硫化物胁迫对凡纳滨对虾血细胞抗氧化酶基因表达的影响
2016-12-09汪蕾张秀霞郑佩华鲁耀鹏冼健安
汪蕾, 张秀霞, 郑佩华, 鲁耀鹏, 冼健安*
(1.华南师范大学药物研究院,广州510631; 2. 中国热带农业科学院热带生物技术研究所,海口571101)
硫化物胁迫对凡纳滨对虾血细胞抗氧化酶基因表达的影响
汪蕾1, 张秀霞2, 郑佩华1, 鲁耀鹏1, 冼健安2*
(1.华南师范大学药物研究院,广州510631; 2. 中国热带农业科学院热带生物技术研究所,海口571101)
硫化物是水产养殖过程中常见的水体污染物之一,为探讨抗氧化酶在凡纳滨对虾Litopenaeusvannamei血细胞抗硫化物胁迫中的作用,以不同浓度(0 mg·L-1和2.0 mg·L-1)的硫化物对凡纳滨对虾进行胁迫,于胁迫后的0 h、6 h、12 h、24 h和48 h取血淋巴,测定血细胞中铜锌超氧化物歧化酶(Cu,Zn-SOD)、锰超氧化物歧化酶(Mn-SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)和硫氧还蛋白(TRx)基因表达量的变化。结果显示,Cu,Zn-SOD和TRx的表达量在胁迫的6~24 h显著上升,在48 h时恢复至对照组水平;Mn-SOD和GPx的表达量在胁迫的6~48 h均显著上调;CAT表达水平在胁迫的12 h开始有显著提高。这些结果表明凡纳滨对虾血细胞抗氧化相关基因的表达均被诱导上调,以进行防御;Cu,Zn-SOD、Mn-SOD、GPx和TRx均对硫化物胁迫敏感,在胁迫前期发挥作用,CAT主要在胁迫后期发挥作用。
硫化物;凡纳滨对虾;血细胞;抗氧化酶;基因表达
硫化物是水产养殖中常见的水体有毒污染物之一,在养殖池塘中主要有2个来源:(1)硫酸盐在硫酸盐还原菌的作用下异化还原形成;(2)水产动物的代谢物、残饵等有机质中的含硫氨基酸被微生物分解利用而产生。硫化物对水产动物具有较强的毒性,但目前针对硫化物对虾类毒性影响的研究较少,主要集中在半致死浓度以及对酶活性影响的研究上(Chengetal.,2007;Hsu & Chen,2007;李建等,2007;管越强等,2009,2011;於叶兵等,2011;余静等,2011;杨世平等,2014)。笔者前期应用流式细胞术分析了硫化物对凡纳滨对虾Litopenaeusvannamei细胞毒性的影响,结果显示硫化物胁迫刺激其血细胞产生大量活性氧(ROS),诱导血细胞凋亡,继而导致血细胞总数(THC)下降,这可能是硫化物对虾类产生细胞毒性的重要机制之一(汪蕾等,待发表)。抗氧化酶是机体内清除过量ROS,进行抗氧化防御的重要工具。根据前期研究结果,笔者推测抗氧化酶可能在抗硫化物胁迫过程中发挥重要作用。因此,本研究针对5种重要的抗氧化酶:铜锌超氧化物歧化酶(Cu,Zn-SOD)、锰超氧化物歧化酶(Mn-SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)和硫氧还蛋白(TRx),分析硫化物胁迫对其在血细胞中表达水平的影响,探讨它们在抗硫化物胁迫中发挥的作用,为进一步研究如何提高虾类的抗硫化物胁迫能力提供理论基础。
1 材料和方法
1.1 材料与试剂
凡纳滨对虾购自广东省广州市番禺区某养殖场。对虾平均体质量为8.69 g±0.75 g,在实验室环境条件下驯养,养殖用水的条件为:盐度20‰,pH7.9~8.0,温度22~24 ℃,一直保持曝气,并进行循环过滤。驯养1周后,选取健康无病患、附肢完整、处于蜕皮间期的对虾进行实验。
DNA marker、PCR试剂盒(Taq Mix)购自东盛公司;Trizol RNA提取试剂、反转录试剂盒(PrimeScriptTMRT reagent Kit)、荧光定量试剂盒(SYBR Premix Ex TaqTM)均购自大连TaKaRa公司;焦炭酸二乙酯(DEPC)和琼脂粉均购自上海生工生物工程有限公司;其他试剂为国产分析纯。所有使用的器具和耗材经过RNase free处理。
1.2 硫化物胁迫
以硫化钠作为硫化物来源,配制硫浓度为1 g·L-1的硫化钠溶液,用于调节水体硫化物浓度。根据参考文献(Hsu & Chen,2007)及前期研究结果,设置硫化物胁迫浓度为2.0 mg·L-1,硫化物浓度采用亚甲基蓝分光光度法测定,实际测定浓度为1.92 mg·L-1±0.09 mg·L-1,对照组为不添加组。每组设3个平行,每个养殖桶放盐度20‰的水180 L,放养对虾20尾。为保证硫化物浓度,每6 h换水1次,换水量约为50%。分别在胁迫的0 h、6 h、12 h、24 h和48 h取样。
1.3 血细胞总RNA提取
用2.5 mL一次性注射器吸取预冷的抗凝剂(葡萄糖20.5 g·L-1、柠檬酸钠8 g·L-1、氯化钠4.2 g·L-1,pH7.5),然后从对虾的围心腔抽取等量的血淋巴,3 000 r·min-1, 4 ℃离心10 min,弃上清,沉淀为血细胞,用于提取总RNA。应用Trizol RNA提取试剂提取总RNA,具体方法参考说明书。得到的RNA样品用1%琼脂糖凝胶电泳检测完整性,取2 μL利用NanoDrop ND-100 Spectrophotometer测定RNA浓度和纯度,OD260/280在1.9~2.0的RNA作为反转录的模板。
1.4 cDNA第一链的合成
根据RNA浓度用DEPC水稀释,在10 μL的反转率体系中加入500 ng RNA作为反转录模板。实验使用TaKaRa公司的PrimeScriptTMRT reagent Kit (Perfect Real Time)试剂盒,按照说明书步骤进行。
1.5 表达定量
从美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)数据库查找所需基因的cDNA序列,根据查找到的序列利用Primer Premier 5进行引物的设计或者参考文献中的引物,各基因的GenBank序列号和引物序列见表1,所有引物由上海生工生物工程技术服务有限公司合成。实时荧光定量PCR实验使用TaKaRa公司的SYBR Premix Ex TaqTM试剂盒,实验步骤按照说明书进行。所得数据以ABI 7500 SDS进行分析,以β-actin作为内参,根据目的基因和β-actin的Ct值用2-ΔΔCt法计算目的基因mRNA的相对表达量。每个样品做3个重复。
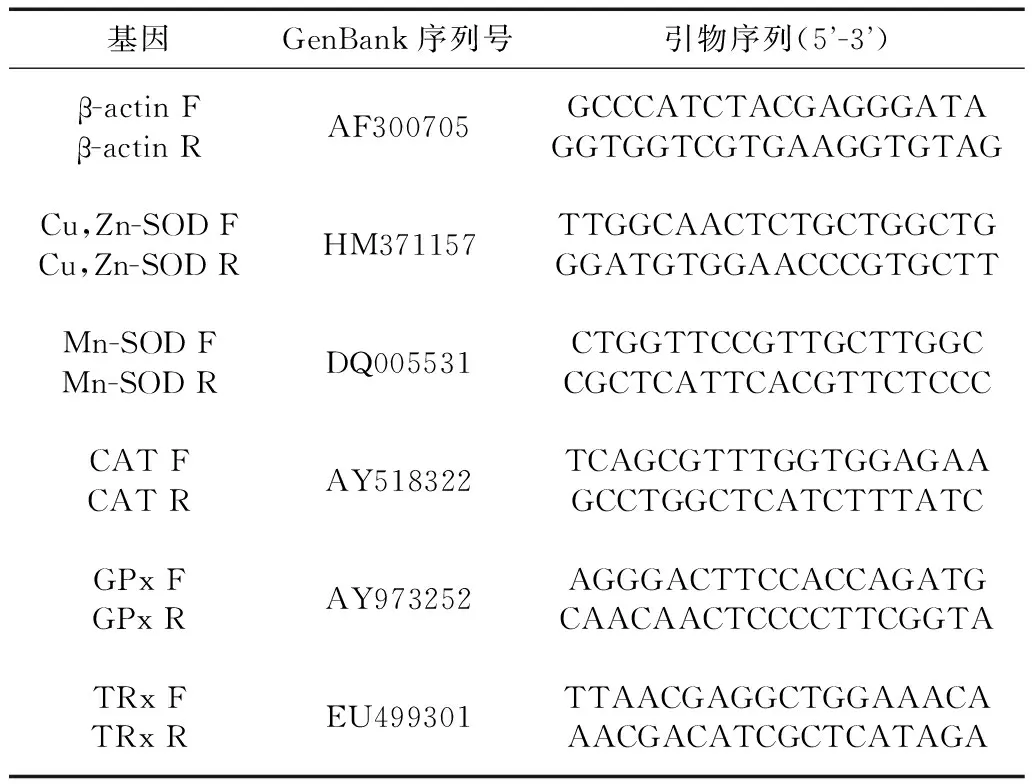
表1 实时荧光定量PCR引物信息
1.6 统计分析
结果显示为平均值±标准差(Mean±SD),实验数据利用SPSS 19.0进行统计单因素方差分析(One-Way ANOVA),显著性水平α=0.05。
2 结果
2.1 对Cu,Zn-SOD表达量的影响
硫化物胁迫对Cu,Zn-SOD表达水平的影响见图1。与对照组相比,胁迫6 h、12 h和24 h对虾血细胞中Cu,Zn-SOD的表达水平显著提高(P<0.05),并在12 h时达到最大值,为对照组的8.16倍;48 h时,Cu,Zn-SOD的表达量下降至对照组水平(P>0.05)。
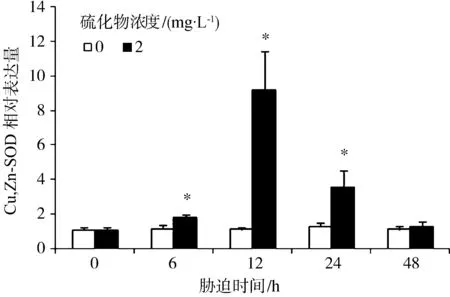
图1 凡纳滨对虾经硫化物胁迫后的血细胞中Cu,Zn-SOD基因表达量
2.2 对Mn-SOD表达量的影响
硫化物胁迫对Mn-SOD表达水平的影响见图2。胁迫6 h、12 h、24 h和48 h对虾血细胞中的Mn-SOD表达水平均显著升高(P<0.05);在6 h时达到最大值,为对照组的14.73倍。
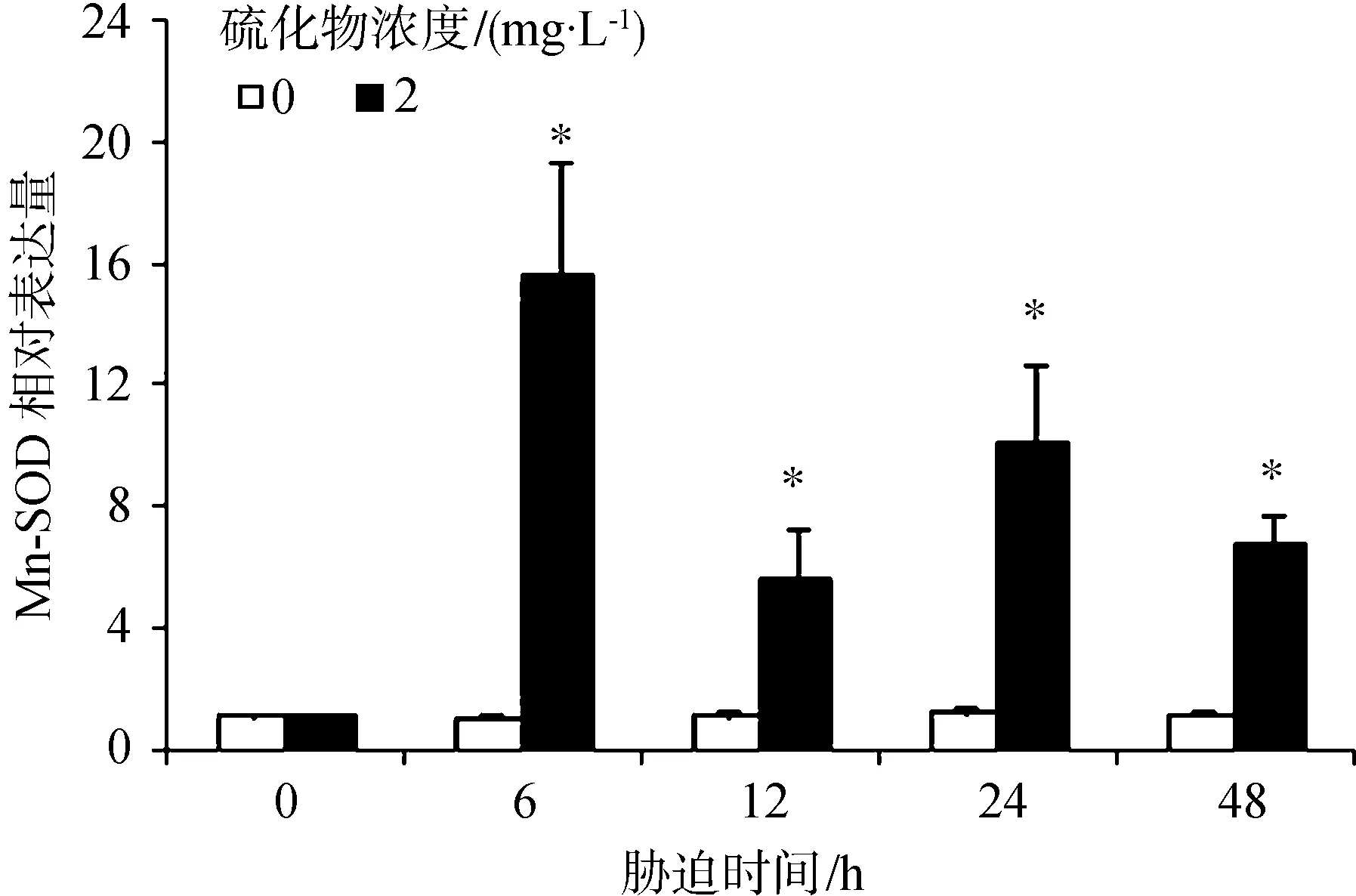
图2 凡纳滨对虾经硫化物胁迫后的血细胞中Mn-SOD基因表达量
2.3 对CAT表达量的影响
硫化物胁迫对CAT表达水平的影响见图3。胁迫6 h对虾血细胞中的CAT表达量与对照组的差异无统计学意义(P>0.05);胁迫12 h、24 h和48 h对虾血细胞中的Mn-SOD表达量均显著升高(P<0.05),在24 h时达到最大值,为对照组的13.78倍。
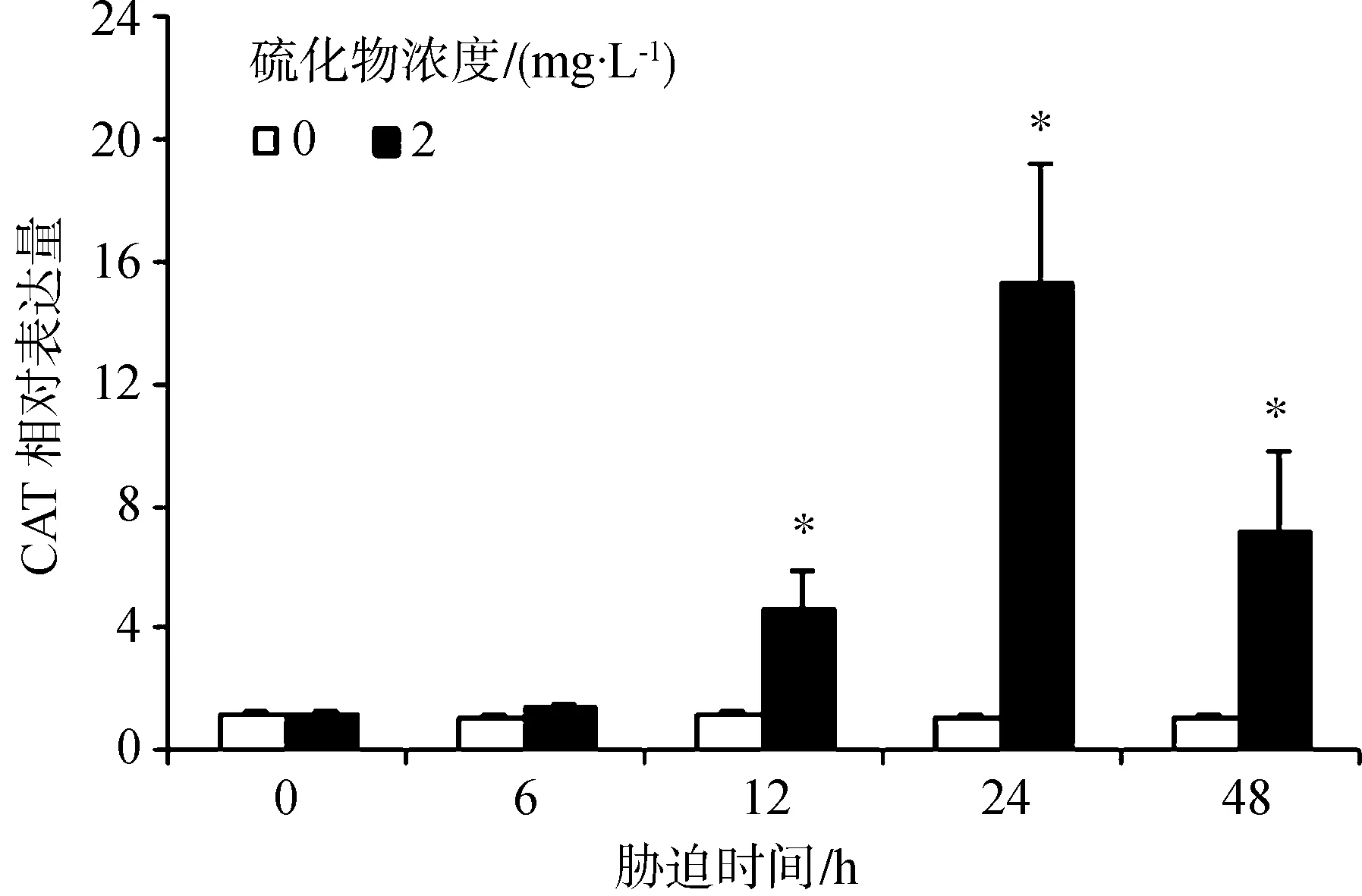
图3 凡纳滨对虾经硫化物胁迫后的血细胞中CAT基因表达量
2.4 对GPx表达量的影响
硫化物胁迫对GPx表达水平的影响见图4。胁迫6 h、12 h、24 h和48 h对虾血细胞中的GPx表达水平均显著高于对照组(P<0.05),在6 h时达到最大值,为对照组的7.80倍。
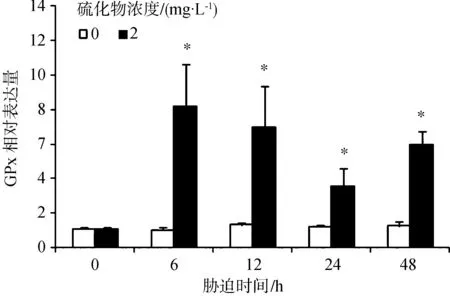
图4 凡纳滨对虾经硫化物胁迫后的血细胞中GPx基因表达量
2.5 对TRx表达量的影响
硫化物胁迫对TRx表达水平的影响见图5。胁迫6 h、12 h和24 h对虾血细胞中的TRx表达量显著高于对照组(P<0.05),在12 h时达到最大值,为对照组的2.84倍;48 h时,TRx的表达量下降至对照组水平(P>0.05)。
3 讨论
众多研究表明,氧化损伤是水体环境因子对水产动物产生胁迫毒害作用的普遍机制之一,如亚硝酸盐(郭慧等,2015)、pH(Maietal.,2010)和重金属(Wangetal.,2013)等。笔者前期研究也发现硫化物胁迫会诱导凡纳滨对虾血细胞产生大量ROS,导致氧化胁迫毒害。抗氧化酶是机体内清除过多ROS、保护机体免受氧化损伤的重要工具。一些研究发现,在环境胁迫过程中,虾会调节抗氧化酶基因的表达水平,使其在抗胁迫中发挥作用(Wangetal.,2009;Guoetal.,2013)。本研究分析5种抗氧化相关基因对硫化物胁迫的分子响应,探讨这些基因在抗硫化物胁迫中的作用。
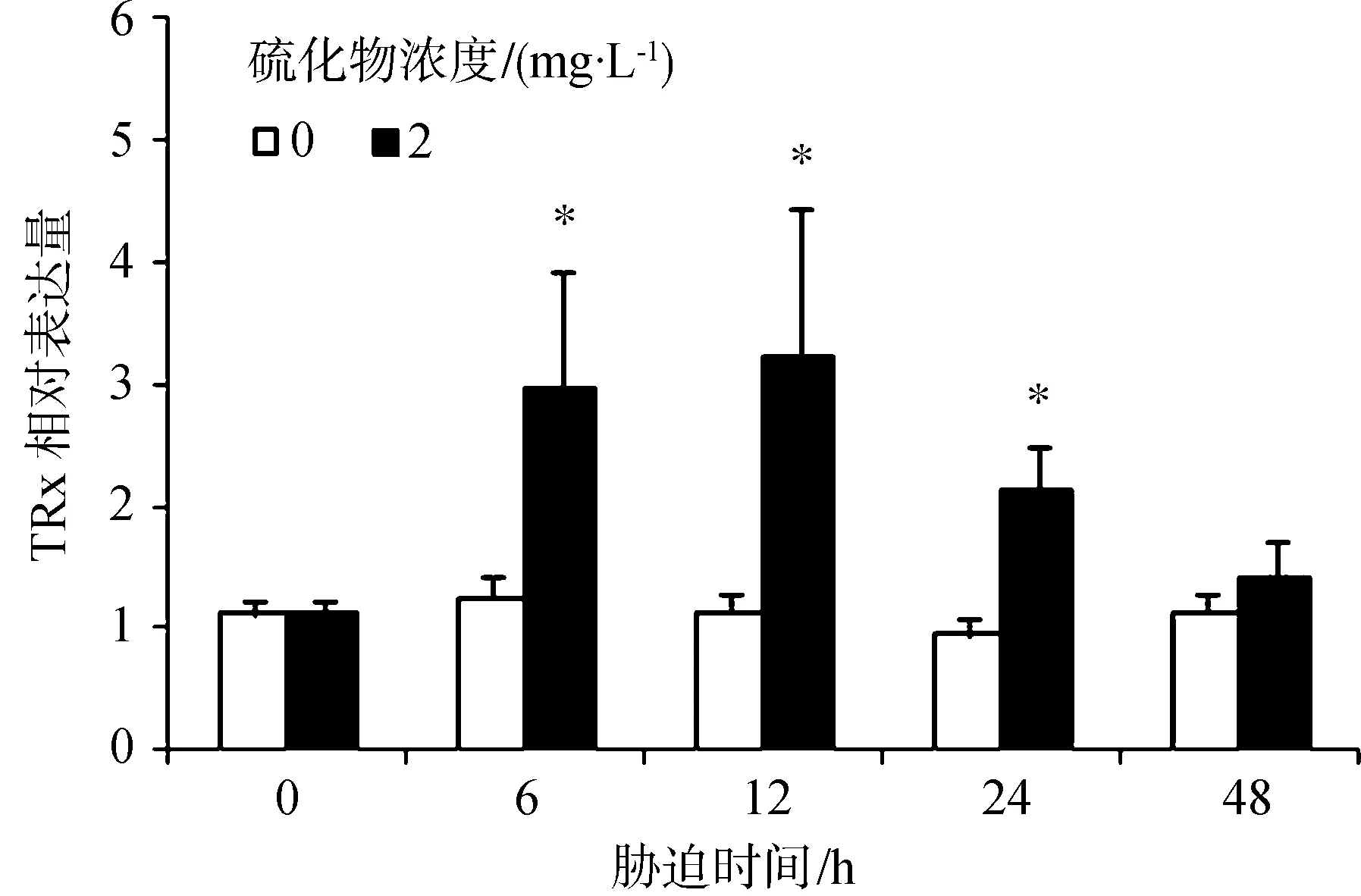
图5 凡纳滨对虾经硫化物胁迫后的血细胞中TRx基因表达量
SOD是抗氧化酶系统中清除氧自由基的第一个酶,它可将超氧阴离子(O2-)转化为过氧化氢(H2O2)。研究表明,甲壳动物的SOD活力会受到硫化物胁迫的影响。日本沼虾Macrobachiumnipponense经1.2 mg·L-1和4.0 mg·L-1硫化物胁迫48 h后,肌肉的SOD活力显著上升(管越强等,2011)。低浓度硫化物胁迫在短期内可使河蟹Eriocheirsinensis血淋巴和鳃组织中的SOD活力提高,胁迫时间延长至10 d后,血淋巴中SOD活力逐渐下降;而高浓度硫化物胁迫则抑制SOD活力(顾顺樟,2007)。在对日本囊对虾Marsupenaeusjaponicus和凡纳滨对虾的研究中也发现,经硫化物胁迫后,血细胞中SOD活力显著上升(Chengetal.,2007;Hsu & Chen,2007)。根据所含金属辅基的不同,在真核生物中SOD可分为Cu,Zn-SOD和Mn-SOD 2种。本研究分别测定了这2种SOD的表达水平的变化,结果显示,2种SOD的基因转录水平均不同程度地被诱导上调。就敏感度而言,2种SOD均在胁迫6 h时被显著诱导上调,胁迫组Cu,Zn-SOD表达水平是对照组的1.80倍,其最大值出现在12 h时,而Mn-SOD在6 h时已达到最大值;就持续性而言,胁迫组Cu,Zn-SOD的表达水平在48 h时恢复至对照组水平,而Mn-SOD在胁迫的48 h内均显著高于对照组;就相对表达量而言,胁迫组Cu,Zn-SOD表达水平达到最大值时是对照组的8.16倍,而胁迫组Mn-SOD达到最大值时是对照组的14.73倍。由此可见,2种SOD均对硫化物胁迫敏感,在胁迫6 h时均被诱导,而Mn-SOD的表达量上调的持续性更强,并且相对表达量更高。一般认为Cu,Zn-SOD主要存在于胞浆中,而Mn-SOD存在于线粒体中,本研究结果显示在硫化物胁迫作用下,2种SOD的表达量均被诱导上调,表明2种SOD可能在血细胞的不同部位发挥抗氧化作用。
CAT是清除H2O2的特异性酶,可以将H2O2分解为H2O和O2。GPx特异性地催化还原型谷胱甘肽对H2O2和氢过氧化物的还原反应,使有毒的过氧化物还原成无毒的羟基化合物,同时促进H2O2的分解,从而保护细胞结构和功能免受过氧化物的损害。因此CAT和GPx在抗氧化防御过程中具有一定的协同作用(Guoetal.,2013)。本研究结果显示,GPx表达水平在胁迫6 h时显著上升,在24 h时略有下调,但仍显著高于对照组;而CAT表达水平在胁迫12 h时才显著被诱导,在胁迫的24 h时表达量达到最大值,为对照组的15.30倍,这些结果表明GPx对硫化物胁迫更敏感,在胁迫的前期发挥作用,而CAT主要在胁迫后期发挥作用。
TRx是细胞内具有氧化还原活性的小分子蛋白质,是蛋白质二硫键的还原酶,参与多种生理过程,其中包括调节抗氧化作用、抗胁迫作用(丛艺等,2008)。结果显示,在硫化物胁迫作用下,TRx表达量在6~24 h时有显著上调,在48 h时恢复至对照组水平,表明TRx对硫化物胁迫敏感,在胁迫前期可能发挥重要的抗胁迫作用。
本研究结果显示,在硫化物胁迫作用下,各类抗氧化相关基因均呈现出不同程度的表达量上调,表明凡纳滨对虾血细胞通过调节胞内抗氧化相关基因的表达水平以进行抗氧化防御。各类抗氧化相关基因表现出对硫化物胁迫的不同敏感度,Cu,Zn-SOD、Mn-SOD、GPx和TRx在胁迫前期发挥作用,CAT主要在胁迫后期发挥作用;各基因在表达量的增量上也有所不同,体现了它们在抗硫化物胁迫过程中具有一定的协同作用。以往一些研究显示,通过合理的营养学手段可显著提高水产动物机体的抗亚硝酸盐、铜离子、氨氮等胁迫的能力(冼健安等,2013;类延菊等,2015;施兆鸿等,2015),合理的营养调控能否有效提高虾的抗硫化物胁迫能力,仍有待深入研究。
丛艺, 蒋圣娟, 张士璀. 2008. 硫氧还蛋白及其相关蛋白的研究进展[J]. 海洋科学, 32(6): 79-84.
顾顺樟. 2007. 硫化物对中华绒螯蟹(Eriocheirsinensis)雌性亲体胁迫效应的研究[D]. 上海: 华东师范大学.
管越强, 裴素蕊, 李泽健. 2011. 急性硫化物胁迫对日本沼虾免疫和抗氧化系统的影响[J]. 水生态学杂志, 32(6): 89-94.
管越强, 王慧春, 李利. 2009. 硫化物胁迫对日本沼虾呼吸代谢和能量代谢酶的影响[J]. 生态环境学报, 18(6): 2017-2022.
郭慧, 冼健安, 王安利. 2015. 亚硝酸盐对凡纳滨对虾血细胞毒性及p53基因表达的影响[J]. 水生态学杂志, 36(2): 61-67.
类延菊, 徐玮, 张彦娇, 等. 2015. 水体铜胁迫条件下饲料中硫辛酸对皱纹盘鲍的保护作用[J]. 中国海洋大学学报, 45(7): 39-45.
李建, 姜令绪, 王文琪, 等. 2007. 氨氮和硫化氢对日本对虾幼体的毒性影响[J]. 上海水产大学学报, 16(1): 22-27.
施兆鸿, 张艳亮, 高权新, 等. 2015. 饲料维生素E水平影响云纹石斑鱼幼鱼对氨氮胁迫的响应[J]. 动物营养学报, 27(5): 1596-1604.
冼健安, 苗玉涛, 叶超霞, 等. 2013. 饲料铜含量对亚硝酸盐胁迫下斑节对虾血细胞的影响[J]. 水生态学杂志, 34(5): 71-75.
杨世平, 阮德雄, 陈兆明, 等. 2014. 不同盐度条件下硫化物对斑节对虾的毒性试验[J]. 安徽农业科学, 42(15): 4673-4675.
於叶兵, 陆伟, 黄金田, 等. 2011. 亚硝酸盐和硫化物对克氏原螯虾幼虾的毒性效应研究[J]. 水生态学杂志, 32(1): 111-114.
余静, 许燕, 王芳. 2011. 硫化氢对罗氏沼虾4种组织中POD、PPO酶活力的影响[J]. 上海师范大学学报(自然科学版), 40(5): 546-550.
Cheng SY, Hsu SW, Chen JC. 2007. Effect of sulfide on the immune response and susceptibility toVibrioalginolyticusin the kuruma shrimpMarsupenaeusjaponicus[J]. Fish & Shellfish Immunology, 22: 16-26.
Guo H, Xian JA, Li B,etal. 2013. Gene expression of apoptosis-related genes, stress protein and antioxidant enzymes in hemocytes of white shrimpLitopenaeusvannameiunder nitrite stress[J]. Comparative Biochemistry and Physiology C, 157: 366-371.
Hsu SW, Chen JC. 2007. The immune response of white shrimpPenaeusvannameiand its susceptibility toVibrioalginolyticusunder sulfide stress[J]. Aquaculture, 271: 61-69.
Mai WJ, Yan JL, Wang L,etal. 2010. Acute acidic exposure induces p53-mediated oxidative stress and DNA damage in tilapia (Oreochromisniloticus) blood cells[J]. Aquatic Toxicology, 100: 271-281.
Wang J, Zhang P, Shen Q,etal. 2013. The effects of cadmium exposure on the oxidative state and cell death in the gill of freshwater crabSinopotamonhenanens[J]. PLoS ONE, 8(5): 1-9.
Wang WN, Zhou J, Wang P,etal. 2009. Oxidative stress, DNA damage and antioxidant enzyme gene expression in the Pacic white shrimp,Litopenaeusvannameiwhen exposed to acute pH stress[J]. Comparative Biochemistry and Physiology C, 150: 428-435.
Effects of Sulfide Stress on Expressions of Antioxidant Enzymes inLitopenaeusvannameiHaemocytes
WANG Lei1, ZHANG Xiuxia2, ZHENG Peihua1, LU Yaopeng1, XIAN Jian’an2*
(1. Institute of Pharmaceutical Research, South China Normal University, Guangzhou 510631, China; 2. Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China)
Sulfide is one of the common water pollutants in aquaculture. To investigate the role of antioxidant enzymes in anti-sulfide defense of shrimp haemocytes, gene expression levels of copper, zinc-superoxide dismutase (Cu, Zn-SOD), manganese-superoxide dismutase (Mn-SOD), catalase (CAT), glutathion peroxidase (GPx) and thioredoxin (TRx) were determined in haemocytes of the white shrimp (Litopenaeusvannamei) exposure to sulfide (0 mg·L-1and 2.0 mg·L-1) for 0 h, 6 h, 12 h, 24 h and 48 h. The results showed that expression levels of Cu, Zn-SOD and TRx significantly increased after 6~24 h exposure, and reduced to the control levels at 48 h. Mn-SOD and GPx mRNA expression levels were up-regulated after 6~48 h exposure. CAT expression level began to increase after 12 h exposure. These results indicated that the mRNA expressions of antioxidant enzymes were induced to protect the haemocytes against sulfide stress. Cu, Zn-SOD, Mn-SOD, GPx and TRx displayed high sensitivity to sulfide stress, and might play the dominant role at the initial stage of sulfide stress, whereas CAT mainly worked at the later period.
sulfide;Litopenaeusvannamei; haemocyte; antioxidant enzyme; gene expression
2016-08-25 接受日期:2016-09-21
国家自然科学基金项目(31500326); 广东省自然科学基金项目(2014A030310185); 广东省普通高校青年创新人才项目(2014KQNCX056); 广东省普通高校特色创新项目(2015KTSCX019); 华南师范大学青年教师科研培育基金项目(13KJ16)
汪蕾(1986—), 女, 讲师, 主要从事水产养殖生态及毒理学研究, E-mail:wanglei@scnu.edu.cn
*通信作者Corresponding author, E-mail:xian-ja@163.com
10.11984/j.issn.1000-7083.20160229
X503.225; Q959.223
A
1000-7083(2016)06-0884-05
