Cellular metabolic energy modulation by tangeretin in 7,12-dimethylbenz(a) anthracene-induced breast cancer
2016-12-13KuppusamyPeriyasamyVenkatachalamSivabalanKuppusamyBaskaranKannayiramKasthuriDhanapalSakthisekaran
Kuppusamy Periyasamy, Venkatachalam Sivabalan, Kuppusamy Baskaran, Kannayiram Kasthuri, Dhanapal Sakthisekaran
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Taramani Campus, Chennai-600 113, Tamil Nadu, India.
Cellular metabolic energy modulation by tangeretin in 7,12-dimethylbenz(a) anthracene-induced breast cancer
Kuppusamy Periyasamy, Venkatachalam Sivabalan, Kuppusamy Baskaran, Kannayiram Kasthuri, Dhanapal Sakthisekaran✉
Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Taramani Campus, Chennai-600 113, Tamil Nadu, India.
Breast cancer is the leading cause of death among women worldwide. Chemoprevention and chemotherapy play beneficial roles in reducing the incidence and mortality of cancer. Epidemiological and experimental studies showed that naturally-occurring antioxidants present in the diet may act as anticancer agents. Identifying the abnormalities of cellular energy metabolism facilitates early detection and management of breast cancer. The present study evaluated the effect of tangeretin on cellular metabolic energy fluxes in 7,12-dimethylbenz(a) anthracene (DMBA)-induced proliferative breast cancer. The results showed that the activities of glycolytic enzymes significantly increased in mammary tissues of DMBA-induced breast cancer bearing rats. The gluconeogenic tricarboxylic acid (TCA) cycle and respiratory chain enzyme activities significantly decreased in breast cancer-bearing rats. In addition, proliferating cell nuclear antigen (PCNA) was highly expressed in breast cancer tissues. However, the activities of glycolytic enzymes were significantly normalized in the tangeretin pre- and post-treated rats and the TCA cycle and respiratory chain enzyme activities were significantly increased in tangeretin treated rats. Furthermore, tangeretin down-regulated PCNA expression on breast cancerbearing rats. Our study demonstrates that tangeretin specifically regulates cellular metabolic energy fluxes in DMBA-induced breast cancer-bearing rats.
tangeretin, mitochondria, glycolysis, tricarboxylic acid cycle, proliferating cell nuclear antigen, breast cancer
Introduction
Breast cancer is the most common malignancy and the leading cause of death among women worldwide. Cancer cells have distinct metabolism and highly depend on glycolysis instead of mitochondrial oxidative phosphorylation. Hexokinase, phosphoglucoisomerase, aldolase and pyruvate kinase catalyze glucose through the glycolytic pathway, and have emerged as a potential regulator of the metabolic phenotype[2]. Warburg first pointed out that the metabolic disorder of cellular respiration is a common characteristic feature of cancer cells[3]. Under aerobic conditions, one glucose molecule is fully oxidized to produce CO2, H2O and 38 adenosine triphosphate (ATP). Cancer cellscan generate 2 molecules of lactic acid and 2 molecules of ATP through anaerobic respiration under hypoxic conditions. However, Warburg found that unlike normal cells, cancer cells generate energy through rapid glycolytic activity even in ample oxygen with the concomitant production of a large amount of lactic acid molecules. Oxidative phosphorylation is inhibited by the production of large amounts of lactic acid. This metabolic phenomenon is often called Warburg effect or aerobic glycolysis. Initially, Warburg thought that aerobic glycolysis was caused by the mitochondrial oxidative damages in cancer cells[4]. Other studies showed that under certain chemotherapeutical circumstances, cancer cells might have switched back to normal mitochondrial respiratory function. Chemotherapeutically-treated cancer cells grow slower than those with cancer-bearing conditions[5]. In these ways, aerobic glycolysis of cancer cells is likely to be controlled by the energy modulating effect of chemotherapeutic agents.
Cellular metabolic enzyme is regulated by many oncogenes and it serves a dual function as the speed regulator of cancer cell metabolism[6]. The activities of glycolytic enzymes are regulated by its own regulatory genes under the active proliferative signal. Pyruvate kinase splits the product of glycolysis into two parts, one for energy requirements and another for transformation into different precursor substances. Pyruvate kinase also forms a complex with other enzymes involved glycolysis and causes glucose to be degraded into pyruvate. It is further transformed into lactic acid, and also produces an ample amount of energy molecules. These energy molecules are used for the growth of tumor cells under hypoxic and anoxic conditions. The expression and translation of hexokinase, phosphoglucoisomerase, aldolase and pyruvate kinase are controlled by many oncogenes and metabolic intermediates[7]. Cancer cells acquire the rapid growth phenotype and ultimately develop mammary carcinoma through energy-producing molecules.
Tangeretin is a ubiquitous bioactive flavonoid in the peel of citrus fruits of the flavone family and also known to have anti-cancer, antioxidant, anti-inflammatory and hypolipidemic effects as well as antimicrobial activity. It is used in traditional Chinese medicine for treatment of various diseases including cancer[8]. In the present study, we evaluated the effect of tangeretin on cellular metabolic energy fluxes in 7,12- dimethylbenz(a) anthracene (DMBA)-induced proliferative breast cancer.
Materials and methods
Chemicals and reagents
Tangeretin (4',5,6,7,8-pentamethoxyflavone) was purchased from the Indofine Chemical Co. (Hillisborough, NJ, USA) and 7,12-dimethylbenz(a) anthracene (DMBA) and other fine chemicals were purchased from Sigma-Aldrich (St Louis, MO, USA). Primary monoclonal antibody PCNA was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Secondary antibodies, horseradish peroxidase (HRP) conjugated goat-antimouse IgG was obtained from Bio-Rad Laboratories (Hercules, CA, USA). The DAB kit was purchased from Genie, Bengaluru, Bangalore. Other chemicals and reagents in the study were of the highest purity and standard, which are commercially available.
Animals
Ninety day old healthy female Sprague-Dawley rats (Rattus norvegicus) weighing around 150 g were used in this study. The rats were housed in clean polypropylene cages, maintained in the air-conditioned animal house with a constant photoperiod of 12 hours light/ dark cycle with the light cycle from 6:00 to 18:00 and the dark cycle from 18:00 to 6:00. The rats were fed with the pellet diet and drinking water ad libitum. The study protocol was approved by the Institutional Animal Ethical Committee, University of Madras, Taramani Campus, Chennai, India (IAEC/No. 01/13/2012) and animal studies were carried out in accordance with the established institutional guidelines regarding animal care and use. Animal welfare and the experimental procedures were carried out strictly in accordance with the Guide for Care and Use of Laboratory Animals (National Research Council of USA, 1996).
Experimental design
The rats were divided into 5 groups, each consisting of 6 rats. Group I served as healthy control rats; Group II received a single dose of 80 mg/kg body weight of DMBA dissolved in 1 mL olive oil by gastric lavage. Breast cancer was allowed 90 days to develop and grow. Group III were pre-treated with tangeretin (50 mg/kg body weight orally) dissolved in olive oil for 30 days prior to administration of DMBA as in Group II. Group IV received DMBA as in Group II and after 90 days, were treated with tangeretin (50 mg/kg body weight) for 30 days orally. Group V rats were treated with tangeretin alone (50 mg/kg body weight) for 30 days and served as a drug control.
At the end of the experiment, blood was collected and rats were perfused with normal saline under sodium thiopentone (40 mg/kg body weight) anesthesia. Mammary and liver tissues were dissected out immediately. Histological evaluation of the samples was prepared in 10% buffered formalin and later embedded in paraffin. The sections were used forimmunohistochemistry. Fresh tissues were used for each experiment. Homogenate of breast tissues (10%) was prepared in 0.1 mol/L, pH 7.4 Tris-Hcl buffer using a Potter-Elvejhem glass homogenizer. Dilutions were performed according to protein concentration. The protein contents were measured by the method of Lowry et al.[9].
Isolation of mitochondria
Mitochondria were isolated from the breast tissues by the method of Johnson and Lardy[10]. Breast tissues were homogenized (10%) in ice-cold 0.25 mol/L sucrose with 50 mmol/L phosphate buffer, pH 7.4 with a Potter-Elvehjem glass homogenizer (Belco Glass, Inc., Vineland, NJ, USA) for 30 seconds. The homogenate was then centrifuged at 500×g for 10 minutes, and the supernatant was again centrifuged at 12,000×g for 15 minutes to obtain the mitochondrial fraction. The pellet, thus obtained, was resuspended in the buffer and used for assaying mitochondrial enzymes.
Hexokinase activity was assayed by the method of Brandstrup et al.[11]. Phosphoglucoisomerase was assayed according to Horrocks et al.[12]. Aldolase was estimated by the method of King[13]. The activities of glucose-6-phosphatase and fructose-1,6-disphosphatase were assayed by the method of Gancedo and Gancedo[14]. Pyruvate dehydrogenase activity was measured according to the method of Chretien et al.[15]. Mitochondrial isocitrate dehydrogenase (ICDH) and α-ketoglutarate dehydrogenase activities were measured according to the method of Duncan et al.[16]. Mitochondrial succinate dehydrogenase activity was measured according to the method of Veeger et al.[17]. NADH-cytochrome c oxidoreductase and cytochrome c oxidase activities were determined by the method of Goyal and Srivastava[18].
Immunohistochemistry
Immunohistochemical analysis of PCNA expression was performed on 5 µm thick paraffin embedded breast tissue section on poly-L-lysine coated glass slides. The tissue slides were deparaffinized by placing the slides in an oven at 60°C for 10 minutes and then rinsed twice in xylene for 5 minutes each. The slides were hydrated in a graded ethanol series for 10 minutes each. Then, the slides were washed in double distilled water for 5 minutes. The sections were incubated with 1% H2O2in double distilled water for 15 minutes at 22°C, to quench endogenous peroxidase activity. The sections were rinsed with Tris-HCl containing 150 mmol/L NaCl (pH 7.4) and blocked with blocking buffer (1×TBS, 0.05% Tween 20, 5% NFDM) for 1 hour at 22°C. After washing with 1×TBS containing 0.05% Tween 20, the sections were incubated with primary antibodies PCNA (1:200 dilution) overnight at 4°C, followed by incubation with the respective secondary antibodies IgG-HRP conjugates for 1 hour at 4°C. After washing with 1×TBS containing 0.05% Tween 20, immune-reactivity was developed with 0.01% DAB and H2O2for 1–3 minutes and the sections were observed (20×) for brown color formation under bright field in a microscope. The sections were photographed using Nikon 80i Eclipse microscope.
Statistical analysis
The values are expressed as mean ± SEM. The results were computed statistically (SPSS software package, Version 17) using one-way analysis of variance (ANOVA). The post-hoc testing was performed for inter-group comparison using the Tukey multiple comparison test. P < 0.05 was considered statistically significant.
Results
Glycolytic enzyme activities
Glycolytic activities in mammary tumors were determined. The glycolytic enzyme activities of DMBA-induced breast cancer cells exhibited substantial increases in cancer-bearing rats. Table 1 shows the activities of glycolytic enzymes in mammary tissues of rats. The activities of hexokinase, phosphoglucoisomerase and aldolase levels significantly increased (P < 0.05 vs. control) in DMBA-induced breast cancer-bearing rats. However, tangeretin treatment for 30 days before the administration of DMBA in Group III rats and post-treatment with tangeretin in breast cancer-bearing Group IV rats significantly reduced the activities of hexokinase, phosphoglucoisomerase, and aldolase compared with tumor-bearing rats (Group II). Group V showed no effective changes (P > 0.05) compared with the control rats (Group I).
Gluconeogenic enzymes
The estimated gluconeogenic activities in the liver tissue of DMBA-induced breast cancer along with tangeretin pre-and post-treated rats are shown in Table 2. The activities of gluconeogenic enzymes such as fructose-1,6-bisphosphatase and glucose-6-phosphatase significantly decreased (P < 0.05) in DMBA-induced breast cancer-bearing rats when compared with the control rats (Group I). However, tangeretin pre-treated rats showed a significant increase of fructose-1,6-bisphosphataseand glucose-6-phosphatase activities (P < 0.05 vs. induced). Furthermore, post-treatment with tangeretin (50 mg/kg body weight) for 30 days significantly restored the activities of fructose-1,6-bisphosphatase and glucose-6-phosphatase in Group IV compared with cancer-bearing rats (P < 0.05). No significant enzyme activities were found in rats treated with tangeretin alone (P < 0.05 vs. control).

Table 1 Effects of tangeretin on the glycolytic activities in the mammary tissues of rats with DMBA-induced breast cancer
Mitochondrial TCA cycle enzymes
Fig. 1 shows a significant decrease in the activity of pyruvate dehydrogenase (PDH), which couples glycolysis to the TCA cycle, in DMBA-induced breast cancer-bearing rats (P < 0.05 vs. control). The activities of PDH were significantly ameliorated (P < 0.05) in tangeretin pre-and post-treated against breast cancer-bearing rats when compared with Group II rats. The mitochondrial TCA cycle enzymes such as isocitrate dehydrogenase, α-ketoglutarate dehydrogenase and succinate dehydrogenase activities were assayed according to the standard protocols. Fig. 1 shows the activities of ICDH, α-KGDH and SDH in the mammary tissue of rats. TCA cycle enzymes were significantly reduced (P < 0.05) in Group II compared with the control rats. However, tangeretin pre-treatment and post-treatment on DMBA-induced breast cancer-bearing rats showed a significant increase of ICDH, α-KGDH and SDH activities with Group II. None of the changes were observed in rats treated with tangeretin alone compared with the control rats.
Mitochondrial respiratory chain enzymes
The activities of mitochondrial respiratory chain enzymes like NADH-cytochrome c oxidoreductase and cytochrome c oxidase in mitochondria are shown in Fig. 2A and 2B, respectively. The mitochondrial activities of NADH cytochrome c oxidoreductase and cytochrome c oxidase were significantly decreased (P < 0.05) in the mammary tissues of rats with DMBA-induced breast cancer. However, Fig. 2C shows the activity of cytochrome c oxidase in the cytosol of DMBA-induced breast cancer cells, and there was a significant increase (P < 0.05) of cytochrome c oxidase in cytosol due to leakage from the mitochondria into the cytosol compared with the control rats. Following treatment with tangeretin in Group III and IV rats, the activities of NADH-cytochrome c oxidoreductase and cytochrome c oxidase significantly increased (P < 0.05) in the mitochondria. Conversely, the activity of cytochrome c oxidase in tangeretin treatment was significantly decreased in the cytosol. These data are presented in Fig. 2.
PCNA
Fig. 3 shows the immunohistochemical expression of PCNA in breast cancer tissues. PCNA was highly expressed in DMBA-induced breast cancer-bearingmammary tissues. However, PCNA expression was significantly decreased in tangeretin pre- and posttreated mammary tissues when compared with breast cancer-bearing mammary tissue. These results indicated that tangeretin may have an antiproliferative effect on DMBA-induced breast cancer.
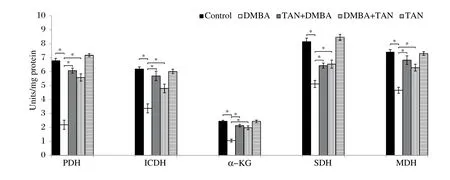
Fig. 1 Protective role of tangeretin on the TCA cycle activities in mammary tissues. The activities of pyruvate dehydrogenase, isocitrate dehydrogenase, α-ketoglutarate dehydrogenase and succinate dehydrogenase are depicted in the Fig. 1. Values are mean ± SEM of 6 rats in each group. The significant level is *P < 0.05.
Discussion
Our previous study showed that tangeretin plays a significant role against oxidative stress-induced proliferative breast cancer[8]. In the present study, we analyzed the effect of tangeretin on cellular energy metabolic variation in rats. We found thatthe cytoplasmic glycolytic enzymes accelerated high-energy production. Furthermore, there was a low-energy production through the TCA cycle as well as the mitochondrial respiratory chain system in mammary cells due to mitochondrial oxidative stress.
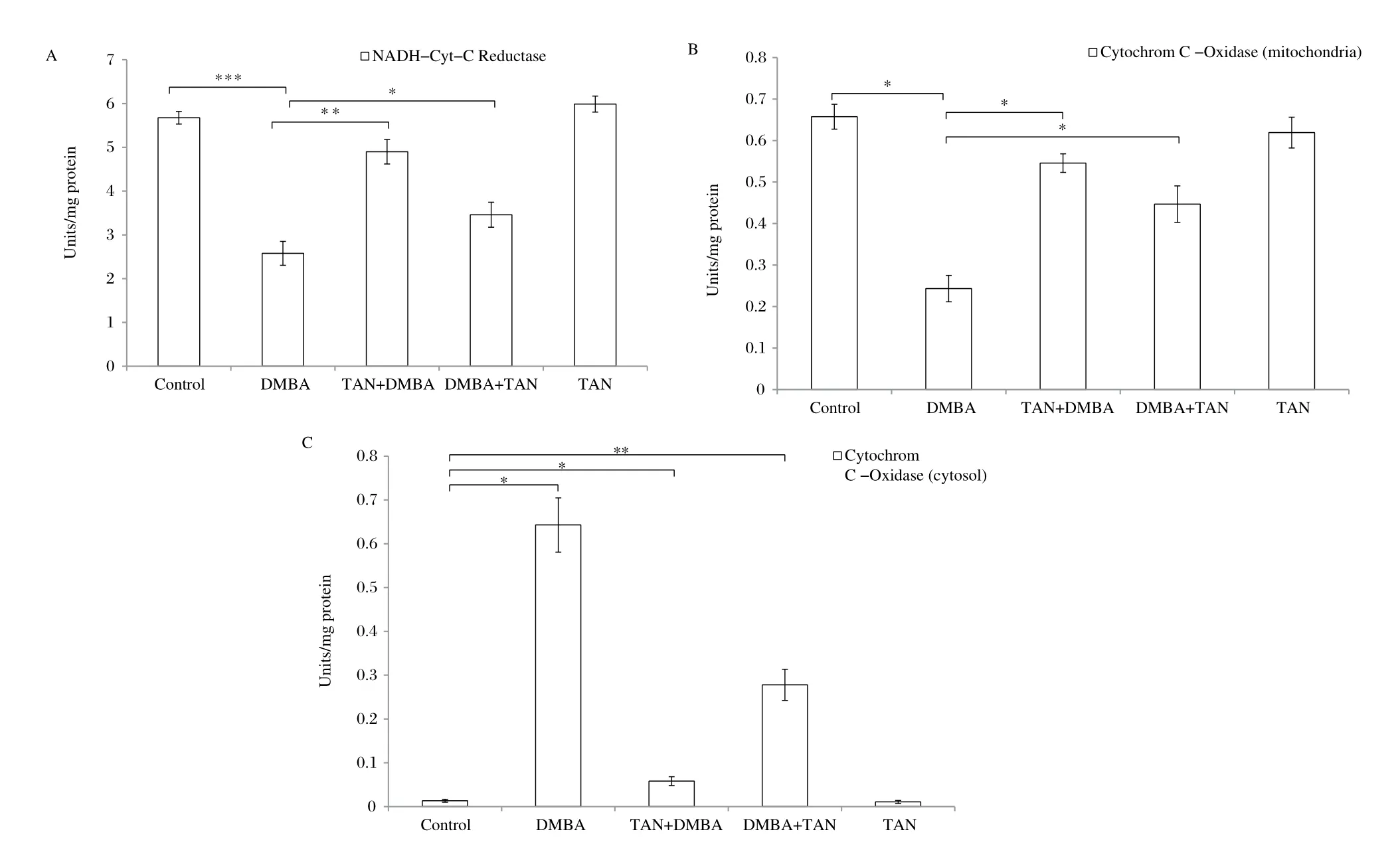
Fig. 2 Effect of tangeretin on mitochondrial respiratory enzyme activities in mammary tissues. The activities of 2(a) NADH-cytochrome-c oxidoreductase, 2(b) cytochrome-c oxidase (mitochondria) and 2(c) cytochrome-c oxidase (cytosol) were determined and values are mean ± SEM of 6 rats in each group. The significant level is *P < 0.05, **P < 0.01.
The elevated cellular metabolic energy levels account for abnormal metabolic properties of tumors. Increased rate of glycolysis in tumor cells leads to an increase in intracellular concentration of glucose-6-phosphate. It is a key precursor in de novo synthesis of nucleic acids, phospholipids and other macromolecules[20]. Increased synthesis of glycolytic components may be essential to keep rapid cell division and membrane biosynthesis during tumor growth. Thus, rapid proliferating tumor cells have an excessive energy demand for nuclear cell division[21]. Phosphoglucoisomerase acts as a catalyst in the conversion of glucose-6-phosphate to fructose-6-phosphate and it is an indicator of metastasis. Alterations in the activities of phosphoglucoisomerase might be expected by the influence of a proportion of glucose 6-phosphate, which is metabolized via the glycolytic pathway[22]. Hennipnan et al. reported increased activity of aldolase in metastatic tumor. Upon tangeretin treatment, breast cancer cells could decrease glycolytic regulatory enzyme activities, which might be due to production of high amounts of glucose-6-phosphate by repairing defective mitochondria. Previous studies reported that natural flavonoids could inhibit glycolysis in tumor cells. Tangeretin inhibits accelerated glycolytic activities in cancer and also induces the apoptotic cell death in malignant tumor cells[23].
In the present study, we have observed that the activities of gluconeogenic enzymes were significantly lowered in the liver tissue of DMBA-induced breastcancer-bearing rats. Reduction rate of fructase-1,6-bisphosphatase might lead to a higher concentration of its substrate fructose-1,6-bisphosphate, which is an allosteric activator of pyruvate kinase. Alternatively, down regulation of glucose-6-phosphatase activity is likely to increase the concentration of its substrate glucose-6-phosphate, which might be used by pentose phosphate shunt pathway to produce ribose-5-phosphate for rapid nucleotide synthesis in cancer[24]. The diminished activities of fructose-1,6- bisphosphatase and glucose-6-phosphatase were due to higher concentration of lactic acid production in neoplastic tissues. Higher rate of glucose utilization and an increased production of lactic acid are characteristic features of neoplastic cells[25]. Gluconeogenesis from lactate on the other hand is an essential energy- requiring process as pointed out by Fenninger and Mider. Gluconeogenesis may play an important role in excessive energy expenditure of the host; thus, it is contributing to promote weight loss[26]. Tangeretin effectively acts on proliferative cancer cells by ameliorating the gluconeogenic activities.
Mitochondria are considered as the major source of cellular ROS, which may inhibit mitochondrial Krebs cycle enzyme activities[27]. The mitochondrial TCA cycle enzyme activities were significantly lowered in cancer, which is due to membrane damages by the oxidative stress-induced free radicals. Formation of ROS disturbs the nitric oxide pathway, resulting in protein nitration, oxidative damage of DNA and lipid peroxidation. However, it is more and more evident that alterations of energy metabolism in cancer are especially due to mitochondrial dysfunction. Decreased activities of ICDH, α-KGDH and SDH could be due to the alteration in mitochondrial membranes, morphology and organelles from peroxidation of macromolecules[28]. Tangeretin plays a major role in restoring the mitochondrial function on DMBA-induced breast cancer-bearing rats.
Inhibition of the mitochondrial TCA cycle and respiratory chain enzyme activities could lead to a leakage of electrons, which produces a reducing environment within the mitochondria, thereby generating free radicals[29]. Reduction activities of NADH cytochrome c oxidoreductase and cytochrome c oxidase were noted in DMBA-induced breast cancer. It clearly indicates that an elevated state of oxidative stress could lead to impaired regulatory functions of mitochondria. These free radicals, particularly free •OH radical, can damage the mitochondrial membrane, which causes leakage of cytochrome c from the mitochondria into the cytosol[30]. Our current study showed that tangeretin could inhibit the highly significant release of cytochrome c in the cytosol from mitochondria. The reason for this may lie in the fact that tangeretin may be able to repair mitochondrial membrane damage. Tangeretin could increase mitochondrial enzymes, which are related to energy metabolism. Tangeretin could completely restore NADH cytochrome c oxidoreductase and cytochrome c oxidase. It indicates that tangeretin is capable of mitigating mitochondrial oxidative stress-induced proliferation of breast cancer cells.
Cellular proliferation of the mammary tissues was observed by immunohistochemical examination of PCNA expression. PCNA was overexpressed in 80% of breast cancer according to a previous study[31]. In our study, the protein expression of PCNA significantly increased due to potential mutagenic activity of DMBA. However, tangeretin drastically decreased PCNA expression, due to the inhibition of rapid DNA synthesis. PCNA primarily expressed during the cell cycle progression and its rate of synthesis is directly correlated with proliferative cancer cells. PCNA is a cell nuclear protein, and it functions as an auxiliary protein for DNA polymerase delta. Tangeretin inhibits the cellular G1, G1/S phases in the cell cycle systemin cancer progression[32]. Juhee et al. also reported that tangeretin significantly inhibited proliferation, DNA synthesis and migration of aortic smooth muscle cells by blocking AKT activation[33]. In that way, anti-proliferative activity of tangeretin control proliferative breast cancer.
In conclusion, pre- and post-treatment with tangeretin in breast cancer could prevent abnormal carbohydrate metabolic energy fluxes and brought back to normal the functions of mitochondria in energy metabolism.
Acknowledgement
The financial support from the University Grants Commission, New Delhi, in the form of UGC-BSR Research Fellowship under the UGC-SAP-DRS-III programme is gratefully acknowledged.
References
[1] Ferlay J, Shin HR, Bray F, et al. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008[J]. Int J Cancer, 2010,127(12):2893-2917.
[2] Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science (New York, NY), 2009,324(5930):1029-1033.
[3] Warburg O. On respiratory impairment in cancer cells[J]. Science, 1956,124(3215):269-270.
[4] Warburg O, Dickens F. Kaiser Wilhelm-Institut fur Biologie B. The Metabolism of Tumours: Investigations from the Kaiser-Wilhelm Institute for Biology, Berlin-Dahlem. London, UK: Constable & Co, Ltd; 1930.
[5] Mazurek S. Pyruvate kinase type M2: a key regulator of the metabolic budget system in tumour cells[J]. Int J Biochem Cell Biol, 2011,43:969-980.
[6] Wu S, Le H. Dual roles of PKM2 in cancer metabolism[J]. Acta Biochim Biophys Sin (Shanghai), 2013,45(1): 27-35.
[7] Wang X, Peralta S, Moraes CT. Mitochondrial alterations during carcinogenesis: a review of metabolic transformation and targets for anticancer treatments[J]. Adv Cancer Res, 2013,119:127-160.
[8] Periyasamy K, Baskaran K, Ilakkia A, et al. Antitumor efficacy of tangeretin by targeting the oxidative stress mediated on 7,12-dimethylbenz(a) anthracene-induced proliferative breast cancer in Sprague-Dawley rats[J]. Cancer Chemother Pharmacol, 2015,75(2):263-272.
[9] Lowry O, Rosebrough N, Farr A, et al. Protein measurement with the Folin phenol reagent[J]. J Biol Chem, 1951,193:265-275.
[10] Johnson, D, Lardy H. Methods in Enzymology (Estabrook RW and Pullman ME, eds. 1967,10:94-96.
[11] Brandstrup N, Kirk JE, Bruni C. The hexokinase and phosphoglucoisomerase activities of aortic and pulmonary artery tissue in individuals of various ages[J]. J Gerontol, 1957,12(2):166-171.
[12] Horrocks JE, Ward J, King J. A routine method for the determination of phosphoglucoseisomerase activity in body fluid[J]. J Clin Pathol, 1963,16:248-251.
[13] King J. Practical clinical enzymology. Van Nostrand, Science 1965, p. 363.
[14] Gancedo JM, Gancedo C. Fructose-1,6-diphosphatase, phosphofructokinase and glucose-6-phosphate dehydrogenase from fermenting and non fermenting yeasts[J]. Arch Mikrobiol, 1971,76(2):132-138.
[15] Chretien D, Pourrier M, Bourgeron T, et al. An improved spectrophotometric assay of pyruvate dehydrogenase in lactate dehydrogenase contaminated mitochondrial preparations from human skeletal muscle[J]. Clin Chim Acta, 1995,240(2):129-136.
[16] Duncan MJ, Fraenkel DG. Alpha-ketoglutarate dehydrogenase mutant of Rhizobium meliloti[J]. J Bacteriol, 1979,37:415-419.
[17] Veeger C, Vartanian DV, Zeylemaker WP. Succinate dehydrogenase[J]. Meth Enzymol, 1969,13:81-90.
[18] Goyal N, Srivastava VML. Oxidation and reduction of cytochrome c by mitochondrial enzymes of Setariacervi[J]. J Helminth, 1995,69:13-17.
[19] Purushothaman A, Nandhakumar E Sachdanandam P. Phytochemical analysis and anticancer capacity of Shemamruthaa, a herbal formulation against DMBA-induced mammary carcinoma in rats[J]. Asian Pac J Trop Med, 2013,6(12):925-933.
[20] Boros LG, Lee WN, Go VL. A metabolic hypothesis of cell growth and death in pancreatic cancer[J]. Pancreas, 2002,24(1):26-33.
[21] Senthilnathan P, Padmavathi R, Magesh V, et al. Modulation of TCA cycle enzymes and electron transport chain systems in experimental lung cancer[J]. Life Sci, 2006,78(9):1010-1014.
[22] Ebrahim GJ. Health care of adolescents[J]. J trop Pediatr, 1996,42(6):316-317.
[23] Hennipman A, Van Oirschot BA, Smits J, et al. Glycolytic enzymes activities in breast cancer metastases[J]. Tumour Biol, 1988,9(5):241-248.
[24] Wang B, Hsu SH, Frankel W, et al. Stat3-mediated activation of microRNA-23a suppresses gluconeogenesis in hepatocellular carcinoma by down-regulating glucose-6-phosphatase and peroxisome proliferator- activated receptor gamma, coactivator 1 alpha[J]. Hepatology, 2012,56:186-197.
[25] Waterhouse C. Lactate Metabolism in patients with Cancer[J]. Cancer 1974,33(1):66-71.
[26] Fenninger LD and Mider GB. Energy and Nitrogen metabolism in cancer[J]. Adv Cancer Res, 1954,2:229-253.
[27] Fariss MW, Chan CB, Patel M, et al. Role of mitochondria intoxic oxidative stress. Mol Interv, 2005,5(2):94-111.
[28] Dimitrova-Shumkovska J, Veenman L, Ristoski T, et al. Decreases in binding capacity of the mitochondrial 18 kdatranslocator protein accompany oxidative stress and pathological signs in rat liver after DMBA exposure[J]. Toxicol Pathol, 2010,38(6):957-968.
[29] Martin M, Macias M, Leon J, et al. Melatonin increases the activity of the oxidative phosphorylation enzymes and the production of ATP in rat brain and liver mitochondria[J]. Int J Biochem Cell Biol, 2002,34:348-357.
[30] Reiter RJ, Tan DX, Mancshester LC, et al. Melatonin reduces oxidant damage and promotes mitochondrial respiration: implications for aging[J]. Ann N Y Acad Sci, 2002,959:238-250.
[31] Urruticoechea A, Smith IE, Dowsett M. Proliferation marker Ki-67 in early breast cancer[J]. J Clin Oncol, 2005,23(28):7212-7220.
[32] Pan M, Chen W, Lin-Shiau S, et al. Tangeretin induces cell-cycle G1 arrest through inhibiting cyclin-dependent kinases 2 and 4 activities as well as elevating Cdk inhibitors p21 and p27 in human colorectal carcinoma cells[J]. Carcinogenesis, 2002,23(10):1677-1684.
[33] Juhee S, Hyun SL, Sungwoo R, et al. Tangeretin, a citrus flavonoid, inhibits PGDF-BB-induced proliferation and migration of aortic smooth muscle cells by blocking AKT activation[J]. Eur J Pharmacol, 2011,673:56-64.
✉ Prof. D. Sakthisekaran, Former Professor and Head, UGC-BSR-Faculty Fellow, Department of Medical Biochemistry, Dr. ALM Post Graduate Institute of Basic Medical Sciences, University of Madras, Taramani Campus, Chennai-600113, Tamil Nadu, India. Tel/Fax: +91-44-24547083/+91-44-24540709,
Email: dsakthisekaran@hotmail.com.
04 April 2015, Revised 02 June 2015, Accepted 10 October 2015, Epub 26 February 2016
R730.22, Document code: A
The authors reported no conflict of interests.
杂志排行

THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Organ donation in China: the major progress and the continuing problem
- Cerebral ischemia during surgery: an overview
- Apolipoprotein A-V gene therapy for disease prevention / treatment: a critical analysis
- HDL signaling and protection against coronary artery atherosclerosis in mice
- Prevalence and risk factors of HIV and syphilis, and knowledge and risk behaviors related to HIV/AIDS among men who have sex with men in Chongqing, China
- Effects of closing and reopening live poultry markets on the epidemic of human infection with avian influenza A virus