两种不同地理种群松毛虫赤眼蜂对米蛾卵的寄生功能及数值反应
2016-12-09杜文梅臧连生张俊杰祁颖慧阮长春
杜文梅,臧连生,张俊杰,祁颖慧,阮长春
两种不同地理种群松毛虫赤眼蜂对米蛾卵的寄生功能及数值反应
杜文梅1,臧连生1,张俊杰1,祁颖慧2,阮长春1
(1吉林农业大学生物防治研究所/天敌昆虫应用工程技术研究中心,长春130118;2吉林省农业技术推广总站,长春130021)
【目的】松毛虫赤眼蜂()是目前生物防治中应用面积最大的一种寄生性天敌。不同地理种群松毛虫赤眼蜂其生物学特性存在一定差异,论文旨在明确两种不同地理种群松毛虫赤眼蜂对米蛾卵的寄生功能反应及数值反应参数,为不同地理种群赤眼蜂的应用提供参考。【方法】选择松毛虫赤眼蜂黑龙江种群(TD-HLJ)和江苏种群(TD-JS),以米蛾卵为供试寄主,研究两个地理种群松毛虫赤眼蜂对米蛾卵的寄生功能反应与数值反应,并利用种群趋势指数(I)和干扰控制作用指数(IIPC)对两个不同地理种群松毛虫赤眼蜂进行分析比较。【结果】功能反应结果表明,TD-HLJ和TD-JS种群的寄生功能反应符合HollingⅡ型方程,TD-HLJ种群最大寄生量为186.78粒/d,比TD-JS的最大寄生量高75.68粒/d。数值反应研究表明,不同米蛾卵密度对松毛虫赤眼蜂TD-HLJ和TD-JS种群的寄生、发育及繁殖有显著影响,米蛾卵密度为1粒时是TD-HLJ和TD-JS种群羽化的临界值,米蛾卵密度为5和10粒时分别是TD-HLJ和TD-JS种群繁殖的临界值。采用I和IIPC分析松毛虫赤眼蜂的控制作用,TD-HLJ和TD-JS种群的I值均随着米蛾卵密度的增加而增加,对照TD-HLJ和TD-JS种群的I值分别为68.70和49.20,即TD-HLJ种群的下一代数量比TD-JS种群的数量多19.5倍;TD-HLJ和TD-JS种群的IIPC值均随着米蛾卵密度的增加而增加,说明随着卵密度的增加,TD-HLJ和TD-JS种群对寄主卵的控制作用逐渐减小。【结论】综合功能反应和数值反应结果,松毛虫赤眼蜂TD-HLJ种群比TD-JS种群具有更好的生物防治潜能。
松毛虫赤眼蜂;地理种群;米蛾卵;功能反应;数值反应
0 引言
【研究意义】松毛虫赤眼蜂()属赤眼蜂科(Trichogrammatidae),赤眼蜂属(),主要用于防治松毛虫()、亚洲玉米螟()、二化螟()等农林害虫,是目前生物防治上应用面积最大、防治效果最好的一种卵寄生性天敌[1-5]。20世纪60年代以来,中国利用大卵(柞蚕卵)繁育赤眼蜂获得成功[6],加快了松毛虫赤眼蜂的应用推广,目前仅在东北地区,每年利用松毛虫赤眼蜂防治玉米螟面积就达到400万hm2以上[1]。【前人研究进展】选择优良蜂种是获得高质量赤眼蜂的前提,也是提高防治效果的关键。许多研究学者指出,不仅要选择一个合适的种,而且要选择一个合适的种下类型[7-8]。张帆等[9]在半自然条件下研究了不同赤眼蜂品系对亚洲玉米螟卵的寄生,发现赤眼蜂不同品系对玉米螟卵的寄生能力存在一定差异;李莹等[10]研究了不同地理种群松毛虫赤眼蜂寄生潜能,发现雌蜂寿命、有效产卵量、羽化率等指标存在显著性差异,因而,筛选最适宜的蜂种或品系对利用赤眼蜂防治尤为重要。在寄生天敌-寄主复杂关系中,研究者先后提出了功能反应、数值反应及空间异质性反应来评价一种天敌昆虫的控害潜能。功能反应是研究一个寄生者在单位时间内对给定不同密度寄主的寄生数量的变化,旨在揭示寄生者的最大寄生潜能;数值反应则是指猎物或者寄主密度的变化对寄生者或者捕食者生长发育、繁殖的影响,旨在揭示当猎物密度低至何种程度时会对寄生者的生存、发育、繁殖产生不良影响及其影响的程度[11-14]。害虫种群数量控制要求把种群数量长期控制在引起经济损失的容许水平以下,消除危害的威胁。庞雄飞等[15]提出了种群干扰作用控制指数(interference of index of population control,IIPC),主要研究各类因子对种群趋势指数的影响。【本研究切入点】目前未见不同地理种群松毛虫赤眼蜂对米蛾卵的功能反应及数值反应相关研究报道。【拟解决的关键问题】明确两种不同地理种群(黑龙江种群和江苏种群)松毛虫赤眼蜂在不同米蛾()卵密度下的寄生、生长发育及繁殖情况,为同种但不同种群赤眼蜂的应用提供参考。
1 材料与方法
试验于2015年吉林农业大学生物防治研究所完成。
1.1 供试昆虫
松毛虫赤眼蜂黑龙江种群(TD-HLJ):2014年7月采自黑龙江省尚志市玉米田,原寄主为亚洲玉米螟卵块;松毛虫赤眼蜂江苏种群(TD-JS):2014年12月采自江苏省杨树林,原寄主为杨扇舟蛾卵()。两个种群采集后在实验室内进行单蜂纯化及解剖雄性外生殖器,由华南农业大学黄寿山教授鉴定。两个不同地理种群的赤眼蜂在吉林农业大学生物防治研究所实验室内温度(25±1)℃;RH(75±5)%;光周期L﹕D=14 h﹕10 h条件下,用米蛾卵续代繁殖10代后用于试验。米蛾在室内用玉米面饲养,试验前取当日新产的米蛾卵制成卵卡,用30 W的紫外灯照射30 min,杀死胚胎,供松毛虫赤眼蜂寄生。
1.2 功能反应
分别取羽化6 h以内的两种不同地理种群的松毛虫赤眼蜂雌蜂,单头引入试管中(10 mm×100 mm)饲喂20%的蜂蜜水,米蛾卵密度设置为20、30、50、60、100、120、150、200粒,每个处理重复20次。温度(25±1)℃;RH(75±5)%;光周期L﹕D=14 h﹕10 h条件下寄生24 h后引出赤眼蜂,5 d后统计寄生卵粒数。
1.3 数值反应
参考文献[16-17]数值反应的方法,分别取羽化6 h以内的两种不同地理种群的松毛虫赤眼蜂雌蜂,单头引入试管中(10 mm×100 mm)饲喂20%的蜂蜜水,米蛾卵密度设置为1、5、10、15、20、25、30、50、70、100粒,对照设足量米蛾卵(150粒),每个处理重复20次。24 h后取出米蛾卵,发育5 d后检查寄生卵粒数,出蜂后调查羽化数、雌雄数、畸形数;出蜂后将单头雌蜂引入试管中(10 mm×100 mm)饲喂20%的蜂蜜水,提供过量的米蛾卵,每24 h更换1次米蛾卵,直到雌蜂死亡,发育5 d后检查寄生卵粒数,重复20次。
1.4 数据处理
数据用DPS14.10软件辅助进行分析。功能反应采用Holling Ⅱ型进行拟合,其方程式为:Na=aTN /(1+aThN)。式中,Na为被寄生的寄主数量;N为寄主密度;T为发现寄主的时间(试验总用时为1 d);a为瞬间攻击率;Th为处置时间(d)。
种群趋势指数(I)根据文献[15,18-19]的方法组建不同米蛾卵密度下赤眼蜂发育繁殖的生命表,分析两种不同地理种群松毛虫赤眼蜂的种群趋势指数。I=N1/N0,式中,N1、N0下一代、当代的种群数量。
干扰作用控制指数(IIPC):种群干扰作用控制指数主要研究各类因子对种群趋势指数的影响。IIPC= I' /I ,I'为每个处理的种群趋势指数,I为对照的种群趋势指数。
2 结果
2.1 赤眼蜂对米蛾卵寄生功能反应
松毛虫赤眼蜂TD-HLJ和TD-JS种群的寄生功能反应拟合方程与圆盘方程模型很好符合,2值均>0.95,达到显著水平。两种不同地理种群松毛虫赤眼蜂瞬间攻击率a(TD-JS)>a(TD-HLJ);TD-JS的处置时间Th比TD-HLJ多0.00365 d;由模拟方程算出的日最大寄生米蛾卵数Namax可以看出TD-HLJ最大寄生量为186.78粒,比TD-JS的最大寄生量高75.68粒(表1)。

表1 不同地理种群松毛虫赤眼蜂对米蛾卵的功能反应
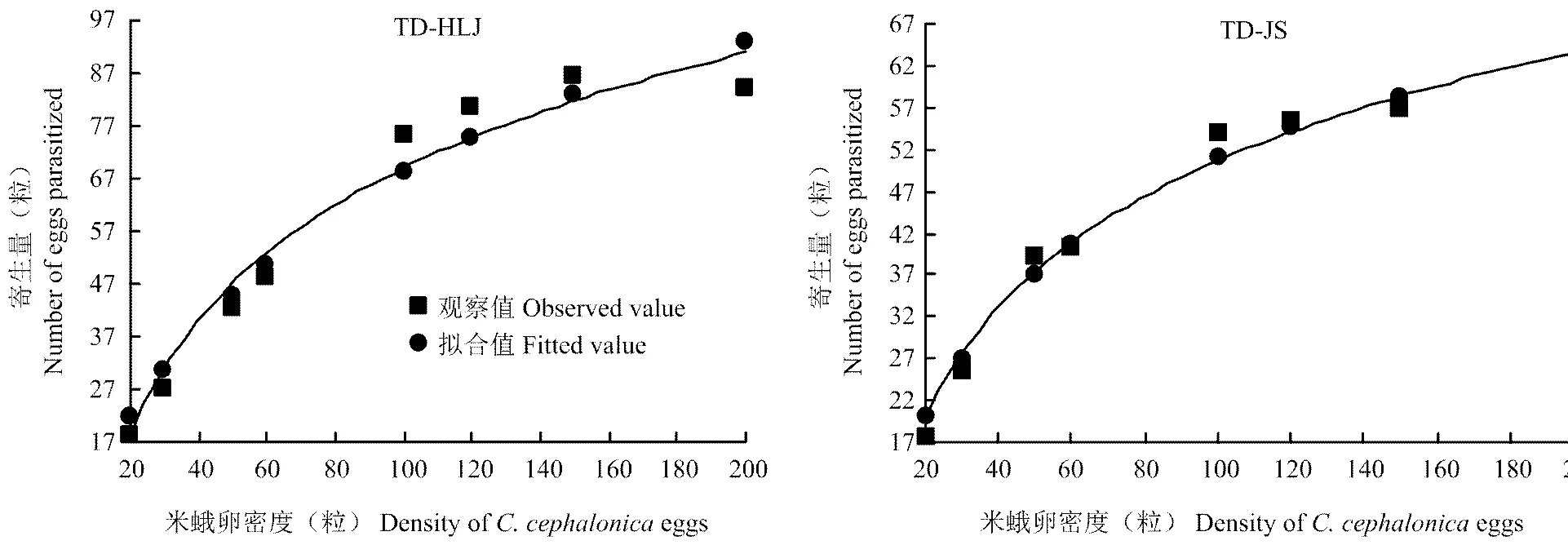
图1 松毛虫赤眼蜂对米蛾卵寄生功能反应
图1为松毛虫赤眼蜂TD-HLJ和TD-JS种群寄生米蛾卵的寄生功能反应曲线,由图可知,两个种群松毛虫赤眼蜂的寄生量均随着猎物密度的增加而增加,当猎物密度增加到一定量后,寄生量增加速度变慢,趋于稳定。
2.2 赤眼蜂对米蛾卵寄生数值效应
松毛虫赤眼蜂TD-HLJ种群在各米蛾卵密度下均可交配产卵,母代寄生米蛾卵数随着米蛾卵密度的增加呈显著上升趋势(=79.45,<0.0001),米蛾卵密度为1粒时,16个寄生卵中仅有1粒卵正常羽化出1头雌蜂和1头雄蜂,因此,该密度为TD-HLJ种群的羽化临界值。随着米蛾卵密度不同时,寄生率、羽化率、雌雄蜂比均显著性差异(寄生率:=2.54,=0.0066;羽化率:=3.04,=0.0013;♀﹕♂:=7.90,<0.0001)。当米蛾卵密度为5粒时,86.00%的寄主卵被寄生,子代♀﹕♂=1.38,子代平均产卵量为76.03,随着米蛾卵密度的增加,羽化率、♀﹕♂值、子代雌蜂寿命、子代产卵量等指标逐渐接近对照,由此可说明人工繁殖松毛虫赤眼蜂TD-HLJ种群时,米蛾卵密度5粒是其维持种群繁殖的临界值(表2)。
松毛虫赤眼蜂TD-JS种群随着米蛾卵密度的增加寄生数、寄生率、羽化率及性指标处理间均存在显著差异(寄生数:=53.92,<0.0001;寄生率:=9.19,<0.0001;羽化率:=10.47,<0.0001;♀﹕♂:=11.90,<0.0001)。当米蛾卵密度为1粒时,TD-JS种群平均寄生米蛾卵数只有0.2粒,羽化率只有20%,且均为雄虫,因此,米蛾卵密度为1粒是TD-JS种群羽化的临界值;米蛾卵密度为5粒时,寄生数仅为1.95粒,显著低于对照,羽化率仅为55.00%,且♀﹕♂=0.70,表现出子代偏雄,不利于续代繁殖,而当米蛾卵密度大于10粒时,随着米蛾卵密度的增加,各指标逐渐升高接近对照,故人工繁殖松毛虫赤眼蜂TD-JS种群时,米蛾卵10粒为维持该种群繁殖的临界值(表3)。
米蛾卵密度对松毛虫赤眼蜂TD-HLJ和TD-JS种群的寄生、发育及繁殖有一定的影响,以米蛾卵密度1粒为例计算TD-HLJ的群的种群趋势指数(I)和干扰作用控制指数(IIPC),I'(0.6)=12/20,对照I(68.7)=1374/20,IIPC(0.0087)=0.6/68.7,TD-HLJ和TD-JS种群的种群趋势指数(I)随着米蛾卵梯度的增加而增加,且TD-HLJ大于TD-JS;对照松毛虫赤眼蜂TD-HLJ和TD-JS的种群趋势指数(I)分别为68.70和49.20,即TD-HLJ种群的下一代数量比TD-JS种群的数量多19.5倍(表4)。

表2 不同米蛾卵密度下的松毛虫赤眼蜂(TD-HLJ)寄生发育及繁殖参数
表中数据为平均值,不同小写字母表示差异显著(<0.05)。下同
Data in the table were average. Different lowercases indicated significant differences (<0.05). The same as below

表3 不同米蛾卵密度下的松毛虫赤眼蜂(TD-JS)寄生发育及繁殖参数

表4 松毛虫赤眼蜂两个地理种群的数值反应参数
以对照TD-HLJ种群为例说明表中的参数,当米蛾卵密度为150粒时,寄生数表示20个重复中有18头赤眼蜂寄生了米蛾卵,羽化出蜂数表示子代共计羽化出赤眼蜂1374头
Here the control population of TD-HLJ was used as an example to illustrate the parameters in the table. When 150 eggs ofwere offered, for parasitism number, 18 individuals offrom 20 replicates parasitized the eggs of rice moth. For number of emerging wasps, total of 1374offsprings emerge
3 讨论
天敌的数值反应和功能反应是评价天敌对害虫的控制能力的基础研究[20-22], Moezipour等[21]研究了甘蓝夜蛾赤眼蜂()对麦蛾()的寄生功能反应,在25℃时,其寄生功能反应为HollingⅡ型;陈鹏等[23]研究了榆紫叶甲赤眼蜂()对榆紫叶甲(i)卵的寄生功能反应,在14—30℃时,其寄生功能反应均为HollingⅡ型;冯宏祖等[24]研究了两种不同地理种群松毛虫赤眼蜂(吉林种群和新疆抗旱种群)对苹果蠹蛾()卵的寄生功能反应均符合HollingⅡ型,且吉林种群和新疆抗旱种群对苹果蠹蛾卵的日最大寄生量分别为21.20和23.24粒。本研究结果表明,松毛虫赤眼蜂TD-HLJ和TD-JS种群对米蛾卵的寄生功能反应也为HollingⅡ型,与文献报道一致。松毛虫赤眼蜂TD-HLJ和TD-JS种群对米蛾卵的最大日寄生量分别为186.78和111.10粒,对比冯宏祖等[24]的研究结果,说明松毛虫赤眼蜂对米蛾卵的寄生能力明显强于苹果蠹蛾卵,且地理种群不同时,对寄主寄生能力也存在差异。而根据报道,在实际应用赤眼蜂防治害虫时,种下类型的选择会直接影响其防治效果[5,25-27]。
通过研究两种地理种群松毛虫赤眼蜂对米蛾卵的数值反应,结果表明米蛾卵1粒是松毛虫赤眼蜂TD-HLJ和TD-JS种群羽化临界值,米蛾卵密度5粒和10粒分别为松毛虫赤眼蜂TD-HLJ和TD-JS种群繁殖的临界值,说明米蛾卵密度过低会影响松毛虫赤眼蜂正常繁殖。谢丽娜等[22]在研究松毛虫赤眼蜂孤雌生殖品系和两性生殖品系对米蛾卵的功能反应时提出米蛾卵密度过低会抑制寄生效率,这与本文结论相一致。因此,在释放赤眼蜂防治害虫时,应将害虫种群密度考虑在内,在害虫密度过低时,应考虑适当补充替代寄主卵,以便天敌顺利在田间定殖。
本研究通过功能反应和数值反应客观地评价了两个不同地理种群的松毛虫赤眼蜂其寄生潜能存在差异性,李莹等[10]研究发现5个不同地理种群的松毛虫赤眼蜂其寄生潜能、生命表参数也存在差异;冯建国等也曾提出松毛虫赤眼蜂不同种型其寄生选择及寄生率均存在差异[28-29]。松毛虫赤眼蜂在中国分布广泛,寄主种类多,目前被普遍用于防治玉米螟,笔者在进行赤眼蜂保存时发现TD-JS种群并不能寄生玉米螟卵(数据未发表),这也许是由于两个不同地理种群的松毛虫赤眼蜂其生态环境、寄主等不同所引发的,因而在应用TD-HLJ和TD-JS种群防治害虫时,应综合考虑蜂种的寄生潜能、蜂种采集地及采集寄主。
4 结论
对两种不同地理种群的松毛虫赤眼蜂在其米蛾卵上的寄生功能反应及数值反应参数的比较表明,松毛虫赤眼蜂TD-HLJ种群寄生能力较强,其主要寄生功能反应及数值反应参数(寄生上限、寄生数、子代产卵量、种群趋势指数(I)和干扰作用控制指数(IIPC))值均优于种群TD-JS。综合功能反应和数值反应结果,作为寄生性天敌松毛虫赤眼蜂TD-HLJ种群比TD-JS具有更好的控制作用。
致谢:华南农业大学黄寿山教授为赤眼蜂蜂种鉴定和试验方案设计提供宝贵意见,在此表示衷心感谢!
References
[1] ZHANG J J, REN B Z , YUAN X H, ZANG L S, RUAN C C, SUN G Z, SHAO X W.Effects of host-egg ages on host selection and suitability of four Chinesespecies, egg parasitoids of the rice striped stem borer,., 2014, 59(2): 159-166.
[2] Smith S M. Biological control with: advances, success, and potential of their use., 1996, 41(1): 375-406.
[3] Pizzol J, Desneux N, Wajnberg E, Thiery D. Parasitoid and host egg ages have independent impact on various biological traits in aspecies., 2012, 85: 489-496.
[4] 刘志诚, 刘建峰, 张帆. 赤眼蜂繁殖及田间应用技术. 北京: 金盾出版社, 2000: 93-119.
Liu Z C, Liu J F, Zhang F.spp. Beijing: Jindun Publishing House, 2000: 93-119. (in Chinese)
[5] 黄寿山, 臧连生, 阮长春. 赤眼蜂寄生生态学与工厂化繁殖应用. 北京: 科学出版社, 2013: 147-150.
Huang S S, Zang L S, Ruan C C.. Beijing: Science Press, 2013: 147-150. (in Chinese)
[6] 林莲欣, 冯建国. 山东济南地区玉米螟卵赤眼蜂(sp.)利用初步研究//中国昆虫学会1962年学术讨论会刊, 1962: 253-255.
Lin L X, Feng J G.egg using preliminary study of Ji’nan area, Shandong//:, 1962: 253-255. (in Chinese)
[7] Wang Z Y, He K L, Zhang F, Lu X, Babendreier D. Mass rearing and release offor biological control of insect pests of corn in China., 2014, 68: 136-144.
[8] 忻介六. 天敌昆虫的品质管理问题. 昆虫天敌, 1982, 4(3): 56-60.
Xin J L. Quality management of natural enemies insects.,1982, 4(3): 56-60. (in Chinese)
[9] 张帆, 王素琴, 张君明, 孟昭君, 李得刚. 半自然条件下几种赤眼蜂及品系对亚洲玉米螟卵寄生能力比较. 植物保护, 2004, 30(4): 29-32.
Zhang F, Wang S Q, Zhang J M, Meng Z J, Li D G. Parasitism of differentspecies and strains on eggs of Asian corn borer,under semi-field condition., 2004, 30(4): 29-32. (in Chinese)
[10] 李莹, 董辉, 丛斌, 钱海涛, 杨海霞, 张柱婷, 孙巍. 温度对松毛虫赤眼蜂不同地理种群寄生潜能及种群参数的影响. 中国生物防治学报, 2013, 29(1): 11-16.
Li Y, Dong H, Cong B, Qian H T, Yang H X, Zhang Z T, Sun W. Effect of temperature on reproductive potential and population parameters of differentgeographic populations., 2013, 29(1): 11-16. (in Chinese)
[11] 张艳璇, 林坚贞, 季洁, 康玉妹, 陈霞. 数值反应和实验种群生命表分析胡瓜钝绥蟎对柑橘全爪螨的控制能力. 中国农业科学, 2004, 37(12) : 1866-1873.
Zhang Y X, Lin J Z, Ji J, Kang Y M, Chen X. Analysis of numerical responses and main life parameters for determining the suppression ofon., 2004, 37(12): 1866-1873. (in Chinese)
[12] 丁岩钦. 天敌-害虫作用系统中的数学模型及其主要参数的估计. 昆虫知识, 1983, 20(4): 120-124.
Ding Y Q. Mathematical model of the predators-insects system and its main parameters estimation., 1983, 20(4): 120-124. (in Chinese)
[13] Montserrat M, Albajes R, Castañé C. Functional response of four Heteropteran predators preying on greenhouse whitefly (Homoptera: Aleyrodidae) and western flower thrips (Thysanoptera: Thripidae)., 2000, 29(5): 1075-1082.
[14] 深谷昌次, 谷圭治. 综合防治. 忻介六, 译. 上海: 上海科学技术出版社, 1980: 97.
Masatsugu F, Keiji K.. Xin J L, Trans. Shanghai: Shanghai Science and Technology Press, 1980: 97. (in Chinese)
[15] 庞雄飞, 梁广文. 害虫种群系统的控制. 广州: 广东科技出版社, 1995: 24-26.
Pang X F, Liang G W.. Guangzhou: Guangdong science and Technology Press, 1995: 24-26. (in Chinese)
[16] 黄林茂, 黄寿山. 黑肩绿盲蝽捕食褐飞虱卵的功能与数值反应. 生态学报, 2010, 30(15): 4187-4195.
Huang L M, Huang S S. Functional and numerical responses of the predacious plant bug on the rice brown planthopper egg., 2010, 30(15): 4187-4195. (in Chinese)
[17] 李水泉, 黄寿山, 韩诗畴, 李志刚, 叶静文, 徐英杰. 黄玛草蛉捕食米蛾卵的功能反应与数值反应. 生态学报, 2012, 32(21): 6842-6847.
Li S Q, Huang S S, Han S C, Li Z G, Ye J W, Xu Y J. Functional and numerical responses offeeding oneggs., 2012, 32(21): 6842-6847. (in chinese)
[18] 庞雄飞, 梁广文, 尤民生. 种群生命系统研究方法概述. 昆虫天敌, 1986, 8(3): 176-186.
Pang X F, Liang G W, You M S. On the methodology of the insect population life systems., 1986, 8(3): 176-186. (in Chinese)
[19] 庞雄飞, 梁广文, 尤民生, 吴伟坚. 昆虫种群生命系统研究的状态方程. 华南农业大学学报, 1988, 9(2): 1-10.
Pang X F, Liang G W, You M S, Wu W J. State-space approach to population life systems., 1988, 9(2): 1-10. (in Chinese)
[20] BAYLISS P, CHOQUENOT D. The numerical response: rate of increase and food limitation in herbivores and predators., 2002, 357(1425): 1233-1248.
[21] MOEZIPOUR M, KAFIL M, Allahyari H. Functional response ofat different temperatures and relative humidities., 2008, 61(2): 245-250.
[22] 谢丽娜, 董辉, 钱海涛, 闫京京, 丛斌. 不同温度下松毛虫赤眼蜂孤雌产雌品系和两性生殖品系对米蛾卵的寄生功能反应. 昆虫学报, 2013, 56(3): 263-269.
XIE L N, DONG H, QIAN H T, YAN J J, CONG B. Functional response of thelytokous and arrhenotokous strains of(Hymenoptera: Trichogrammatidae) to eggs of(Lepidoptera: Pyralidae) at different temperatures., 2013, 56(3): 263-269. (in Chinese)
[23] 陈鹏, 王凤珍, 李春成, 王秀梅, 阮长春. 榆紫叶甲赤眼蜂寄生功能反应对梯度恒温的响应. 东北林业大学学报, 2015, 43(1): 114-116.
CHEN P, WANG F Z, LI C C, WANG X M, RUAN C C. Effect of temperature on function response of., 2015, 43(1): 114-116. (in Chinese)
[24] 冯宏祖, 王兰, 郭文超, 聂蒙, 张红涛, 许建军. 赤眼蜂对苹果蠹蛾卵功能反应的研究. 中国生物防治学报, 2013, 29(2): 307-311.
Feng H Z, Wang L, Guo W Q, NIE M, ZHANG H T, XU J J. Functional response ofon eggs of., 2013, 29(2): 307-311. (in Chinese)
[25] 张芝利, 黄融生, 朱庸, 王素琴, 康总江, 潘永成, 尹英华, 张文山, 恽秀芹, 孙爱华. 利用玉米螟赤眼蜂防治玉米螟的研究初报. 昆虫知识, 1979(5): 207-210.
Zhang Z L, Huang R S, Zhu Y, Wang S Q, KANG Z J, PAN Y C, YIN Y H, ZHANG W S, YUN X Q, SUN A H. Study oncontrolling corn borer preliminary utilization., 1979(5): 207-210. (in Chinese)
[26] 沈伯钧, 邱鸿贵, 邱中良, 符文俊. 松毛虫赤眼蜂对不同植物的行为反应. 昆虫知识, 1991, 28(6): 359-361.
Shen B Y, Qiu H G, QIU Z L, FU W J.on different plant response behavior., 1991, 28(6): 359-361. (in Chinese)
[27] 张光美, 刘树生, 杨坚伟, 张帆, 孙彤. 影响松毛虫赤眼蜂寄生亚洲玉米螟的因子观察.植物保护学报, 1995, 22(3): 205-210.
Zhang G M, Liu S S, Yang J W, Zhang F, Sun T. On the restrictive factors of parasitism ofon the eggs of Asian corn borer., 1995, 22(3): 205-210. (in chinese)
[28] 冯建国, 董建棠. 利用赤眼蜂防治玉米螟等害虫试验简报. 昆虫知识, 1975(1): 20-21.
Feng J G, Dong J T.The test ofto control corn borer pests briefing etc., 1975(1): 20-21. (in chinese)
[29] 施祖华, 刘树生, 何立新, 何俊华. 松毛虫赤眼蜂种群生物学特性的比较研究VI. 寄主选择和寄主的适合性. 生物防治通报, 1994, 10(1): 1-6.
Shi Z H, Liu S S, He L X, He J H. Comparative studies on the biological characteristics of geographic hose populations ofin china Ⅳ: host selection and host suitability., 1994, 10(1): 1-6. (in chinese)
(责任编辑 岳梅)
Functional Response and Numerical Response of Two Geographic Populations ofto the Eggs of Rice Moth,
DU Wen-mei1, ZANG Lian-sheng1, ZHANG Jun-jie1, QI Ying-hui2, RUAN Chang-chun1
(1Institute of Biological Control, Jilin Agricultural University/Engineering Research Center of Natural Enemy Insects, Changchun 130118;2Jilin Provincial Agro-Tech Extension Center, Changchun 130021)
【Objective】is a widely used parasitoid in biological control.There are some differences in different geographic populations ofand its biological characteristics. This study aimed at understanding the parasitic functional response and numerical response of two typical geographic populations to eggs of the rice moth,so that provided a reference for the application of different geographical populations of the parasitic wasp species.【Method】eggs were used as the host, and the functional response and numerical response of two geographic populations offrom Heilongjiang and Jiangsu parasitizing were investigated. The species flocks of trend index (I) and index of population control (IIPC) of the two different geographical populations were compared and analyzed.【Result】The parasitic functional responses of Heilongjiang population (TD-HLJ) and Jiangsu population (TD-JS) offitted in with HollingⅡ equation. The daily maximum parasitic number of TD-HLJ was 186.78 eggs, more 75.68 eggs than that of TD-JS. The density of 1 grain ofegg was the critical value of TD-HLJ and TD-JS populations of eclosion. The density ofeggs had significant impact on parasitism, development and reproduction of both populations. 5 and 10 eggs ofwere the critical values of TD-HLJ and TD-JS population reproduction. The population trend index (I) of both populations increased with the increase of host egg density. Generally, both of populations had the values of I under the control density with 68.70 for TD-HLJ and 49.20 for TD-JS, which indicated that the number of offsprings of TD-HLJ was 19.5 times of that of TD-JS. Similarly, interference of index of population control (IIPC) of both populations increased with the increase of host egg density, indicating that the biological control effectiveness ofagainst hosts decreased with egg density increasing.【Conclusion】Based on the comprehensive analysis on the results of functional response and numerical response, TD-HLJ exhibits better biological control potential than TD-JS in the insect pest management.
; geographic populations;egg; functionalresponse; numerical response
2016-05-04;接受日期:2016-07-06
国家自然科学基金(31572058)、国家重点基础研究发展计划(2013CB127605)、吉林农业大学青年启动基金(201232)
杜文梅,E-mail:280114191@qq.com。通信作者臧连生,Tel:0431-84533236;E-mail:lsz0415@163.com。通信作者阮长春,E-mail: bio-control@126.com
