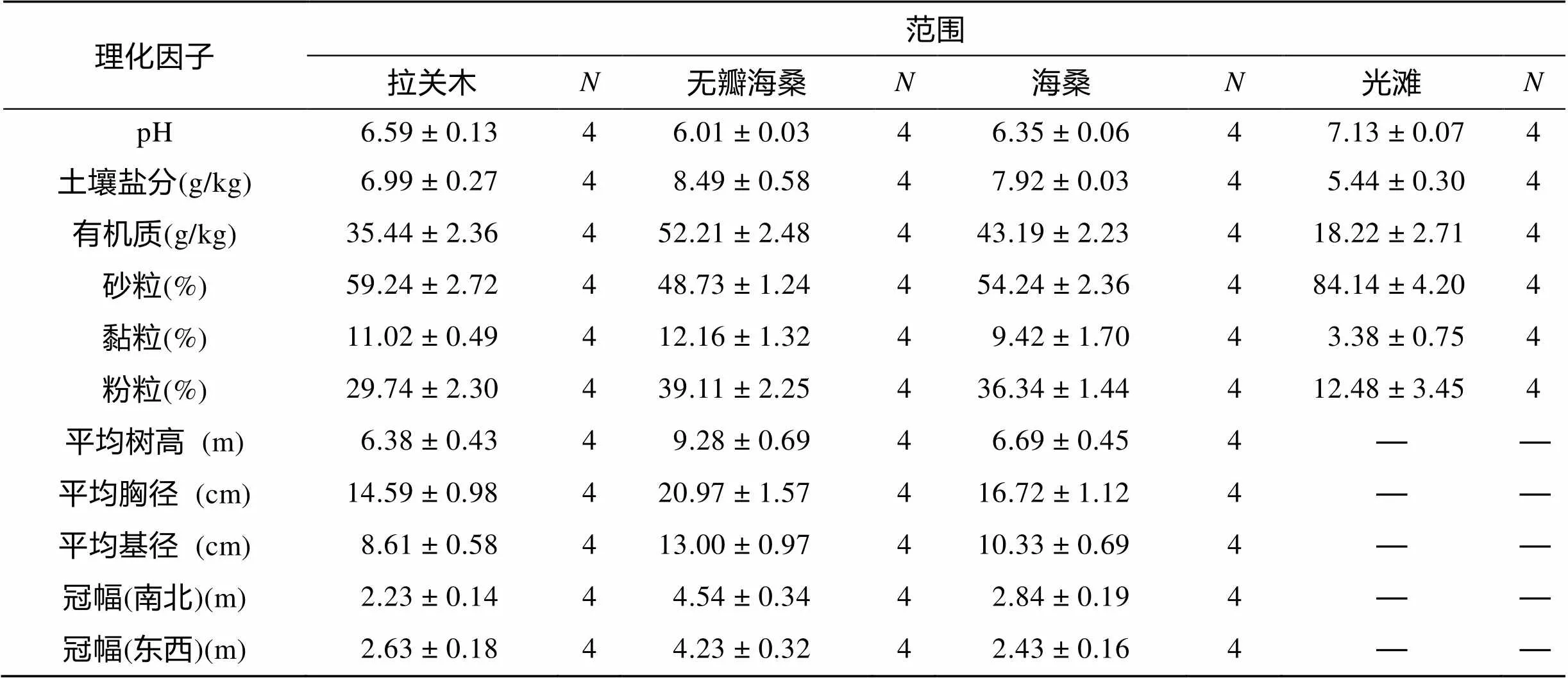
Tab.1 Levels of physicochemical factorsin different habitats (mean ± S.D.)
2.2 大型底栖动物种类组成和优势种
调查发现, 除光滩外, 3种人工林湿地均未发现底内型生活的多毛类, 而且, 3种人工林湿地底内型的双壳类物种数也明显低于光滩。3种人工林湿地均以腹足类和甲壳类为主, 总的物种数从大到小的排列次序为: 拉关木>海桑>无瓣海桑(表2)。

表2 各生境大型底栖动物物种分布
不同生境大型底栖动物的优势种存在差异(表3)。4种生境优势种物种数从大到小排列次序为: 光滩(7)>拉关木(6)=海桑(6)>无瓣海桑(5), 同一物种在不同生境中优势度指数有差异, 比如琵琶拟沼螺在不同生境的优势度指数从大到小排列次序为: 无瓣海桑>拉关木>海桑>光滩。3种人工林中物种优势度指数最高和最低值相差分别为: 无瓣海桑(0.576)> 拉关木(0.41) >海桑(0.26)。
2.3 大型底栖动物密度、生物量
不同季节不同生境大型底栖动物群落密度和生物量的季节变化见表4, 以季节和生境为因素的密度和生物量无重复双因素方差分析显示: 大型底栖动物密度在4个生境间差异显著(= 17.94,<0.01), 季节间差异不显著(= 3.49,>0.05); 生物量在4个生境间差异显著(= 38.34,<0.01), 季节间差异不显著(= 2.15,>0.05)。3种人工林湿地底栖动物群落密度和生物量排列次序均为: 拉关木>海桑>无瓣海桑。

表3 不同生境大型底栖动物优势种
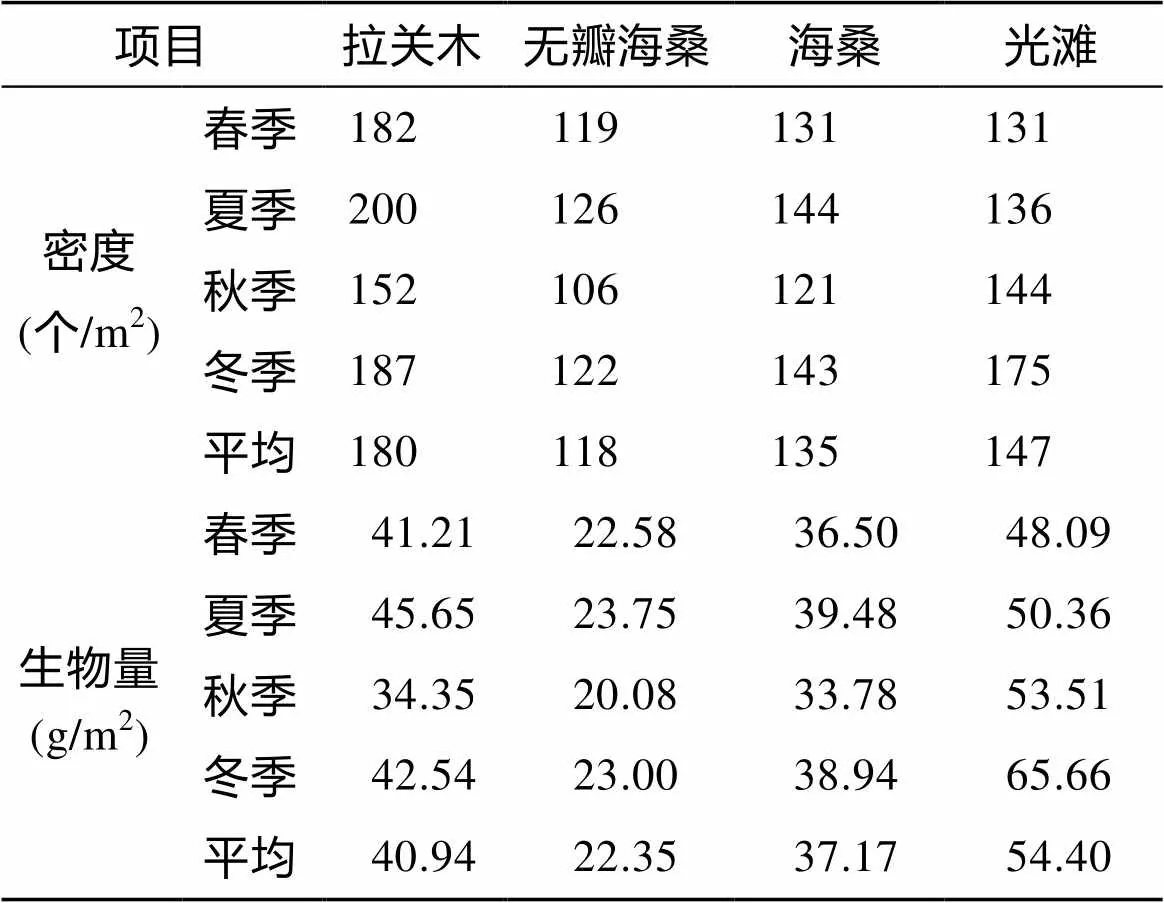
表4 各生境大型底栖动物群落不同季节的密度和生物量
2.4 4种生境大型底栖动物物种多样性
不同季节不同生境大型底栖动物群落物种多样性指数的变化见表5, 以季节和生境为因素的多样性指数无重复双因素方差分析显示: 大型底栖动物Shannon-Wiener多样性指数在4个生境间差异显著(= 816.01,<0.01), 季节间差异不显著(= 0.16,> 0.05); Pielou均匀度指数生境间差异显著(= 406.06,<0.01), 季节间差异不显著(= 3.00,>0.05); Margalef丰富度指数生境间差异显著(= 82.59,<0.01), 季节间差异不显著(= 0.30,>0.05)。4种生境中, 全年平均的Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数均呈现相同的变化趋势, 即光滩>拉关木>海桑>无瓣海桑。
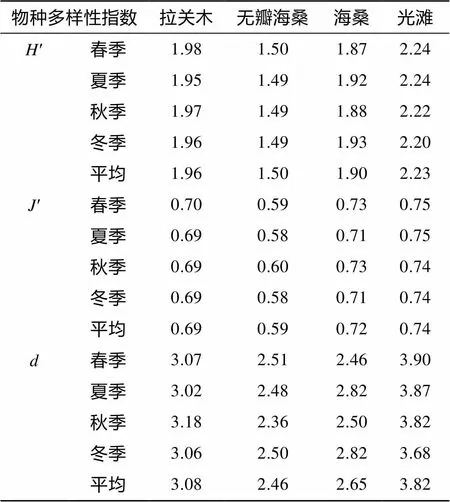
表5 各生境大型底栖动物群落不同季节的物种多样性指数
2.5 4种生境大型底栖动物群落结构
聚类分析结果如图1所示: 把16个站点分成了4个不同的组, 可见, 4种生境的大型底栖动物群落可以区分开来, 3种人工林湿地大型底栖动物群落之间相似性相对较高, 而光滩与这3种人工林的大型底栖动物群落结构相似性相对较低。3种人工林中, 无瓣海桑、海桑人工林中大型底栖动物群落结构较为相似。
排序分析结果如图2所示: stress=0.01<0.1说明这个结果支持聚类分析结果, 也将相同生境中的大型底栖动物群落分别划分到一个组内。图上代表无瓣海桑人工林的4个站点(B1~B4)与代表海桑人工林的4个站点(C1~C4)间的距离最小, 说明2种人工林湿地的大型底栖动物群落结构最相似, 且与其他生境动物群落结构差异较大。
用One-Way ANOSIM检验4种生境间的差异性, 结果显示, 4种生境大型底栖动物群落结构间的差异极显著(Global= 1.00,= 0.001)。而且每2种生境间动物群落结构的差异显著(Global= 1.00,= 0.029)。
2.6 大型底栖动物同环境各理化因子的关系
利用BIOENV分析, 找出与各站点大型底栖动物丰度最为匹配的环境理化因子, 结果发现pH、有机质、砂粒、粉粒、平均株高、平均胸径、平均基径和冠幅(南北×东西)这些理化因子的组合与底栖生物丰度均达到显著的相关水平(=0.746)。RELATE检验结果均显著(= 0.849,=0.001)。这表明上述的理化因子是影响大型底栖动物群落结构变化的重要因子。
3 讨论
红树林大型底栖动物群落受到生境条件的影响, 例如潮汐条件、有机质和沉积物物理性质等[6-9]。而红树植被能够改变红树林的生境条件, 例如改变林内光照、减缓水分蒸发、提供凋落物和改变沉积物性质等[10-12], 因此红树植被在改变林内大型底栖动物的多样性和分布有潜在重要作用。已有研究证明, 红树林种植种类的不同可能导致底栖动物亚系统生态修复过程和结果的差异[13-15]。本研究的3种红树人工林, 虽然恢复背景和林龄相同, 可等级聚类和非参数多变量标序结果均显示3种人工林间大型底栖动物群落结构差异显著。相似性分析检验(One- Way ANOSIM)也表明3种人工林间大型底栖动物群落结构差异显著。这就进一步证实了在小范围区域, 林内动物的分布更多地与红树林植被特性有关[16]。
首先, 红树林植被会改变林内土壤的性质[17-20], 而这些底质因素是影响大型底栖动物组成和分布的重要因子。国内外极少见有关红树林生态恢复对土壤理化性质动态影响的报道[21], 但由于红树植物的介入, 必然导致这些方面的变化。而土壤理化性质如有机质含量和氧化还原条件等又是影响底栖动物分布和栖息密度的重要因素[22]。唐以杰等[13]发现无瓣海桑刚开始引种的前3年, 其林下大型底栖动物生物多样性恢复快于乡土红树植物人工林, 这可能与无瓣海桑人工林快速的植被恢复, 迅速改变荫蔽状况及林内光强有关, 但随后, 其林下大型底栖动物生物多样性反而减少, 这可能与无瓣海桑快速改变生境中的土壤理化性质有关。黄建荣等[23]对珠海淇澳岛红树林不同生境和不同植被群落的蟹类进行了调查, 认为无瓣海桑群落土壤速效磷与蟹类生物量呈极显著正相关, 与蟹类密度呈显著正相关; 全磷与蟹类密度呈显著负相关。本研究中, 3种红树植物的生长速度为: 无瓣海桑>海桑>拉关木(表1), 红树植物生长速度快的人工林, 林地土壤成红树林酸性硫酸盐土的速度也快[24]。因此, 3种红树人工林土壤的pH为: 无瓣海桑<海桑<拉关木(表1), 而酸性土壤明显不利于林下底内型底栖动物的生长, 因此, 三种人工林均未采集到多毛类, 双壳类的物种类也很少(表2); 而且, 随着土壤pH的下降, 林地大型底栖动物生物量和物种多样性明显下降(表4和表5)。
其次, 人工林内较高的郁闭度提高了林地的荫蔽条件, 减轻了高温和水分蒸发对林下动物的胁迫, 同时也给动物提供了一个躲藏和逃避敌害的场所, 因此有利于湿地底栖动物的生长。Nobbs[25]对澳大利亚天然红树林的研究表明, 红树林植被的存在与否对蟹类的种类与分布有很大影响, 主要是由于荫蔽条件的差异。Vannini等[26]的研究也表明, 天然红树林内光照水平对相手蟹的分布有重要影响。泰国白海榄雌()红树林主要大型底栖动物拟沼螺科种类()的栖息密度与红树植物幼苗密度呈极显著正相关关系[27]。虽然, 红树植被的恢复对林下底栖动物有上述益处, 但红树植被郁闭度同时也影响林下一些海藻类的生长。刘玉等[28]和徐姗楠等[29]通过稳定同位素、脂肪酸标志等自然示踪技术, 认为红树林区的鱼虾等捕捞产量较高, 还因红树林提供了凋落物以外的其他食物来源(如林区内浮游植物、附生藻类和底栖微藻)。本研究还显示, 各站点大型底栖动物丰度与红树植物树冠幅(南北×东西)相关显著, 而且树冠幅越大, 动物多样性越低, 这可能是由于人工林的郁闭度过高, 不利于林下浮游植物和海藻类的光合作用, 对于以浮游植物和海藻类为食的动物造成不利影响。
最后, 在红树林生态系统中, 有些无脊椎动物对红树植物叶片的取食具有很强的选择性, 其原因是不同红树植物不同状态的叶片在营养水平(如C/N比)和单宁含量上有差异[3, 16, 30-32]。因此, 外来树种的引进可能会造成湿地动物食物源的改变[33], 从而影响群落结构和物种多样性。比如, 相手蟹属蟹类的分布与红树林土壤类型没有密切的相关性[34], 而与红树林植被的类型则有明显的关系[31]。在香港米埔红树林的调查表明, 两个相距很近的秋茄林和白骨壤()林内相手蟹的种类和数量有很大差异[34]。这可能是由于不同的蟹对不同红树植物凋落腐解叶在啃食上有不同的“选择性”。李旭林等[35]和晏婷等[36]证实了一些动物对无瓣海桑和乡土红树植物凋落叶片取食上却存在显著差异, 说明大面积引种外来物种无瓣海桑将可能对华南沿海红树林生态系统的食物链结构产生潜在影响。因此, 3种人工林间大型底栖动物群落结构差异显著, 还可能由于3种人工林提供给动物的凋落物种类不同, 导致3种人工林间大型底栖动物群落结构差异显著。相比较而言, 3种人工林湿地中, 无瓣海桑与海桑林中大型底栖动物群落结构较为相似, 这可能由于无瓣海桑与海桑同属海桑属, 两种红树植物在各种生理生化特性上相对接近, 对大型底栖动物影响也可能较为接近, 但这需要进一步研究。可见, 通过野外和室内模拟试验, 研究红树林湿地大型底栖动物主要种类对不同红树植物叶片的选择性啃食作用, 在红树林恢复措施的重要方面即红树植物的种类选择上, 对预测红树林植被恢复地底栖动物多样性动态具有指导意义[3]。
参考文献:
[1] 廖宝文, 李玫, 郑松发, 等. 外来种无瓣海桑种内、种间竞争关系研究. 林业科学研究[J], 2003, 16(4): 418-422. Liao Baowen, Li Mei, Zheng Songfa, et al. Study on intraspecific and interspecific competition in exotic species[J]Forest Research, 2003, 16(4): 418-422.
[2] 钟才荣, 李诗川, 杨宇晨, 等. 红树植物拉关木的引种效果调查研究[J]. 福建林业科技, 2011, 38(3): 96-99. Zhong Cairong, Li Shichuan, Yang Yuchen, et al. Analysis of the introduction effect of a mangrove species[J]Journal of Fujian Forestry Science and technology, 2011, 38(3): 96-99.
[3] 叶勇, 翁劲, 卢昌义, 等. 红树林生物多样性恢复[J]. 生态学报, 2006, 26(4): 1243-1250. Ye Yong, Weng Jin, Lu Chang yi, et alMangrove biodiversity restoration[J]. Acta Ecologica Sinica, 2006, 26(4): 1243-1250.
[4] 陈远合, 肖泽鑫, 彭剑华, 等. 粤东海桑、无瓣海桑、拉关木冻害调查报告[J]. 防护林科技, 2010, (4): 15-17. Chen Yuanhe, Xiao Zexin, Peng Jianhua, et al. Investigation on freeze injury of,&in Eastern of Guangdong Province[J]. Protection Forest Science and Technology, 2010, (4): 15-17.
[5] 周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J]. 青岛海洋大学学报, 2003, 33(1): 58-64. Zhou Hong, Zhang Zhinan. Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology[J]. Journal of Ocean University of Qingdao, 2003, 33(1): 58-64.
[6] 余日清, 陈桂珠, 黄玉山, 等. 深圳福田红树林区底栖大型动物群落的空间分带及灌污的可能影响[J]. 生态学报, 1996, 16(3): 283-288. Yu Riqing, Chen Guizhu, Wong Yukshan, et al. Sapatial zonation of benthic macrofauna and possible effects of sewage discharge on it in Futian mangrove swamp, Shenzhen[J]. Acta Ecologica Sinica, 1996, 16(3): 283 288.
[7] 蔡立哲, 谭凤仪, 黄玉山. 香港东部红树林区大型底栖动物种类组成与数量分布特点[J]. 厦门大学学报: 自然科学版, 1998, 37(1): 115-121. Cai Lizhe, Tan Fengyi, Huang Yushan. Characteristics of quantitative distribution and species composition of macrozoobenthos in mangrove stands in Eastern Hong Kong[J]. Journal of Xiamen University (Natural Science), 1998, 37(1): 115-121.
[8] 邹发生, 宋晓军, 陈伟, 等. 海南东寨港红树林滩涂大型底栖动物多样性的初步研究[J]. 生物多样性, 1999, 7(3): 175-180. Zou Fasheng, Song Xiaojun, Chen Wei, et al. The diversity of benthic macrofauna on mud flat in Dongzhaigang Mangrove Rserve, Hainan[J]. Chinese Biodiversity[J]. 1999, 7(3): 175-180.
[9] 张雅芝, 陈灿忠, 王渊源, 等. 福建红树林区底栖生物生态研究[J]. 生态学报, 1999, 19(6): 896-901. Zhang Yazhi, Chen Canzhong, Wang Yuanyuan, et al. The ecology of benthos in Fujian mangrove swamps[J]. Acta Ecologica Sinica, 1999, 19(6): 896-901.
[10] 厉红梅, 李适宇, 蔡立哲. 深圳湾潮间带底栖动物群落与环境因子的关系[J]. 中山大学学报: 自然科学版, 2003, 42(5): 93-97. Li Hongmei, LiShiyu, Cai Lizhe. Relationship between benthic community and environmental hactors in Shenzhen Bay[J].Acta Scientiarum Naturalium Universitatis Sunyatseni, 2003, 42(5): 93-97.
[11] 高爱根, 陈全震, 曾江宁, 等. 西门岛红树林区大型底栖动物的群落结构[J]. 海洋学研究, 2005, 23(2): 33-40. Gao Aigen, Chen Quanzhen, Zeng Jiangning, et al. Macrofauna community in the mangrove area of Ximen Island, Zhejiang[J]. Journal of Marine Sciences, 2005, 23(2): 33-40.
[12] 刘劲科, 韩维栋, 何秀玲, 等. 雷州半岛红树林海区底栖动物多样性的研究[J]. 海洋科学, 2006, 30(10): 65-69, 74. Liu Jingke, Han Weidong, He Xiuling et al. Studies on the biodiversity of benthic macrofauna of mangrove area in the Leizhou Peninsula, China[J].Marine Sciences, 2006, 30(10): 65-69, 74.
[13] 唐以杰, 方展强, 钟燕婷, 等. 不同生态恢复阶段无瓣海桑人工林湿地中大型底栖动物群落的比较[J]. 生态学报, 2012, 32(10): 3160-3169. Tang Yijie, Fang Zhan Qiang, Zhong Yanting, et al. Succession of macrofauna communities in wetlands ofartificial mangroves during different ecological restoration stages[J]. Acta Ecologica Sinica, 2012, 32(10): 3160-3169.
[14] Tang Y J, Fang Z Q, Chen K, et al. Ecological influence of exotic plants ofon understory macrofauna[J]. Acta Oceanologica Sinica, 2012, 31(5): 115-125.
[15] Tang Y J, Fang Z Q, Chang Z W, et al. Ecological indicators showing the succession of macrofauna communities inartificial mangrove wetlands on Qi’ao Island at Zhuhai, South China[J]. Acta Oceanologica Sinica, 2014, 33(8): 62-72.
[16] 陈光程, 余丹, 叶勇, 等. 红树林植被对大型底栖动物群落的影响[J]. 生态学报, 2013, 33(2): 0327-0336. Chen Guangcheng, Yu Dan, Ye Yong, et al. Impacts of mangrove vegetation on macro-benthic faunal communities[J]. Acta Ecologica Sinica, 2013, 33(2): 327-336.
[17] Kryger L, Lee S K. Effects of soil ageing on the accumulation of hydrogen sulphide and metallic sulphides in mangrove areas in Singapore[J]. Environment International, 1995, 21(1): 85-92.
[18] 何斌, 温远光, 刘世荣. 广西英罗港红树植物群落演替阶段的土壤化学性质[J]. 广西科学, 2001, 8(2): 148-151, 160. He Bin, Wen Yuanguang, Liu Shirong. Soil chemical properties at succession stages of mangrove communities in Yingluo Bay of Guangxi[J]. Guangxi Sciences, 2001, 8(2): 148-151, 160.
[19] [Gleason S M, Ewel K C, Hue N.Soil redox conditions and plant-soil relationships in a micronesian mangrove forest[J]. Estuarine, Coastal and Shelf Science, 2003, 56(5/6): 1065-1074.
[20] Lee H Y, Shih S S. Impacts of vegetation changes on the hydraulic and sediment transport characteristics in Guandu mangrove wetland[J].Ecological Engineering, 2004, 23(2): 85-94.
[21] 曹知勉, 叶勇, 卢昌义, 等. 红树林恢复对海岸湿地土壤影响的初步研究[J]. 生态科学, 2004, 23: 110- 113. Cao Zhimian, Ye Yong, Lu Changyi, et al. Preliminary studies on effects of restored mangroves on coastal soils[J]. Ecologic Science, 2004, 23: 110-113.
[22] 魏德重, 项长友, 鲍毅新, 等. 红树林种植对大型底栖动物群落结构及功能群的影响[J]. 浙江师范大学学报(自然科学版), 2012, 35(2): 195-202. Wei Dezhong, Xiang Changyou, Bao Yixin, et al. Effect of mangrove planting on macrobenthic community structure and functional group[J]. Acta Ecologica Sinica, 2012, 35(2): 195-202.
[23] 黄建荣, 刘启智, 赵一臣, 等. 珠海淇澳岛红树林蟹类区系[J]. 生态环境学报, 2011, 20(4): 730-736. Huang Jianrong, Liu Qizhi, Zhao Yichen, et al. The crab fauna in the mangrove of the Qi’Ao Island, Zhuhai[J]. Ecology and Environmental Sciences, 2011, 20(4): 730-736.
[24] 韩维栋, 凌大炯, 李燕, 等. 人工无瓣海桑林的土壤动态研究[J]. 南京林业大学学报(自然科学版), 2003, 27(2): 49-54. Han Weidong, Ling Dajiong, Li Yan, et al. The soil dynamic study of restoredplantations[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2003, 27(2): 49-54.
[25] Nobbs M. Effects of vegetation differ among three species of fiddler crabs (Uca spp.)[J]. Journal of Experimental and Marine Biology and Ecology, 2003, 284: 41-50.
[26] Vannini M, Cannicci S, Ruwa K. Effect of light intensity on vertical migrations of the tree crab,Hilgendorf (Decapoda, Grapisidae)[J]. Journal of Experimental and Marine Biology and Ecology, 1995, 185: 181-189.
[27] Suzuki T, Nishihira M, Paphavasit N. Size structure and distribution of(Gastropoda) in a Thai mangrove swamp[J]. Wetlands Ecology and Management, 2002, 10: 265-271.
[28] 刘玉, 余日清, 陈桂珠. 红树林区硅藻丰度和生物量构成及与底栖动物的关系[J]. 海洋环境科学, 2001, 20(3): 24-28. Liu Yu, Yu Riqing, Chen Guizhu. Abundance and biomass of diatom in mangrove areas and its relationship with benthos[J]. Marine Environmental Science, 2001, 20(3): 24-28.
[29] 徐姗楠, 陈作志, 李适宇. 红树林水生动物栖息地功能及其渔业价值[J]. 生态学报, 2010, 30(1): 186-196. Xu Shannan, Chen Zuozhi, Li Shiyu. The habitat functions of mangroves for aquatic fauna and fisheries[J]. Acta Ecologica Sinica, 2010, 30(1): 186-196.
[30] Ashton E C. Mangrove sesarmid crab feeding experiments in Peninsular malaysia[J]. Journal of Experimental and Marine Biology and Ecology, 2002, 273: 97-119.
[31] Lee C Y, Kwok P W. The importance of mangrove species association to the population biology of sesarmine crabs Parasesarma affinis and Perisesarma bidens[J]. Wetlands Ecology and Management, 2002, 10: 215-226.
[32] Erickson A A, Saltis M, Bell S S, et al. Herbivore feeding preferences as measured by leaf damage and stomatal ingestion: a mangrove example[J]. Journal of Experimental and Marine Biology and Ecology, 2003, 289: 123-138.
[33] Boswell C C, Espie P R. Uptake of moisture and nutrients by Hieracium pilosella and effects on soil in a dry sub-humid grassland[J]. New Zealand Journal of Agricultural Research, 1998, 41(2): 251-261.
[34] Macnae W. A general account of the fauna and flora of mangrove swamps and forests in the Indo-West-Pacific region. Advance in Marine Biology, 1969, 6: 73-103.
[35] 李旭林, 彭逸, 万如, 等. 两种相手蟹对不同红树植物叶片取食的偏好性[J]. 生态学报, 2010, 30(14): 3752-3759. Li Xulin, Peng Yi, Wan Ru, et al. Food preference of two sesarmid crabs to different mangrove leaves[J]. Acta Ecologica Sinica, 2010, 30(14): 3752-3759.
[36] 晏婷, 陈鹭真, 张宜辉, 等. 外来种无瓣海桑和乡土红树群落中大型底栖动物食物来源的比较研究[C]//中国第五届红树林学术会议论文摘要集. 2011: 15. Yan Ting, Chen Luzhen, Zhang yihui, et al.Comparative study of macrofauna’s food sources between exotic speciesand local mangrove communities[C]//The fifth mangrove academic conference abstract set of China. 2011: 15.
(本文编辑: 梁德海)
Comparison of macrofaunal communities in wetlands of three kinds of artificial mangroves in Shantou
TANG Yi-jie1, CHEN Si-min1, FANG Zhan-qiang2, LUO Li-fen1, CHEN Xiao-fen1, HUANG Hui-juan1, LU Di1
(1. School of biotechnology and food engineering, Guangdong University of Education, Guangzhou 510303, China; 2. Key Laboratory of Ecology and Environmental Science in Guangdong Higher Education, College of Life Sciences, South China Normal University, Guangzhou 510631, China)
This study was conducted to compare macrofaunal communities in the wetlands of three kinds of artificial mangroves of,andwhose restoration background had been the same, with those of the tidal flats at the Lianyang estuary of Shantou City, Guangdong Province. The results illustrated considerable differences in dominant species of macrofauna(>0.02)in four kinds of habitats. Additionally, the analysis of one-way ANOSIM illustrated considerable structural differences in macrofaunal communities from the four kinds of habitats. Further, the analysis of hierarchical clustering and non-metric multidimensional scaling showed that the structure of macrofaunal communities in the four habitats were considerably different. Among the three kinds of artificial mangroves, the macrobenthos community structure ofwas similar to that of. Moreover, the results showed that the order of species diversity and biomass of macrofauna at the three kinds of artificial mangroves was>>BIOENV analysis showed that the distribution of macrofauna was closely related to the physical and chemical factors of soil pH, organic matter, sand and silt content, average tree height, average diameter at breast height, average base diameter, and average crown (north and south × west and east) of mangrove plants. This further confirmed that the species diversity and distribution of macrofauna in wetlands of artificial mangroves were influenced by the species of mangrove plant.
;;; macrofauna
Oct. 18, 2015
Q958
A
1000-3096(2016)09-0053-08
10.11759/hykx20150124001
2015-10-18;
2016-01-15
国家自然科学基金项目(31570525); 广东省自然科学基金项目(2014A030313750); 广东大学生科技创新培育专项资金项目(pdjh2015a0394)
唐以杰(1972-), 男, 博士, 教授, 主要从事红树林底栖动物生态学研究, E-mail: tyj@gdei.e. du.cn; 方展强, 通信作者, E-mail: fangzhq@scnu.edu.cn
[Foundation: National Natural Science Foundation of China, No.31570525, Natural Science Foundation of Guangdong Province, No. 2014A030313750, The college students’ innovation cultivate project of science and technology of Guangdong Province, No. pdjh2015a0394]
