干旱胁迫下杜鹃叶片生理与PSII活性变化
2016-12-08周媛董艳芳童俊毛静徐冬云
周媛++董艳芳++童俊++毛静++徐冬云++何朝斌


摘要:以6个杜鹃(Rhododendron L.)品种胭脂蜜[Rhododendron obtusum (Lindl.) Planch. cv. Yanzhimi]、鸳鸯锦(R. obtusum cv. Yuanyangjin)、花蝴蝶(R. hybridum Hort. cv. Variegated Butterfly)、紫宸殿[R. indicum (L.) Sweet. cv. Zichendian]、锦袍(R. hybridum cv. Jinpao)、紫鹤(R. championiae Hook. cv. Zihe)为研究对象,比较了干旱胁迫下不同杜鹃品种的外观形态、生理指标和叶绿素荧光参数的变化。结果表明,随着干旱胁迫时间的延长,所有杜鹃品种叶片出现了不同程度的下垂、萎蔫甚至死亡的情况,其中锦袍的受伤害程度相对最轻。MDA含量呈持续上升或先升后降的趋势;可溶性蛋白含量变化较平缓,总体呈下降的趋势,其中紫鹤在干旱第10天时含量达到最高;叶绿素含量呈先下降后上升的变化趋势;初始荧光(F0)总体呈现下降再平缓上升的状态,其中花蝴蝶与紫鹤在10 d后急剧下降直至消失,这与外观形态表现一致;最大光化学效率(Fv/Fm)与PSII潜在活性(Fv/F0)变化规律一致,除2个干旱敏感品种花蝴蝶和紫鹤急剧降低以外,其他品种均保持平稳状态。
关键词:杜鹃(Rhododendron L.);干旱胁迫;抗旱性;生理响应;叶绿素荧光参数
中图分类号:S685.21+3 文献标识码:A 文章编号:0439-8114(2016)18-4730-05
DOI:10.14088/j.cnki.issn0439-8114.2016.18.026
杜鹃是杜鹃花科(Ericaceae)杜鹃花属(Rhododendron L.)木本植物的总称,别名映山红、满山红等,其种类繁多、花色艳丽、姿态优美,为世界上著名的观赏花卉,对世界园林植物有着重大影响;也是中国传统十大名花之一,有“花中西施”的美誉[1,2]。干旱是影响园林植物正常生长并导致其观赏性下降的重要因素,不同的杜鹃(种、品种)其耐旱性具有显著的不同;李娟[3]对西鹃(R. hybridum Hort.)和毛鹃(R. pulchrum Sweet.)进行了干旱处理,并对西鹃和毛鹃的叶片含水量、叶片相对含水量和叶绿素、脯氨酸、蛋白质含量以及过氧化物酶、过氧化氢酶、抗坏血酸过氧化物酶活性等生理特性做了比较,得出西鹃抗旱性大于毛鹃的结论。黄承玲等[4]对持续干旱条件下高山类型杜鹃露珠杜鹃(R. irroratum Franch.)、迷人杜鹃(R. agastum Balf. f. et W. W. Smith)、大白杜鹃(R. decorum Franch.)的保护酶活性、渗透调节物质、丙二醛(MDA)含量和质膜相对透性进行了测定,并进行抗旱评价,得出丙二醛含量和保护酶活性可以作为高山类型杜鹃抗旱性评价的主要指标结论。叶绿素荧光技术是研究植物光合作用的机制和探测光合生理状况的一种新兴技术,能够快速灵敏、无损伤地反映光合系统Ⅱ(Photosynthetic system Ⅱ,PSⅡ)对光能的吸收、传递、耗散、分配等方面的动态,被认为是研究植物光合能力及对环境胁迫响应的有效手段[5-7]。在杜鹃抗旱性研究中,关于生理指标的研究相对较多,但是将生理指标与叶绿素荧光参数相结合来比较杜鹃抗旱性的报道相对较少。为了更全面地了解杜鹃在干旱胁迫条件下的生理响应机制,课题组对前期筛选出的观赏价值高的5个杜鹃品种与目前园林中栽培应用较多的1个杜鹃品种进行了干旱胁迫条件下的外观形态、生理指标和叶绿素荧光参数测定,以期综合反映杜鹃对干旱逆境的抗性,从而为杜鹃的抗旱育种提供理论依据。
1 材料与方法
1.1 材料
试验于2014年6~10月在武汉市农业科学院林业果树科学研究所内进行。供试材料为6个杜鹃品种,分别为胭脂蜜[R.obtusum(Lindl.) Planch. cv. Yanzhimi]、鸳鸯锦(R. obtusum cv. Yuanyangjin)、花蝴蝶(R. hybridum cv. Variegated Butterfly)、紫宸殿[R. indicum(L.)Sweet. cv. Zichendian]、锦袍(R. hybridum cv. Jinpao)、紫鹤(R. championiae Hook. cv. Zihe)。前5个品种为前期筛选出的观赏价值高的品种,紫鹤为园林绿化市场上常见栽培种。所有参试品种均为大棚内常规管理的扦插盆苗,选取二年生生长一致、健壮的盆苗参加试验。
1.2 取样及方法
在温室内采用盆栽控水法实施水分胁迫处理,并保持与室外的通风。干旱处理为先给盆苗浇清水至饱和后不再浇水,每隔5 d在上午8:00-9:00采叶片样,采样后立即剪碎,混合均匀后对样品进行生理生化指标(MDA、可溶性蛋白、叶绿素含量)检测;试验期间观察记录植株叶片形态变化,以叶片伤害率表示叶片萎蔫程度,直至供试植株叶片严重萎蔫为止。叶片伤害率计算公式:
叶片伤害率=(单株叶片萎蔫数/单株叶片数)×100%。
在测定生理指标的同时进行叶绿素荧光参数检测,叶绿素荧光参数采用OS-1p便携式脉冲调制荧光测定仪(美国OPTI-sciences 公司)检测,选择每一参试品种的叶片,都以6个叶片作为重复,分别进行初始荧光值(Minimal fluorescence,Fo)、最大荧光值(Maximal fluorescence,Fm)、暗适应最大可变荧光(Fv=Fm-Fo)等有关荧光参数的测定;在叶片测定前充分暗适应30 min,然后测定初始荧光Fo,此时叶片PSII反应中心全部处于开放状态;之后用强饱和脉冲光激发,使原初电子受体全部处于还原状态,激发最大荧光Fm;根据测得的荧光参数,可得PSII的最大光化学效率(Fv/Fm),非光化学猝灭(Non-photochemical quenching,NPQ,叶绿素荧光产量的下降称淬灭,由光合作用引起的荧光淬灭称之为光化学淬灭,由热耗散引起的荧光淬灭称之为非光化学猝灭)按Kooten等[8]的公式计算,
NPQ=Fm′/Fm′-1。
2 结果与分析
2.1 干旱胁迫对杜鹃叶片外观形态的影响
干旱胁迫对杜鹃叶片伤害的轻重程度见表1。从表1可见,在干旱处理5 d时,各杜鹃品种基本上生长良好,仅花蝴蝶部分叶片出现了失水下垂萎蔫现象;在干旱处理10 d时,除紫宸殿、锦袍无明显伤害外,其他品种叶片开始出现明显萎蔫,其中花蝴蝶与紫鹤的萎蔫较为严重,整体植株濒临死亡,因此10 d以后不再进行这2个品种的生理与光合指标检测;至干旱15 d时,胭脂蜜与鸳鸯锦叶片出现了明显萎蔫,而紫宸殿与锦袍仍然处于伤害较轻的状态;至干旱20 d时,所有杜鹃品种均出现大面积叶片萎蔫,但锦袍的伤害程度相对最轻。
2.2 干旱胁迫对杜鹃叶片生理指标的影响
2.2.1 干旱胁迫对杜鹃叶片MDA含量的影响 植物在逆境条件下,往往发生膜脂过氧化作用,MDA是其产物之一,通常利用它作为膜脂过氧化指标,表示细胞膜脂过氧化程度。干旱胁迫对杜鹃叶片MDA含量的影响情况见图1。从图1可知,干旱胁迫会使所有杜鹃品种的MDA含量呈上升趋势或先升后降的变化趋势,其中紫宸殿与锦袍在干旱5 d与15 d出现峰值,并且15 d的峰值要高于5 d的峰值;花蝴蝶与紫鹤在干旱5 d即达到高峰,之后迅速下降;胭脂蜜与鸳鸯锦的MDA含量在干旱胁迫下平缓上升,其中胭脂蜜在10 d达到最大值后平缓下降。
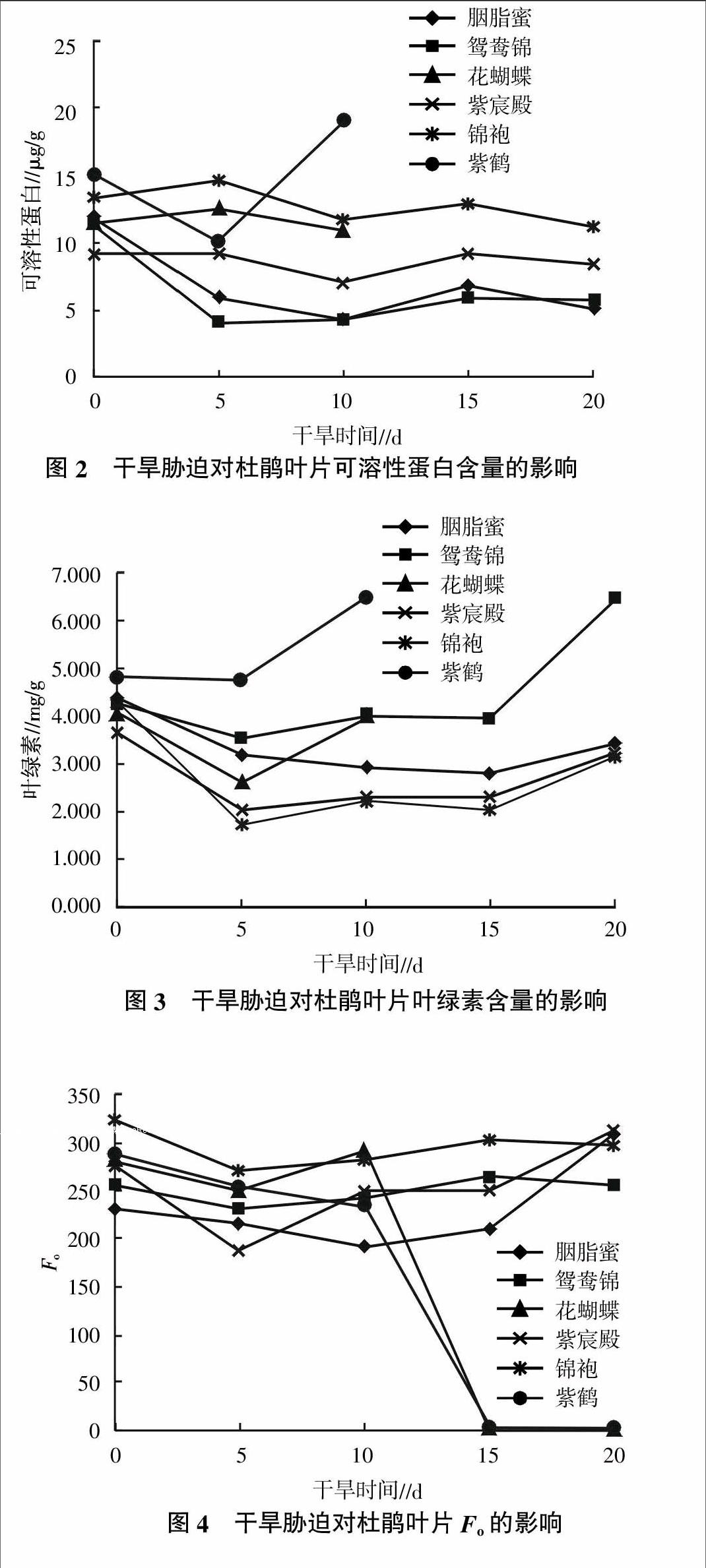
2.2.2 干旱胁迫对杜鹃叶片可溶性蛋白含量的影响 干旱胁迫对杜鹃叶片可溶性蛋白含量的影响情况见图2。从图2可见,杜鹃叶片可溶性蛋白含量除紫鹤先降低后急剧升高以外,大部分品种的可溶性蛋白含量变化较平缓,基本呈现缓和波动状态。其中胭脂蜜、鸳鸯锦与紫宸殿是前期缓慢下降、后期缓慢增加;而花蝴蝶与锦袍是前期缓慢增加、后期缓慢下降。
2.2.3 干旱胁迫对杜鹃叶片叶绿素含量的影响 干旱胁迫对杜鹃叶片叶绿素含量的影响情况见图3。从图3可见,干旱胁迫后不同杜鹃品种叶片叶绿素含量变化趋势基本呈现先下降后上升的变化趋势,其中花蝴蝶与紫鹤在干旱5 d后急剧上升,而其他品种在15 d以后才开始明显上升,以鸳鸯锦在后期上升的幅度相对较大。
2.2 干旱胁迫下杜鹃叶片叶绿素荧光参数的变化
2.3.1 干旱胁迫对杜鹃叶片Fo的影响 Fo是暗适应下的初始荧光,可反映捕光色素复合体(Light Harvest Complex of PSⅡ,LHCII)与PSⅡ的结合状态。干旱胁迫对杜鹃叶片叶绿素荧光参数F0的影响情况见图4。从图4可以看出,干旱胁迫后不同杜鹃品种的Fo变化趋势不同,基本趋势是胁迫初期略有下降,随着胁迫时间延长Fo上升,其中干旱敏感品种花蝴蝶在胁迫5 d后急剧上升,紫鹤则一直下降,都在胁迫5 d后由于叶片失水萎蔫死亡,Fo也检测不到;而其他4个品种在干旱10 d后Fo才开始增加,其中胭脂蜜与紫宸殿在处理15 d后Fo增加幅度加大,而锦袍与鸳鸯锦变化较平缓,这与其叶片外观形态的变化是一致的。
2.3.2 干旱胁迫对杜鹃叶片Fv/F0的影响 Fv/Fo可以反映PSII的潜在活性,干旱胁迫对杜鹃叶片叶绿素荧光参数Fv/Fo的影响情况见图5。从图5可见,干旱胁迫后不同杜鹃品种的Fv/Fo变化规律不同,其中花蝴蝶与紫鹤从干旱初始就一直呈现下降趋势,在5 d后急剧下降;而干旱胁迫10 d前,其他4个杜鹃品种的变化趋势不大,在干旱胁迫10 d后,除鸳鸯锦变化不明显外,其他品种均有小幅增加,但在15 d后均下降。
2.3.3 干旱胁迫对杜鹃叶片Fv/Fm的影响 Fv/Fm表示植物叶片PSII最大或潜在的光化学效率,适宜环境条件并经充分暗适应的叶片Fv/Fm一般保持在0.75~0.85之间,当植物遭受逆境胁迫时,Fv/Fm会明显降低;干旱胁迫对杜鹃叶片叶绿素荧光参数Fv/Fm的影响情况见图6。从图6可见,干旱胁迫后不同杜鹃品种的Fv/Fm变化趋势不同,如紫鹤在干旱处理 5 d后就发生急剧下降,在干旱处理10 d时已接近于0;花蝴蝶在干旱处理10 d后发生急剧下降;而其他品种胭脂蜜、紫宸殿、锦袍、鸳鸯锦在干旱处理过程中变化不明显,一直保持着较稳定的状态,只是在干旱胁迫15 d后略有降低。
3 讨论
植物抗旱能力受多因素影响,测定外观形态、生理指标与光合指标可以较全面地反映植物对干旱逆境的抗性。从外观形态看,杜鹃品种花蝴蝶与紫鹤较早出现伤害症状,而紫宸殿与锦袍较晚出现,表明后两者抗旱性较强。MDA含量与细胞膜相对透性具有相关性,是植物受伤害程度的标志之一;一般MDA含量越高,植物受伤害程度越重。本研究表明MDA含量总体呈上升趋势,其中花蝴蝶与紫鹤是在干旱胁迫初期迅速上升,至峰值后又迅速下降;紫宸殿与锦袍出现2个峰值,这与黄承玲等[4]在迷人杜鹃等高山杜鹃上的研究结果一致,表明植物感受到干旱胁迫后,会启动防御机制来抑制膜脂过氧化进程;后期随着胁迫程度增加,防御系统损伤较大,导致MDA又急剧上升。干旱胁迫使6个杜鹃品种的细胞膜相对透性明显增强,这与前人的研究结果一致[9]。
杜鹃的可溶性蛋白含量变化趋势较平缓,主要呈现先增加后降低或从胁迫初始就持续降低的状态;末期的上升可能是由于叶片水分急剧丧失、以至单位鲜重范围内的叶片数量增加导致。有研究认为,可溶性蛋白具有渗透调节功能,胁迫初期的可溶性蛋白增加可维持细胞保持较低的渗透势,以抵抗水分胁迫带来的伤害[10]。另有报道认为,逆境胁迫诱导了某些调控蛋白表达,导致可溶性蛋白增加[11]。本试验发现参试杜鹃品种可溶性蛋白含量变化平缓,可能是由于杜鹃的可溶性蛋白含量与干旱胁迫关系较弱的缘故。
叶绿素总含量在一定程度上反映了植物同化物质的能力,干旱胁迫下叶绿素含量的变化可以指示植物对干旱胁迫的敏感性。植物受到干旱胁迫后,由于叶片严重失水,常常造成细胞质的破坏,叶绿素也随之降解。研究显示,抗旱性强的植物,其叶片叶绿素含量表现为下降的趋势,而抗性弱的则表现为上升[12],这与本试验结果一致。较早出现伤害症状的花蝴蝶与紫鹤在干旱胁迫初期急剧上升,而其他品种在胁迫前期下降,至胁迫末期才缓慢上升,并且紫宸殿、锦袍与鸳鸯锦的上升幅度较小。
叶绿素荧光参数的测定可以更清晰地显示植物光合作用中对光能的吸收、传递、耗散和分配情况。Fo是暗适应下的初始荧光,一般认为,逆境胁迫下类囊体膜结构改变导致Fo缓慢增加,反映了PSII反应中心失活或LHCII与PSII的分离[13,14]。本试验结果表明,大部分参试杜鹃品种的Fo基本表现为在干旱胁迫初期略微下降,而后期缓慢上升,其中花蝴蝶与紫鹤在10 d后急剧下降至无,这与花蝴蝶与紫鹤较早出现失水萎蔫伤害症状较一致。Fv/Fo可以反映PSII的潜在活性,试验中除对干旱敏感的花蝴蝶与紫鹤Fv/Fo在胁迫5 d后迅速下降外,其他品种一直处于平缓波动状态。Fv/Fm是PSII最大或潜在光化学效率,在逆境胁迫下常被用于指示PSII的受损程度,Fv/Fm的下降与Fo的上升相伴出现是光抑制发生的标志。试验发现Fv/Fm与Fv/Fo变化规律一致,除2个干旱敏感杜鹃品种急剧降低以外,其他品种均保持平稳状态,表明干旱胁迫对杜鹃叶片PSII的伤害不是渐进式的,而是突发式的,在没有达到品种耐热阈值时对PSII没有造成明显伤害,而达到某一阈值时对PSII的伤害可突然导致不可逆转的水平,继而使叶片或整株植物死亡。
参考文献:
[1] 朱春燕,包志毅,唐宇力.杜鹃花赏析[J].生物学通报,2006,41(6):16-17.
[2] 张永辉,姜卫兵,翁忙玲.杜鹃花的文化意蕴及其在园林绿化中的应用[J].中国农学通报,2007,23(9):11-17.
[3] 李 娟.两种杜鹃花的土壤干旱胁迫研究[D].贵阳:贵州师范大学, 2009.
[4] 黄承玲,陈 顺,高贯龙. 3种高山杜鹃对持续干旱的生理响应及抗旱性评价[J].林业科学,2011,47(6):48-55.
[5] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444- 448.
[6] MAXWELL K,JOHNSON G N. ChlorophyⅡ fluorescence-A practical guide[J].Journal of Experimental Botany,2000,51(34):659-668.
[7] 赵会杰,邹 琦,于振文.叶绿素荧光分析技术及其在植物光合机理研究中的应用[J].河南农业大学学报,2000,34(3):248-251.
[8] VAN KOOTEN O,SNELL J F H. Leaf xanthophyll content and composition in sun and shade determined by HPLC[J].Photosynth Res,1990,25:147-150.
[9] 柯世省,杨敏文.水分胁迫对云锦杜鹃光合生理和光温响应的影响[J].园艺学报,2007,34(4):959-964.
[10] 郭华军.水分胁迫过程中的渗透调节物质及其研究进展[J].安徽农业科学,2010,38(15):7750-7753,7760.
[11] 史玉炜,高述民,王燕凌,等.植物干旱诱导蛋白及相关基因的表达与调控[J].新疆农业科学,2007,44(2):126-131.
[12] 李 涛,王 飞,卢 艳,等.4种宿根花卉在自然失水胁迫下的生理变化与抗旱性关系[J].西北农业学报,2010,19(10):146-151.
[13] 罗明华,胡进耀,吴庆贵,等.干旱胁迫对丹参叶片气体交换和叶绿素荧光参数的影响[J].应用生态学报,2010,21(3):619-623.
[14] DEMMING A B,ADAMS W W,BARKER D H,et al. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation[J]. Physiologia Plantarum,1996,98:253-264.
