泉州湾红树林区鱼类物种多样性及其对鱼类生态的支撑作用
2016-12-07郭治明鞠培龙杜建国杨圣云陈明茹肖佳媚
郭治明,陈 杰,鞠培龙,方 聪,杜建国,杨圣云,陈明茹*,肖佳媚
(1.厦门大学海洋与地球学院,福建厦门361102;2.国家海洋局第三海洋研究所,福建厦门361005)
泉州湾红树林区鱼类物种多样性及其对鱼类生态的支撑作用
郭治明1,陈 杰1,鞠培龙1,方 聪1,杜建国2,杨圣云1,陈明茹1*,肖佳媚1
(1.厦门大学海洋与地球学院,福建厦门361102;2.国家海洋局第三海洋研究所,福建厦门361005)
2013—2014年在泉州湾洛阳江口天然红树林区进行4个季节每季连续15 d的鱼类采样,以研究该区域内鱼类的种类组成、区系特征、优势种/类群及多样性指数等的季节变化,并讨论红树林区对鱼类的生态支撑作用.共采集鱼类72种(其中2种鉴定到目,1种鉴定到科,1种鉴定到属,其余均鉴定到种),隶属于9目34科,其中91.3%为暖水性种类,8.7%为暖温性种类;优势种/类群存在季节变化,春、夏季优势种以定居种(如中华乌塘鳢(Bostrychus sinensis)、虎鱼科(Gobiidae)的鱼类等)为主,秋、冬季优势种则以鲻科(Mugilidae)的鱼类、花(Clupanodon thrissa)、七丝鲚(Coilia grayii)等非定居中上层鱼类为主;物种数、物种丰富度指数、多样性指数和均匀度指数的季节变化均为夏季>秋季>冬季>春季.与其他相似的红树林区域比较,泉州湾红树林区的鱼类物种丰富度指数和多样性指数均较高.在此基础上从鱼类体长生长的季节变化角度,讨论了泉州湾红树林区作为鱼类育幼场、索饵场和躲避敌害场所的生态功能,说明红树林生态系统存在完整的海洋食物网,对区内鱼类具有生态支撑作用,强调了保护红树林这一典型生态系统的重要科学意义.
泉州湾;红树林区;鱼类;物种多样性;生态支撑
红树林生态系统是热带、亚热带的一类特殊的潮间带生态系统,林区水深较浅、底质中有机质含量高,初级生产力高,生物资源丰富,幼鱼被捕食率较低.红树林区作为许多近岸鱼类理想的索饵、育幼和避敌场所已成为国内外学者的共识[1-2].
近年来,国内外学者对于红树林区鱼类的研究,已不仅仅局限于红树林区鱼类多样性的调查[3-4],还涉及到红树林与鱼类的关系[5-6]、鱼类在红树林与其他河口生境(如海草床、珊瑚礁等)之间的转移及生态连通性[7-8]、红树林区鱼类对所在海区渔业资源的补充[9-10]等方面的研究,为红树林等特殊生境的保护和渔业资源的修复等提供了大量科学依据.
我国红树林主要分布在广东、广西、海南、福建等省.其中,对于广西和广东红树林区内鱼类的调查研究起步较早,多以鱼类多样性调查和多样性时空变化为主要研究对象[11-12];而对海南东寨港红树林区鱼类的研究目前已较为全面,除对鱼类群落时空变化的研究[13]外,还讨论了有关网具[14]、红树林异质性[15]对渔获物和鱼类多样性的影响等问题.然而,有关福建省红树林区鱼类的研究,目前还鲜有报道.
福建泉州湾洛阳江口是我国东南沿海主要的红树林分布区之一.早期大面积天然红树林由于历年围垦和养殖业的发展,遭受了严重破坏,仅存0.17 km2.虽然现已人工恢复红树林至4 km2以上[16],但当地滩涂养殖业的发展与红树林区保护之间的矛盾依旧十分尖锐.本研究拟通过对泉州湾洛阳江口红树林区内鱼类物种多样性的调查,分析各季节红树林区内鱼类物种多样性和优势种等重要生态学参数的变化,并从育幼、索饵和躲避敌害三方面讨论红树林区与鱼类之间的关系,研究红树林生态系统对鱼类的生态支撑作用,为该典型生态系统鱼类资源和红树林生境的保护提供科学依据.
1 材料与方法
1.1研究区域概况
泉州湾(图1)属亚热带海洋性季风气候,年均降水量为1 095.4 mm,年均温度20℃,极端最低、最高温为0和38.9℃;滩涂以淤泥质为主,盐度较低,平均盐度为19.62;水体受北支潮控制,为正规半日潮,平均潮差4.27 m;内湾面积79.51 hm2,外湾面积56.91 hm2,湿地占99%之多[17].

图1 泉州湾地理位置图Fig.1 Geographic location of Quanzhou Bay
泉州湾适宜红树林生长,主要有红树植物3种,分别是桐花树(Aegiceras corniculata)、秋茄(Kandelia obovata)和白骨壤(Avicennia marina)[16].
1.2采样断面与方法
2013年5、8、11月和2014年2月分别在福建泉州湾3处(洛阳、西方和增垵)天然红树林内潮滩设置调查断面(图2).选择原因是其周围红树植被属于发育较好的天然红树林,受人为干扰较少,可更好地研究自然状态下红树林区的鱼类群落状况.各断面顺着潮水进入的方向分高潮、中潮、低潮设置3个站点,以减少采样的偶然性.按季节周年采集鱼类样品.
每个季节逐日连续采样一潮期(15 d),于每日涨潮前布网,退潮后收网,采集渔获物样品.采样所用网具为“火车网”(又称“蜈蚣网”,是一种定置串联倒须笼),总长约5 m,网目8 mm,于各断面的高潮、中潮、低潮站点各设一个网,共9个.采用“火车网”是由于在红树林区水深较浅、红树植物气生根和支柱根众多的特殊生境下,其相对于其他网具而言更方便作业且能捕获更多种鱼类[13],从而较全面地反映该区域的鱼类群落情况.使用电子天平(精确度为0.01 g)和电子数显游标卡尺(精确度为0.01 mm)测量样品质量和体长等生物学参数.记录数据后,样品经95%(体积分数)乙醇保存后运至实验室进行形态学分类鉴定[18].

图2 采样站位图Fig.2 Sampling sites in Quanzhou Bay mangrove zone
1.3数据处理
物种多样性分析所采用的相关指数及计算公式如下[19-20]:

其中Pi为i种类的生物量(或个数)占总生物量(或总个数)的百分比,S为种类数,N为总个体数.
本文中采用生物量计算多样性指数,分析泉州湾红树林各个季度航次所采得样品的多样性分布特征,并进行季节之间的聚类分析和非度量多维标度排序(NMDS)分析,以判定各季节之间鱼类群落的相似性.通常认为,NMDS分析得出的Stress值小于0.1,则得到的NMDS图形可以正确解释样本间的相似关系;当其值介于0.1~0.2之间,认为其图形有一定的解释意义;当其值大于0.2,则认为其图形不能正确解释群落结构的相似性[21].以上D、H′、J、聚类分析和NMDS分析结果均采用PRIMER V5软件计算得出.
用Pinkas相对重要性指数(IRI)表示优势种的优势度[22]:

其中W为该类鱼质量占鱼类总质量的百分比,n为该类鱼尾数占鱼类总尾数的百分比,F为该类鱼出现的站数占调查总站数的百分比.本研究中以每个季节IRI值大于500的鱼类作为该季节的优势种/类群[23].
应用SPSS 18.0进行t-检验和方差分析,p<0.05表示差异显著,p<0.01表示差异极显著.
2 结果与讨论
2.1种类组成
4个季节共采集到鱼类72种,隶属于9目34科[18],其中2种鉴定到目,1种鉴定到科,1种鉴定到属,其余均鉴定到种(表1).样品中鲈形目种类最多,共20科47种,占总种类数的65.3%,其中虎鱼科的种类最多,共计19种,占27.8%.其余各目中鳗鲡目3科6种,占8.3%;鲱形目2科6种,占8.3%;鲻形目1科5种,占6.9%;鲽形目3科4种,占5.6%;其余各目种类很少.

表1 泉州湾红树林区鱼类种类名录Tab.1 Directory of fish in Quanzhou Bay mangrove zone
2.2鱼类生态特征
根据泉州湾红树林区鱼类组成,本研究主要分析鱼类的栖息水层、食性、适温性和生活方式等生态学特征[24-25].

续表
此外,本研究区域鱼类就生活方式而言,大致可以分为三类[12,26]:1)长居于红树林中的种类(定居种),如虎鱼科的鱼类、中华乌塘鳢、弹涂鱼等,这类鱼通常个体较小且生命周期较短;2)随潮水偶然进出红树林区的种类(偶见种),如沟鲹、勒氏笛鲷等,这些鱼个体数极少,对群落结构的影响基本可以忽略;3)仅在个体发育的某个阶段(通常是稚幼鱼阶段)进入红树林的近岸或河口种类(近岸种),以鲱科、鳀科和鲻科等中上层鱼类为代表.
2.3鱼类群落结构的季节变化
2.3.1优势种/类群的季节变化
依据各季节IRI值大于500的鱼类为该季节的优势种/类群的判定方法,泉州湾红树林区鱼类各季节优势种/类群研究结果如表2所示.
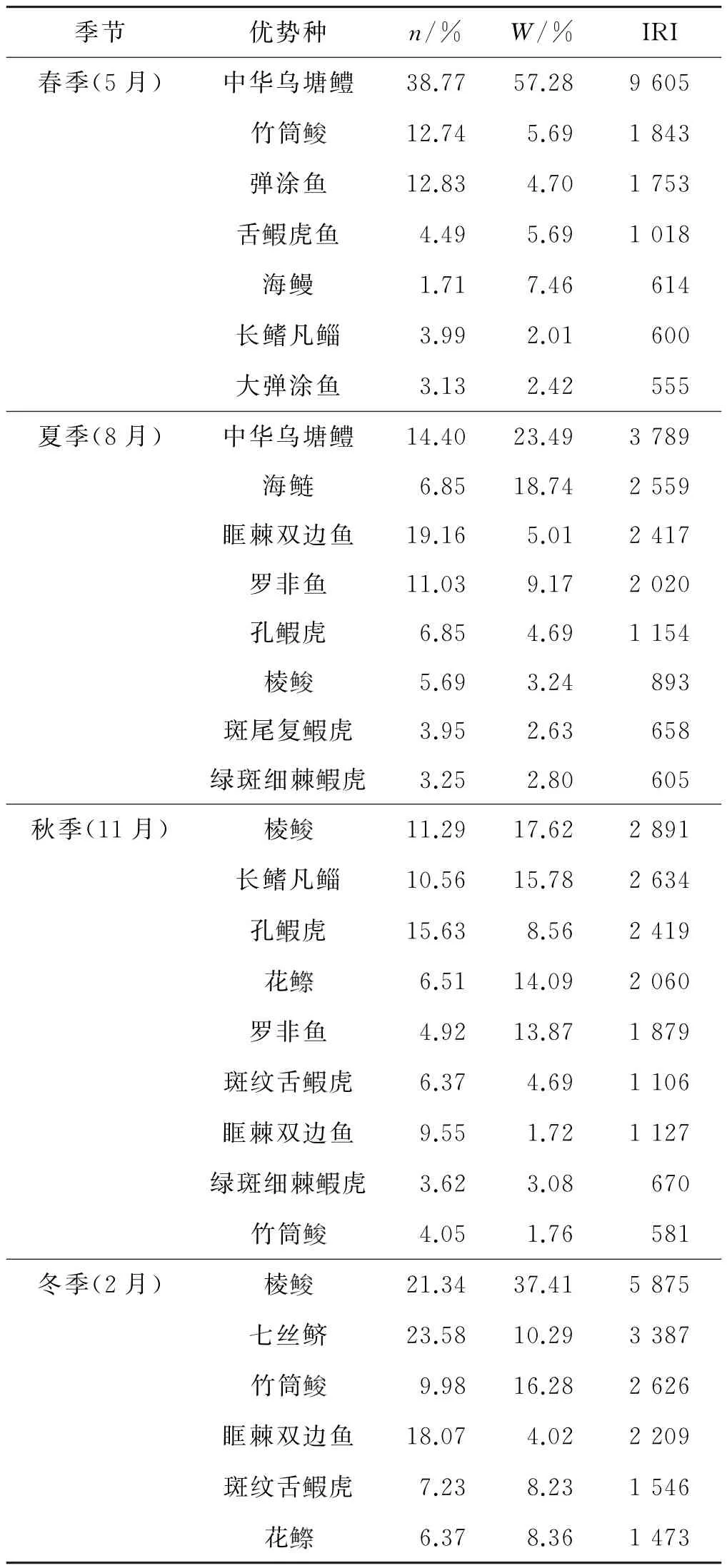
表2 泉州湾红树林区鱼类各季节优势种/类群Tab.2 Dominant species in four seasons in Quanzhou Bay mangrove zone
由表2可见,泉州湾红树林区鱼类优势种类群存在明显的季节变化:优势种类群中定居鱼类(如虎鱼科的鱼类和中华乌塘鳢、弹涂鱼等)所占比例春季>夏季>秋季>冬季;而到了秋、冬季,林区则以中上层鱼类(如棱、竹筒、七丝鲚等)为主.
2.3.2物种数及多样性的季节变化
各季节鱼类多样性计算结果如表3所示.可以看出,泉州湾红树林区鱼类物种数的季节变化为:夏季>秋季>冬季>春季;总个体数和总生物量季节变化为:春季>夏季>秋季>冬季;D的季节变化为:夏季>秋季>冬季>春季;H′的季节变化为:夏季>秋季>冬季>春季;J的季节变化为:夏季=秋季>冬季>春季.
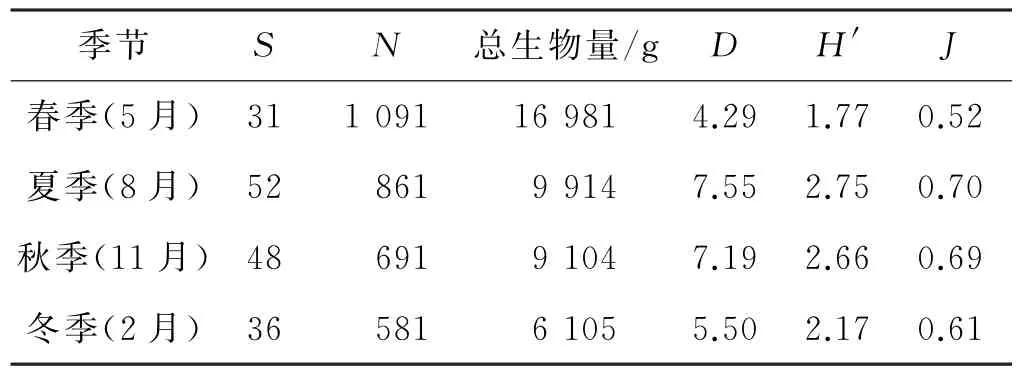
表3 泉州湾红树林区各季节鱼类多样性指数Tab.3 Index offish biodiversity in four seasons in Quanzhou Bay mangrove zone
2.3.3各季节鱼类群落的相似性
根据各季节所得鱼类的种类及各种类的生物量数据,通过季节间聚类分析和NMDS分析比较各季节之间鱼类群落的相似性,结果如图3所示.可以看出,夏、秋季鱼类群落的相似性最高,其次为春季和夏季;冬季与其他三季相比,相似性均较低.NMDS分析也得出了类似的结果,即夏、秋两季由于鱼类群落的高相似性而被划分到同一组,其他两季则分别单独成一组,检验系数Stress值为0(<0.1),说明NMDS图可以很好地表示各季节间鱼类群落的相似性.
结合上文分析结果,可以看出泉州湾红树林区鱼类群落结构的季节变化:春、夏季适逢红树林定居种的繁殖期,泉州湾红树林区聚集了大量定居种,某些种类在个体数和生物量上均占较大优势(如中华乌塘鳢),而进入红树林区的近岸种相对较少.秋、冬季红树林区多数定居种过了繁殖期,且在冬季定居种多栖息于洞穴中,故其个体数和生物量较春、夏季大幅度下降;而近岸种比例相对增多,特别是一些中上层鱼类的幼鱼(如鲱科、鲻科鱼类等),这些种类成为了秋、冬的主要优势种.
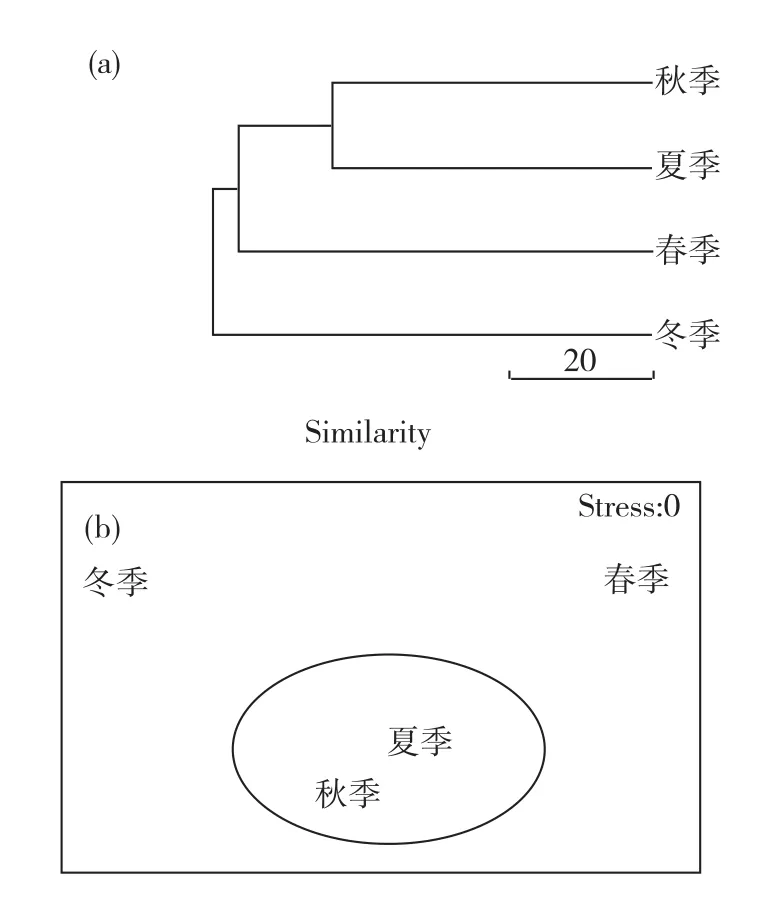
图3 泉州湾红树林区各季节鱼类群落结构聚类分析(a)和NMDS分析(b)Fig.3 Cluster analysis(a)and NMDS analysis (b)of fish community structure in four seasons in Quanzhou Bay mangrove zone
由此可见,泉州湾红树林区在秋、冬季对近岸中上层稚幼鱼有更大的吸引力.其原因一方面是秋、冬季林区定居种过了繁殖期,且多数定居种在冬季有穴居的习性,使得进入林区的中上层稚幼鱼拥有更多空间和资源;另一方面是红树林区物理条件较其他生境温和,缓冲能力强,更适合稚幼鱼度过寒冷的季节.
2.4与其他相似红树林区及邻近海区鱼类生物多样性的比较
如表4所示:与其他相似的红树林区域比较,泉州湾红树林区鱼类的D和H′均较高(仅低于海南东寨港),这可能与本研究周年且连续一个潮周期的采样方法有关,一些随着潮水进入红树林的数量较少的偶见种在一定程度上使得H′和D值偏高;与泉州湾海域比较,本研究区域的H′和J也较高,说明相对于周边海区,泉州湾红树林区的鱼类物种多样性更高.

表4 不同区域鱼类多样性指数比较Tab.4 Comparion of fish diversity index in different areas
2.5泉州湾红树林生态系统对鱼类的生态支撑
如前文所述,泉州湾红树林区对近岸中上层鱼类的稚幼鱼具有较强的吸引力,在秋、冬季林区更是以这些中上层稚幼鱼占较大优势.另外,对比于泉州湾海域,本研究区域内鱼类的H′和J′均较高,说明泉州湾红树林区除了适合某些定居鱼类(如弹涂鱼、中华乌塘鳢、虎鱼科的鱼类)长期定居之外,还适合许多中上层鱼类(尤其是其稚幼鱼)在此栖息.稚幼鱼在红树林中具有更高的存活率和摄食率,而且红树林生态系统以其特殊的结构及生产力,对栖息于该区的鱼类在育幼、索饵以及躲避敌害等方面具有生态支撑作用.
2.5.1鱼类育幼场
比较泉州湾红树林区几种较常见中上层鱼类的体长与所在海区成鱼体长[29],结果如表5所示.根据t-检验结果可以看出,红树林区几种常见的中上层鱼类体长均极显著地低于所在海区成鱼的体长,多数种类样品的最大体长与其成鱼的平均体长比较仍有较大差距,说明除少数个体较小的定居种以成鱼为主外,泉州湾红树林区鱼类以稚幼鱼为主,这与其他研究结果[30]类似.同时表明泉州湾红树林生态系统是近岸某些中上层鱼类理想的育幼场,如海鲢、鲱科、鳀科和鲻科鱼类在其幼鱼阶段常分布于红树林区.
2.5.2幼鱼索饵场
泉州湾红树林位于洛阳江和晋江的入海口,潮汐为正规半日潮,随潮水涨落和陆地径流流入,林区内聚集了大量营养物质,构成了泉州湾红树林初级生产力的基础.而红树林区的红树植物、附生植物、浮游植物以及底栖藻类等产生大量有机碎屑物质,构成许多食碎屑的稚幼鱼的主要饵料,同时在红树林区土壤中还原微生物的参与下,这些碎屑物质可直接为红树林区碎屑食物网提供有机质和能量.
此外,红树林区存在大量浮游植物、浮游动物以及浮游幼体(如甲壳类幼体)[2],是林区内许多稚幼鱼的主要饵料.泉州湾红树林区有肉食性鱼类53种,占总种类数的76.8%;其余的为杂食性种类,共16种,占23.2%.其中许多鱼类(如鲱科、鳀科和鲻科的鱼类)均摄食浮游植物、浮游动物以及浮游幼体.
以上分析说明,红树林生态系统中不同营养级的海洋生物之间构成完整的海洋食物网.
2.5.3鱼类躲避敌害场所
泉州湾红树林区所采的鱼类样品中,仅海鳗、食蟹豆齿鳗、长尾蛇鳗、日本花鲈、黄鳍鲷5种是以其他鱼类为食的肉食性鱼类,但基本为稚幼鱼,除海鳗外,其他肉食性鱼类均罕见.林区内未发现大型捕食者,说明该林区更适合小型鱼类及稚幼鱼的生长.相对于红树林外的海域,泉州湾红树林内水深较浅,而个体较大的捕食者往往只分布于较深的水域以获得更高的捕食效率,很少进入较浅的水域[32].因此小型鱼类如稚幼鱼可进入水深较浅的红树林区有效地躲避捕食者.另外,红树林内水体的浑浊度较高,能见度较低,也是限制捕食者捕食的一个重要因素[33].由此可见,红树林可有效降低区内鱼类的被捕食率,因而成为许多稚幼鱼和小型鱼类理想的躲避敌害场所.

表5 红树林区常见中上层鱼类体长与所在海区成鱼平均体长的t-检验比较Tab.5 t-test results of body length between common pelagic fish in mangrove zone and in nearby sea area

表6 常见鲻科鱼类各季节体长差异的方差分析Tab.6 ANOVA results of body length of common Mugilidae fish in different seasons
3 结 论
综上所述,本研究表明,泉州湾红树林区中鱼类的优势种、物种数及多样性指数均呈现明显的季节变化,主要表现为:1)春、夏以定居种类占较大优势(如中华乌塘鳢、虎鱼科鱼类等),秋、冬季则以非定居的中上层稚幼鱼占较大优势(如鲻科鱼类、七丝鲚、花等);2)物种数和各多样性指数(H′,D,J)季节变化趋势为夏季>秋季>冬季>春季.
此外,泉州湾红树林区对进入该区域的鱼类具有重要的生态支撑作用,是许多近岸中上层鱼类理想的育幼、索饵和避敌的场所.同其他红树林生境一样,泉州湾红树林区对污染物质具有净化作用,林区内海水流速低,受海浪和潮汐影响较小,红树林对风浪缓冲能力较强[34],极端灾害天气也少.这些适宜的物理化学环境使得泉州湾红树林区为林内稚幼鱼提供类相对稳定的生态环境,有利于其育幼、索饵和生长.这些稚幼鱼生长到一定阶段后,离开红树林区,补充到邻近海域的渔业资源中去.可见,保护红树林生态系统具有重要的科学和实践意义.
[1] TSE P,NIP T H M,WONG C K.Nursery function of mangrove:a comparison with mudflat in terms of fish species composition and fish diet[J].Estuarine,Coastal and Shelf Science.2008,80(2):235-242.
[2] LAEGDSGAARD P,JOHNSON C R.Mangrove habitats as nurseries:unique assemblages of juvenile fish in subtropical mangroves in eastern Australia[J].Marine Ecology Progress Series,1995,126:67-81.
[3] 何斌源,范航清,莫竹承.广西英罗港红树林区鱼类多样性研究[J].热带海洋学报,2001,20(4):74-79.
[4] 黄德练,吴志强,黄亮亮,等.广西钦州港红树林区鱼类物种多样性分析[J].海洋湖沼通报,2013(4):135-142.
[5] 施富山,王瑁,王文卿,等.红树林与鱼类关系的研究进展[J].海洋科学,2005,29(5):54-59.
[6] NANJO K,NAKAMURA Y,HORINOUCHI M,et al. Predation risks for juvenile fishes in a mangrove estuary: a comparison of vegetated and unvegetated microhabitats by tethering experiments[J].Journal of Experimental Marine Biology and Ecology,2011,405(1):53-58.
[7] KIMIREI I A,NAGELKERKEN I,GRIFFIOEN B,et al. Ontogenetic habitat use by mangrove/seagrass-associated coral reef fishes shows flexibility in time and space[J]. Estuarine,Coastal and Shelf Science,2011,92(1):47-58.
[8] VASLET A,PHILLIPS D L,FRANCE C,et al.The relative importance of mangroves and seagrass beds as feeding areas for resident and transient fishes among different mangrove habitats in Florida and Belize:evidence from dietary and stable-isotope analyses[J].Journal of Experimental Marine Biology and Ecology,2012,434/ 435:81-93.
[9] ISLAM M S,IKEJIMA K.Gear type,species composition and economic value of fisheries in the mangroves of Pak Phanang,Thailand[J].Wetlands Ecol Manage,2010,18 (1):27-36.
[10] 刘超,胡文佳,陈明茹,等.山口红树林区稚幼鱼多样性及其对渔业资源的补充作用[J].厦门大学学报(自然科学版),2013,52(2):274-280.
[11] 何斌源.广西两港湾红树林鱼类生态的比较研究[J].海洋通报,1999,18(1):29-35.
[12] 叶宁,吴晓东,张苇.湛江高桥红树林区鱼类调查[J].广东海洋大学学报,2007,27(6):56-61.
[13] 施富山.海南东寨港国家级红树林保护区鱼类生态学研究[D].厦门:厦门大学,2005:1-10.
[14] 王瑁,张尽函,施富山.海南东寨港红树林区的渔具及渔获物调查[J].水产科技情报,2007,34(1):6-10.
[15] 颜素贞.红树林微生境异质性对鱼类对鱼类多样性的影响[D].厦门:厦门大学,2011:46-50.
[16] 许建林.福建泉州湾红树林生态系统价值评价初探[J].农技服务,2009,26(9):103-104.
[17] 陈文堃.2011年泉州市海洋环境状况公报[R].泉州:泉州市海洋与渔业局,2012.
[18] 朱元鼎.福建鱼类志[M].福州:福建科学技术出版社,1984.
[19] LUDWIG J A,REYNOLDS J F.Statistical ecology[M]. New York:John Wiley&Sons,1988:102-111.
[20] 马克平.生物多样性研究的原理和方法[M].北京:中国科学技术出版社,1994:141-166.
[21] CLARKE K R.Non-parametric multivariate analysis of changes in community structure[J].Australian Journal of Ecology,1993,18:117-143.
[22] PINKAS L,OLIPHANT M S,IVERSON I L K.Food habits of albacore,bluefin tuna,and bonito in California waters[J].California Department of Fish and Game Fish Bulletin,1971,152:101-105.
[23] 陈国宝,李永,陈新军.南海主要珊瑚礁水域的鱼类物种多样性研究[J].生物多样性,2007,15(4):373-381.
[24] 卢振彬,戴泉水,肖方森,等.闽南—台湾浅滩渔场鱼类及其不同生态类群的资源生产量[J].水产学报,2006,30 (3):360-366.
[25] 张雅芝,李福振,刘向阳.东山湾鱼类食物网研究[J].台湾海峡,1994,13(1):53-55.
[26] MANSON F J,LONERANGAN N R,SKILLETER G A,et al.An evaluation of the evidence for linkages between mangroves and fisheries:a synthesis of the literature and identification of research directions[J].Oceanography and Marine Biology:An Annual Review,2005, 43:485-515.
[27] WERNER E E,GILLIAM J F,HALL D J,et al.An experimental test of the effects of predation riskon habitat use in fish[J].Ecology,1983,64(6):1540-1548.
[28] 杜建国,陈彬,卢振彬,等.泉州湾海域鱼类多样性及营养级变化[J].生物多样性,2010,18(4):420-427.
[29] LAEGDSGAARD P,JOHNSON C.Why do juvenile fish utilize mangrove habitats?[J].Journal of Experimental Marine Biology and Ecology,2001,257:229-253.
[30] LITTLE M C,REAY P J,GROVE S J.The fish community of an east African mangrove creek[J].Journal of Fish Biology,1988,32:729-747.
[31] LONERAGAN N,BUNN S,KELLAWAY D.Are mangroves and seagrasses sources of organic carbon for penaeid prawns in a tropical Australian estuary?A multiple stable-isotope study[J].Marine Biology Progress Series,1997,130(2):289-300.
[32] BOESCH D F,TURNER R E.Dependence of fishery species on salt marshes:the role of food and refuge[J]. Estuaries,1984,7:460-468.
[33] COCHERET DE LA MORINIÈRE E,NAGELKERKEN I,VAN DER MEIJ H,et al.What attracts juvenile coral reef fish to mangroves:habitat complexity or shade?[J]Marine Biology,2004,144:139-145.
[34] WOLANSKI E,MOORE K,SPAGNOL S,et al.Rapid, human-induced siltation of the macro-tidal Ord River estuary,western Australia[J].Estuarine,Coastal and Shelf Science,2001,53(5):717-732.
Fish Biodiversity and Ecological Support for Fishes in Quanzhou Bay Mangrove Zone
GUO Zhiming1,CHEN Jie1,JU Peilong1,FANG Cong1,DU Jianguo2,YANG Shengyun1, CHEN Mingru1*,XIAO Jiamei1
(1.College of Ocean&Earth Sciences,Xiamen University,Xiamen 361102,China; 2.Third Institute of Oceanography,State Oceanic Administration,Xiamen 361005,China)
Seasonal changes in species composition,ecotype,dominant species and biodiversity of fishes were analyzed in this study. Sampling was conducted in Quanzhou Bay mangrove zone in 4 seasons year-round from 2013 to 2014.A total of 72 fish species were recorded,belonging to 34 families,9 orders and 1 class,with 8.7%warm-temperate species and 91.3%warm-water species.Investigation showed that the dominant species varied with season.Dominant species in spring and summer mainly consisted of resident species,such as Bostrychus sinensis,Gobiidae spp.,etc.;while dominant species in autumn and winter mainly consisted of non-resident and pelagic species,such as Mugilidae spp.,Clupanodon thrissa,Coilia grayii,etc..Number of species,the species richness index (D),Shannon-Wiener index(H′)and Pielou evenness index(J)showed a tendency of summer>autumn>winter>spring.The species richness index(D)and Shannon-Wiener index(H′)in Quanzhou bay mangrove zone were relatively high,compared with other similar mangrove areas.In addition,the ecological function of Quanzhou bay mangrove zone for fishes was discussed based on nursery,feeding and shelter.Our study suggests that the ecological support of Quanzhou bay mangrove zone for fishes is important and there is a complete oceanic food web in Quanzhou bay mangrove zone,which emphasizes the importance of the protection of mangrove.
Quanzhou Bay;mangrove zone;fishes;biodiversity;ecological support
Q 145+.2
A
0438-0479(2016)06-0860-09
10.6043/j.issn.0438-0479.201508008
2015-08-11 录用日期:2015-11-10
海洋公益性行业科研专项(201305030-4,201505027-2)
mrchen@xmu.edu.cn
郭治明,陈杰,鞠培龙,等.泉州湾红树林区鱼类物种多样性及其对鱼类生态的支撑作用[J].厦门大学学报(自然科学版),2016,55(6):860-868.
GUO Z M,CHEN J,JU P L,et al.Fish biodiversity and ecological support for fishes in Quanzhou Bay mangrove zone [J].Journal of Xiamen University(Natural Science),2016,55(6):860-868.(in Chinese)
