贝加尔针茅光合特征与叶片功能特性对长期氮添加的响应
2016-12-06刘红梅李洁皇甫超河陈新微杨殿林
刘红梅,李洁,皇甫超河,陈新微,杨殿林
(农业部环境保护科研监测所,天津300191)
贝加尔针茅光合特征与叶片功能特性对长期氮添加的响应
刘红梅**,李洁**,皇甫超河,陈新微,杨殿林
(农业部环境保护科研监测所,天津300191)
以贝加尔针茅草甸草原建群种贝加尔针茅为研究对象,通过一个连续6年氮添加野外控制试验,设置5个氮素处理:N0(0 kg N/hm2)、N30(30 kg N/hm2)、N50(50 kg N/hm2)、N100(100 kg N/hm2)、N150(150 kg N/hm2),探讨氮沉降变化对贝加尔针茅种群光合特性与叶片功能特性变化规律及其影响因子。结果表明,氮素添加变化显著影响贝加尔针茅的净光合速率等光合特性,氮素添加处理净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、蒸腾速率(transpiration rate,Tr)、光合氮利用效率(photosynthetic nitrogen use efficiency, PNUE)和光合能量利用效率(photosynthetic energy use efficiency, PEUE)均低于或显著低于无氮素添加处理。氮素添加处理叶片比叶面积(specific leaf area, SLA)、叶片N含量(leaf N content,Nmass)、建成成本(leaf construction cost,CCmass)、叶片N∶P均高于或显著高于无氮素添加处理。相关分析表明,5个氮素处理的Pn与Gs、Tr、PNUE、PEUE、叶片P含量(leaf P content,Pmass)、土壤N∶P呈显著正相关,与叶片比叶面积(SLA)、Nmass、CCmass、土壤含水量、土壤pH值呈极显著负相关;叶片Nmass与SLA、CCmass、土壤N∶P呈显著正相关,与PNUE、PEUE、Pmass、土壤含水量、土壤pH值呈显著负相关。综合以上研究表明,长期氮添加会降低贝加尔针茅净光合速率、养分利用效率,提高叶片建成成本和叶片N∶P,土壤含水量和土壤pH值是引起这种改变发生的主要因子。
氮添加;贝加尔针茅;光合特性;叶片功能特性
高氮沉降的增加呈现出全球化的趋势,成为科学家重点关注的热点生态问题之一[1]。我国已成为继北美和欧洲之后第三大沉降集中区域,在过去的30年内年均氮沉降高达8 kg/hm2[2]。化肥的使用和工业氮沉降向生态系统中输入了大量氮元素,生态系统由原来的受氮限制转变为氮饱和,由于营养元素的失衡,植物生长受到其他元素或者资源的限制。国内外一些研究认为,氮元素的大量添加会改变生态系统中植物群落的结构和组成[3-5]、功能性状[6-7],进一步影响其生态学过程。
光合作用是植物生长发育的基础和生产力高低的决定性因素,是植物体将太阳能转化为化学能并贮存在植物体内的过程。光合作用显著影响植物叶片的发育、化学成分、生物产量形成等各种生理过程[8],是植物对环境影响研究的重要指标。氮素对植物光合器官的合成和活性具有十分重要的调节作用[9],影响光合能量的分配和利用效率[10]。由于人类活动引起的大气氮沉降的增加必然会改变植物的生长发育。一些模拟氮沉降的实验结果表明,氮素输入对植被的生长有明显的促进作用[11-12]。也有一些研究表明,过量氮输入会引起土壤的酸化,导致土壤pH值降低,引起土壤中磷及一些其他盐基阳离子的有效性降低,引起植物中养分的失衡,从而会抑制植物的生长发育[13]。大量研究发现,氮添加会显著增加草原植物叶片N∶P[14-15]。长期氮添加会改变不同植物种群的生长速率,使植物群落组成和多样性发生变化[3,16],而这些改变大多数是由于土壤中养分的变化所致。北美和欧洲的氮施肥实验表明,有效N的增加会提高养分含量高的物种的生长速率和优势度,同时减少本地种的物种丰富度[17]。氮沉降可以改变植物体内C、N、P的化学计量特征,使得土壤全N含量、土壤N∶P提高。
贝加尔针茅(Stipabaicalensis)草原是欧亚大陆草原的重要组成部分,是内蒙古草甸草原的代表类型之一,在我国畜牧业生产中占有重要的地位[18]。贝加尔针茅草原处于氮沉降较为严重的区域,受氮沉降影响显著。贝加尔针茅作为贝加尔针茅草地的建群种,其光合特征和叶片功能特性变化必然会对贝加尔针茅草地生态系统功能的变化起到决定性的作用。目前对于贝加尔针茅种群光合特征和叶片功能特性对长期氮沉降做出的响应,还不十分清楚。本研究以贝加尔针茅草甸草原建群植物贝加尔针茅为研究对象,在自然生境中通过模拟氮沉降发生,比较分析长期不同氮添加处理贝加尔针茅光合特性、叶片比叶面积(specific leaf area, SLA)、建成成本(leaf construction cost,CCmass)、叶片N含量(leaf N content,Nmass)、叶片P含量(leaf P content,Pmass)等的差异,并综合分析不同氮素添加处理下贝加尔针茅光合特性指标和叶片特性、土壤理化性质之间的相关性,从植物生理生态学的角度阐述氮沉降对草地生态系统的影响机制,对预测未来氮沉降对贝加尔针茅草地生态功能的影响和维持当地畜牧业稳定生产具有重要的意义。
1 材料与方法
1.1 研究区概况与样地设置
研究区位于内蒙古自治区鄂温克自治旗(48°27′-48°35′ N,119°35′-119°41′ E),属于半干旱大陆性季风气候,年均气温-1.6 ℃,大于0 ℃年积温2567.5 ℃,无霜期113 d,年均降水量328.7 mm,年蒸发量1478.8 mm,土壤类型为暗栗钙土[19]。主要植被类型为贝加尔针茅群落,共有66种植物类型,建群种是贝加尔针茅,常见种有草地麻花头(Serratulayamatsutanna)、羊草(Leymuschinensis)、柴胡(Bupleurumchinense)、线叶菊(Filifoliumsibiricum)、洽草(Koeleriaaltaica)、扁蓿豆(Melissitusruthenica)、寸草苔(Carexduriuscula)、日荫菅(Carexpediformis)、冷蒿(Artemisiafrigida)、多茎野豌豆(Viciamulticaulis)、肾叶唐松草(Thalictrumpetaloideum)等。
试验布设开始于2010年6月,在围栏样地内设置氮素添加试验,共设 5 个水平依次为:0 kg N/hm2(N0)、 30 kg N/hm2(N30)、50 kg N/hm2(N50)、100 kg N/hm2(N100)、150 kg N/hm2(N150),每个处理 6 次重复,小区面积 8 m×8 m,小区间设2 m 隔离带。氮素为硝酸铵(NH4NO3),每年分两次施入,第1次6月15 日施氮50%处理水平;第2次7月15日施氮50%。通过计算,将每个小区所需要添加的硝酸铵充分搅拌溶解于8 L水中,均匀喷施到小区内,两次施肥增加的水量相当于增加降水1.0 mm。对照N0小区喷洒相同量的水。
1.2 样品采集与测定
试验于2015年8月中旬进行田间土壤样品采集、叶片比叶面积和光合指标测定。土壤样品采集用直径 2.5 cm土钻,按S型选取10 个点,采取0~10 cm 土壤样品混匀,将植物根系和土壤入侵物去除,用“四分法”选取 1 kg 土壤,分成两部分,一部分保存于低温冰盒中,运回实验室在-20 ℃冰柜中保存,用于土壤速效养分和含水量分析;另一部分土样常温保存,运回实验室,室内自然风干后研磨过筛,用于土壤理化性质分析。经过连续6年氮添加处理后,土壤理化指标测定见表1。

表1 不同氮素处理下土壤理化性质
注:同行不同字母表示差异显著(P<0.05)。下同。
Note: Different letters in a line indicate the significant differences atP<0.05. The same below.
叶片比叶面积测定:在选定的5个处理6个重复小区内选取生长相对一致的植株各15株,保存于低温冰盒中,采用叶面积仪(Li-3100A)测定叶面积。将测定完叶面积的叶片带回实验室,105 ℃杀青30 min,然后65 ℃(72 h)烘干至恒重,测定叶片干重。叶片比叶面积按以下公式计算:
SLA=ULA/DY
(1)
式中:ULA表示单位叶片面积,单位cm2;DY表示叶片干重,单位g;SLA表示比叶面积,单位cm2/g。
光合-光强响应测定:在晴朗天气的上午9:00-11:00每小区内选取贝加尔针茅处于营养生长期的植株。采用便携式光合测定仪(LI-6400)对植株叶片的净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、蒸腾速率(transpiration rate,Tr)等指标进行测定[20]。将光合测定仪叶室的温度设定成25 ℃,CO2浓度控制在400 μmol/mol,光量子通量密度(photosynthetic photon flux density, PPFD)设定为800 μmol/(m2·s)。测定时尽量保持叶片的自然生长角度不变,每片叶测定3次,每个处理小区重复测定15次。水分利用效率按照以下公式计算:
WUE=Pn/Tr
(2)
土壤和叶片元素的测定:土壤样品自然风干后,研磨,分别过0.85和0.15 mm筛并保存。植物样品叶片粉碎过0.15 mm筛,保存在塑封袋中。植物样品和土壤样品全N含量用凯氏定氮法测定,全P用钼锑抗比色法测定。土壤含水量用烘干法,土壤有机C采用重铬酸钾氧化-比色法测定,土壤pH采用酸度计法(1∶2.5 土水比)[21],土壤NH4+-N和NO3--N含量采用流动分析仪(QC8000)测定。
叶片灰分浓度测定:将经过烘干处理和过筛的植物样品在马弗炉中500 ℃灼烧6 h,将剩余的残渣称重。干重热值用氧弹式热量计(HWR-15E)测定:称取0.5 g植物样品,经压片完全燃烧后测定干重热值,每个处理小区的样品分别测定6个重复,测定前用苯甲酸标定[22]。
Hc=CV/(1-Ash)
(3)
CCmass=[(0.06968Hc-0.065)(1-Ash)+7.5(kN/14.0067)]/EG
(4)
式中:Hc表示去灰分热值,单位为kJ/g;CV表示干重热值,单位为kJ/g;Ash表示灰分浓度,单位为%;CCmass表示叶片建成成本,单位为g glucose/g;N表示有机氮浓度,单位为mg/g;EG表示生长效率;k为N的氧化态形式(若为NH4+,k=-3;若为NO3-,k=5)。不同植物物种的生长效率为0.87[23],对于单个植物样品,先以NO3-和NH4+作为N的氧化态分别进行计算,然后按照土壤中NO3--N和NH4+-N的比例,求加权平均值作为叶片CCmass。
CCarea=CCmass/SLA
(5)
Narea=Nmass/SLA
(6)
PNUE=Pn/Narea
(7)
PEUE=Pn/CCarea
(8)
式中:CCarea表示叶片单位面积建成成本,单位为g glucose/m2;Narea表示叶片单位面积氮含量,单位为g/m2;PNUE表示光合氮利用效率,单位为μmol CO2/(g·s);PEUE表示光合能量利用效率,单位为μmol CO2/(g glucose·s)。
1.3 数据分析
利用Excel 2010对观测数据进行整理,在数据预处理过程中,去掉极端值。应用SPSS 16.0进行方差分析和相关性分析,统计数据以平均值和标准差表示。
2 结果与分析
2.1 不同氮素添加下贝加尔针茅气体交换参数的比较
在有效光合辐射强度为800 μmol/(m2·s)时,5种氮素处理的贝加尔针茅的气体交换参数测定结果列于表2。
由表2可以看出,N30、N50、N100、N150处理贝加尔针茅的Pn分别显著低于对照N022.38%、36.86%、31.12%和59.93%(P<0.05),说明在相同的有效光合辐射下,N0处理的贝加尔针茅叶片光合同化能力更强。N50、N100、N150处理贝加尔针茅的Gs和Tr都显著低于对照N0(P<0.05),N30处理贝加尔针茅的Gs和Tr都低于对照N0,但无显著差异(P>0.05)。N100处理贝加尔针茅的WUE显著高于其他4种氮素处理,N0、N30、N50和N150处理的WUE无显著差异(P>0.05)。氮添加量越大,Pn、Tr、Gs三者降低的幅度也越大。说明氮添加对贝加尔针茅的Pn、Tr和Gs有极大影响。
2.2 不同氮素添加下贝加尔针茅叶片特性的比较
氮素添加显著影响了贝加尔针茅的叶片SLA、Nmass、叶N∶P和CCmass。由表3可以看出,贝加尔针茅叶片SLA随着氮素添加量的增大表现为先升高后降低的趋势,N50、N100和N150处理的叶片SLA显著高于对照N0(P<0.05),N30处理的SLA高于对照N0,但无显著差异(P>0.05)。Nmass随着氮素添加量的增大而增加,N30、N50、N100和N150处理的叶N含量分别显著高于对照N038.84%、38.97%、56.86%和64.96%(P<0.05)。Pmass呈逐渐降低趋势。叶N∶P、CCmass随着氮素添加量的增大而增大。叶片Ash随着氮素添加量的增大而减小,N30、N50、N100和N150处理的叶Ash显著低于对照N0(P<0.05)。5种氮素处理的叶片Hc之间没有显著差异(P>0.05)。PEUE、PNUE没有一致的变化规律,N30、N50、N100和N150的PEUE和PNUE均显著低于对照N0(P<0.05)。
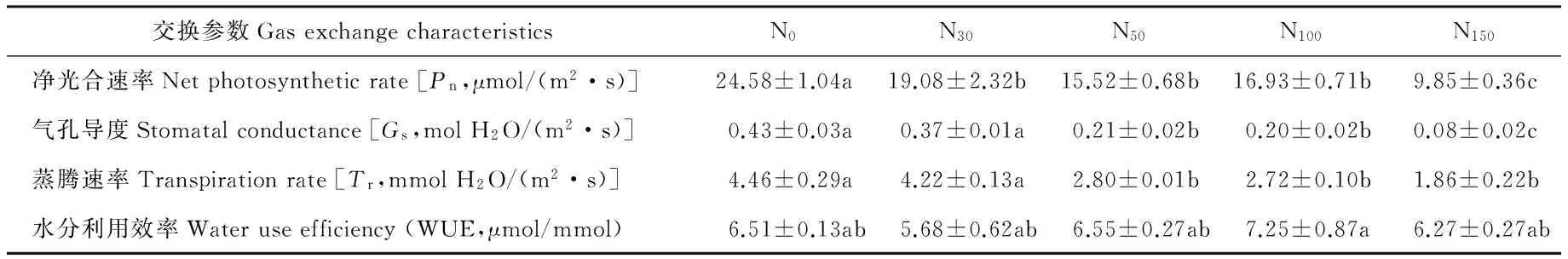
表2 不同氮素添加下贝加尔针茅的气体交换参数

表3 不同氮素添加下贝加尔针茅叶片特性的比较
2.3 不同氮素添加下贝加尔针茅气体光合参数的相关性
Gs表示植物叶片气孔张开的程度,对光合作用、蒸腾作用均产生影响。植物叶片与外界进行的气体交换都是通过气孔进行的[24]。5种氮素添加下贝加尔针茅气体光合参数的相关分析表明(表4),Pn与Tr、Gs呈极显著正相关关系(P<0.01);Gs与Tr呈极显著正相关关系(P<0.01);WUE与Pn、Gs、Tr无显著相关性(P>0.05)。
2.4 不同氮素添加下贝加尔针茅Pn与叶片Nmass、Pmass、SLA、CCmass、PEUE、PNUE之间的相关性
不同氮素添加下贝加尔针茅Pn与叶片功能指标之间的相关分析表明(表5),在不同氮素添加下贝加尔针茅Pn与PNUE、PEUE、Pmass呈极显著正相关(P<0.01),与叶片SLA、Nmass、CCmass之间呈极显著负相关(P<0.01);贝加尔针茅Nmass与叶片SLA、CCmass呈极显著正相关(P<0.01),与叶片Pmass呈显著负相关(P<0.05);叶片CCmass与叶片SLA之间呈极显著正相关(P<0.01)。
2.5 不同氮素添加下Pn、叶片功能性状与土壤理化性质之间的关系
不同氮素添加下Pn、叶片功能性状与土壤理化性质相关分析表明(表6),Pn与土壤含水量、土壤pH值呈极显著负相关(P<0.01),与土壤全N、土壤N∶P呈显著正相关(P<0.05)。 叶片SLA与土壤pH呈极显著负相关(P<0.01)。CCmass与土壤含水量呈显著负相关(P<0.05),与土壤N∶P呈显著正相关(P<0.05),与土壤pH值呈极显著负相关(P<0.01)。Pmass与含水量、土壤pH呈极显著正相关(P<0.01),与土壤全N呈极显著负相关(P<0.01)),与土壤N∶P呈显著负相关(P<0.05)。Nmass与土壤含水量呈显著负相关(P<0.05),与土壤N∶P呈显著正相关(P<0.05),与土壤pH值呈极显著负相关(P<0.01)。叶片N∶P与土壤含水量、土壤pH值呈极显著负相关(P<0.01),与土壤全N、土壤N∶P呈极显著正相关(P<0.01)。
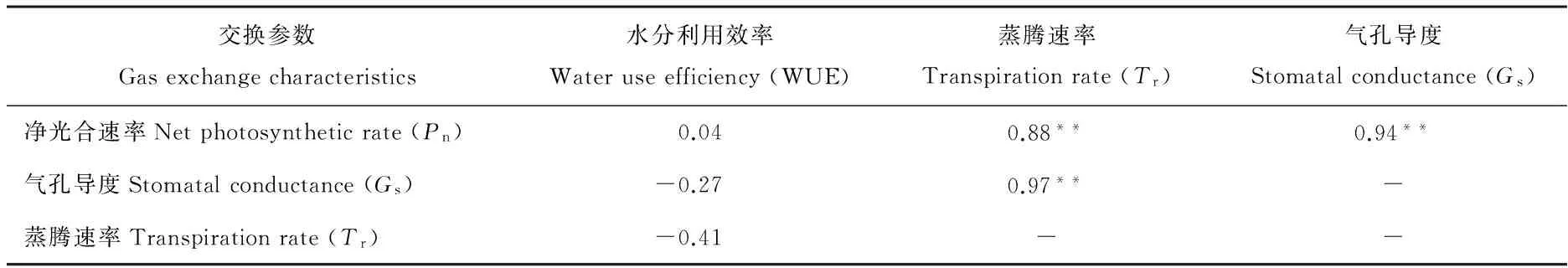
表4 不同氮素添加下贝加尔针茅气体光合参数相关性
注:*表示显著相关(P<0.05), **表示极显著相关(P<0.01)。下同。
Note: *indicated significant correlation (P<0.05), **indicated highly significant correlation (P<0.01). The same below.

表5 不同氮素添加下贝加尔针茅Pn与叶片Nmass、Pmass、SLA、CCmass、PEUE、PNUE之间的相关性

表6 不同氮素添加下Pn、叶片功能性状与土壤理化性质相关性
3 讨论与结论
氮素是植物生长需求最大量的矿质营养元素之一,大多数研究表明,氮素通常是草原生态系统中限制植物生长的最主要因子[25-26],但也有研究认为,由于氮沉降的加剧,草地生态系统可能由氮限制逐渐转向受氮、磷共同限制[27]。长期的氮肥添加实验已经证实,当生态系统中固有的氮素含量满足或者已经超过植物对氮素需求量时,过量氮输入显著降低生态系统的生产力。氮添加对生态系统生产力的影响还受土壤含水量的影响。在土壤含水量较高的情况下,适量氮添加会提高系统的生产力,在土壤含水量较低的情况下,生态系统的生产力对氮素添加的响应不明显,甚至还会出现降低的趋势[28]。在本研究中,随氮添加量增大,贝加尔针茅的Pn、Tr、Gs三者降低的幅度也越大(表2)。相关分析表明,5种氮素添加处理的Pn与Tr、Gs呈极显著正相关关系(表4)。植物在光合作用时,气孔打开并吸收CO2的同时,也伴随着蒸腾作用,本研究测定的Gs与Pn、Tr呈极显著正相关,是符合光合规律的。该研究结论与许大全[29]研究结论一致。低量添加氮素下,贝加尔针茅的Pn未出现升高现象,这与前人的研究结果“低量添加氮素提高牧草的光合能力”[30-31]有差异。供氮过高导致植物光合与生长受抑制的现象有许多报道[11]。也有研究报道,在干旱条件下氮添加对牧草光合作用无显著影响[32]。可能是研究的植物种类、氮添加量、添加年限、生境条件不同造成的。本研究认为长期氮添加使贝加尔针茅的光合作用受氮素积累和干旱的影响,从而导致光合作用受到限制。
水分和氮素是植物光合作用和生长必不可少的因子[33],PNUE是衡量植物利用氮营养和合理分配氮的能力,是氮对植物光合生产力产生影响的重要指标,WUE和PNUE可以预测植物是如何最优化地增加净光合速率的[34]。只有氮对植物生理过程起到限制作用时,植物对氮的分配才达到最大效率。当土壤供水不足时,植物通过降低Gs、Tr等生理活动降低Pn,减少自身对水分的需求[35]。本研究中,随着氮添加量的增加,土壤表层水分含量呈降低趋势(表1),贝加尔针茅的Pn、Gs和Tr呈降低趋势,WUE无一致变化(表2),Nmass呈升高趋势,而PNUE和PEUE呈降低趋势(表3)。本研究发现,长期施氮(6年)贝加尔针茅叶片SLA随着氮添加量的变化发生了较大变化,可能是因为随着氮添加量的增加,群落环境发生变化,氮可利用性和水分可利用性对植物的SLA产生了影响。这与窦晶鑫等[36]对小叶章(Calamagrostisangustifolia)叶片比叶面积对氮沉降的响应研究结果一致。相关分析表明,Nmass与PNUE和PEUE呈极显著负相关(表5)。Lambers等[37]研究表明,在植物有效光合辐射情况下,氮含量较低的叶片PNUE较大。本研究与其研究结论一致。说明随着氮添加量的增大,贝加尔针茅资源利用效率降低。
土壤养分组成是植物的外界环境中非常重要的因子,植物光合作用和叶片矿质代谢等过程与土壤养分供应状况有显著相关性。氮添加对土壤理化性质产生了影响,随着氮添加量的增大,土壤含水量、土壤pH值呈降低趋势,土壤含N量,土壤N∶P呈升高趋势(表1)。这些环境因子的改变都会导致植物光合生理特征、叶片功能性状变化。SLA与土壤全N、土壤全P、土壤N∶P呈正相关,但是这种相关性均没有达到显著水平(表6),说明SLA与土壤理化性质关系受很多因子的调控,使得这种关系变得复杂。相关分析表明,净光合速率Pn与土壤含水量、土壤全N、土壤N∶P、土壤pH均显著相关;叶片CCmass与土壤含水量、土壤N∶P、土壤pH显著相关;叶片N∶P与土壤含水量、土壤全N、土壤N∶P、土壤pH显著相关(表6)。随着氮添加量的增大,土壤含水量下降,贝加尔针茅光合能力减弱,同化产物减少,这与李林芝等[38]研究呼伦贝尔草甸草原羊草的光合特性时得到的结果是一致的。叶片的N、P含量反映了土壤N、P的有效性,本研究结果表明氮素添加增加了土壤N供应,随N素添加量的增加,土壤中的有效N供应也变得相对充足,贝加尔针茅Nmass增加,叶片N∶P由未添加氮素的15.05增加到31.79,表明长期氮添加会显著提高贝加尔针茅叶片N∶P。这与李博[14]对松嫩草原羊草的研究结论一致。大量研究表明,氮沉降可以通过改变植物对氮素、磷素的吸收,从而改变植物叶片N∶P等生态化学计量特征[6-7,15,39]。由于土壤养分的可利用性影响植物对养分的摄取,本研究中贝加尔针茅叶片N∶P表现出较大的变化范围,各个氮素处理的土壤养分供给的差异可能是引起这种变化的一个重要原因。贝加尔针茅光合作用特征、叶片功能性状变异机制随氮添加量变化而发生差异的原因还需进一步深入研究。
简而言之,长期氮添加会降低贝加尔针茅的净光合速率、气孔导度和蒸腾速率,且氮添加量越大,光合作用能力所受抑制越强烈。长期氮添加,可提高贝加尔针茅叶片氮含量、比叶面积、叶片氮磷比值、建成成本,降低养分资源的利用效率。长期氮添加引起的土壤水分含量、土壤pH改变是导致这种转变发生的主要因子。
[1] Galloway J N, Dentener F J, Capone D G,etal. Nitrogen cycles: past, present, and future. Biogeochemistry, 2004, 70(2): 152-226.
[2] Liu X J, Zhang Y, Han W X,etal. Enhanced nitrogen deposition over China. Nature, 2013, 494: 459-462.
[3] Li W J, Liu H M, Zhao J N,etal. Effects of nitrogen and water addition on plant species diversity and biomass of common species in theStipabaicalensisSteppe, Inner Mongolia, China. Acta Ecologiga Sinica, 2015, 35(19): 6460-6469.
[4] Clark C M, David T. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451: 712-715.
[5] Chalcraft D R, Cox S B, Clark C,etal. Scale-dependent responses of plant biodiversity to nitrogen enrichment. Ecology, 2008, 89(8): 2165-2171.
[6] Huang J Y, Yu H L, Yuan Z Y,etal. Effects of long-term increased soil N on leaf traits of several species in typical Inner Mongolian grassland. Acta Ecologiga Sinica, 2012, 32(5): 1419-1427.
[7] Wan H W, Yang Y, Bai S Q,etal. Variations in leaf functional traits of six species along a nitrogen addition gradient inLeymuschinensissteppe in Inner Mongolia. Journal of Plant Ecology, 2008, 32(3): 611-621.
[8] Ceulemans R, Janssens I A, Jach M E. Effects of CO2enrichment on trees and forests: lessons to be learned in view of future ecosystem studies. Annuals of Botany, 1999, 84(5): 577-590.
[9] Maroco J P, Breia E, Faria T,etal. Effects of long-term exposure to elevated CO2and N fertilization on the development of photosynthetic capacity and biomass accumulation inQuercussuberL. Plant Cell and Environment, 2002, 25(1): 105-113.
[10] Zhang X C, Shangguan Z P. The responses of photosynthetic electron transport and partition in the winter wheat leaves of different drought resistances to nitrogen levels. Plant Physiology Communications, 2009, 45(1): 13-18.
[11] Nakaji T, Fukami M, Dokiya Y,etal. Effects of high nitrogen load on growth, photosynthesis and nutrient status ofCryptomeriajaponicaandPinusdensifloraseedlings. Trees, 2001, 15(8): 453-461.
[12] Neff J C, Townsend A R, Gleixner G,etal. Variable effects of nitrogen additions on the stability and turnover of soil carbon. Nature, 2002, 419: 915-917.
[13] Guo J H, Liu X J, Zhang Y,etal. Significant acidification in major Chinese croplands. Science, 2010, 327: 1008-1010.
[14] Li B. Influence of Warming and N Deposition on Physiological and Ecological Characteristic ofLeymuschinesisin the Songnen Grassland[D]. Changchun: Northeast Normal University, 2015.
[15] Bin Z J, Wang J J, Zhang W P,etal. Effects of N addition on ecological stoichiometric characteristics in six dominant plant species of alpine meadow on the Qinghai-Xizang Plateau, China. Chinese Journal of Plant Ecology, 2014, 38(3): 231-237.
[16] Hu J Y, Zhu J X, Zhou Z,etal. Effects of experimental nitrogen additions on understory species diversity in four forests in China. Acta Scientiarum Naturalium Universitatis Pekinensis, 2014, 50(5): 904-910.
[17] Huenneke L F, Hamburg S P, Koide R,etal. Effects of soil resources on plant invasion and community structure inCalifornianserpentinegrassland. Ecology Usa, 1990, 71(2): 478-491.
[18] Zhao S, Zhang J N, Lai X,etal. Analysis of microbial biomass C, N and soil microbial community structure ofStipasteppes using PLFA at grazing and fenced in Inner Mongolia, China. Journal of Agro-Environment Science, 2011, 30(6): 1126-1134.
[19] Zhang N L, Wan S Q, Li L H. Impacts of urea N addition on soil microbial community in a semiarid temperate steppe in northern China. Plant and Soil, 2008, 311(1/2): 19-28.
[20] Deng Y, Liu X N, Xin X P,etal. Diurnal dynamics of photosynthetic characteristics ofLeymuschinensisunder different grazing intensities taking the Hulunber meadow steppe as an example. Acta Prataculturae Sinica, 2012, 21(3): 308-313.
[21] Bao S D. Soil and Agricultural Chemistry Analysis[D]. Third Edition. Beijing: China Agricultural Press, 2000.
[22] Williams K, Percival F, Merinao J,etal. Estimation of tissue construction cost from heat of combustion and organic nitrogen content. Plant, Cell & Environment, 1987, 10(9): 725-734.
[23] Vries F W, Brunsting A H, Laar H H V. Products, requirements and efficiency of biosynthesis: A quantitative approach. Journal of Theoretical Biology, 1974, 45(2): 339-377.
[24] Inclan R, Alonso R, Pujadas M,etal. Ozone and drought stress: Interactive effects on gas exchange in Aleppo pine (PinushalepensisMill). Chemosphere, 1998, 36(4/5): 685-690.
[25] Wan S, Xia J, Liu W,etal. Photosynthetic overcompensation under nocturnal warming enhances grassland carbon sequestration. Ecology, 2009, 90: 2700-2710.
[26] Bai Y F, Wu J G, Xing Q,etal. Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau. Ecology, 2008, 89(8): 2140-2153.
[27] Craine J M, Morrow C, Stock W D. Nutrient concentration ratios and co-limitation in South African grasslands. New Phytologist, 2008, 179(3): 829-836.
[28] Asner G P, Townsend A R, Riley W J,etal. Physical and biogeochemical controls over terrestrial ecosystem responses to nitrogen deposition. Biogeochemistry, 2001, 54(1): 1-39.
[29] Xu D Q. Some problems in stomatal limitation analysis of photosynthesis. Plant Physiology Communications, 1997, 33(4): 241-244.
[30] Xiao S S, Dong Y S, Qi Y C,etal. Effects of mineral fertilizer addition on leaf functional traits and photosynthetic characteristics ofLeymuschinensisfrom a temperate grassland in Inner Mongolia in China. Acta Scientiae Circumstantiae, 2010, 30(12): 2535-2543.
[31] An H, Shangguan Z P. Specific leaf area, leaf nitrogen content, and photosynthetic acclimation ofTrifoliumrepensL. seedlings grown at different irradiances and nitrogen concentrations. Photosynthetica, 2008, 46(1): 143-147.
[32] Yang H, Luo Y C. Responses of the functional traits inCleistogenessquarrosato nitrogen addition and drought. Chinese Journal of Plant Ecology, 2015, 39(1): 32-42.
[33] Novriyanti E, Watanabe M, Makoto K,etal. Photosynthetic nitrogen and water use efficiency of acacia and eucalypt seedlings as afforestation species. Photosynthetica, 2012, 50(2): 273-281.
[34] Castellanos A E, Martinez M J, Llano J M,etal. Successional trends in Sonoran desert abandoned agricultural fields in northern Mexico. Journal of Arid Environments, 2005, 60(3): 437-455.
[35] Mahajan S, Tuteja T. Cold, salinity and drought stresses: An overview. Archives of Biochemistry & Biophysics, 2005, 444(2): 139-158.
[36] Dou J X, Liu J S, Wang Y,etal. Ecophysiological responses ofCalamagrostisangustifoliato nitrogen deposition. Wetland Science, 2009, 7(1): 40-46.
[37] Lambers H, Chapin III F S, Pons T L. Plant Physiological Ecology[M]. Second Edition. New York: Springer-Verlag, 2008.
[38] Li L Z, Zhang D G, Xin X P,etal. Photosynthetic characteristics ofLeymuschinensisunder different soil moisture grades in Hulunber prairie. Acta Ecologiga Sinica, 2009, 29(10): 5271-5279.
[39] Bergstrom A K, Faithfull C, Karlsson D,etal. Nitrogen deposition and warming-effects on phytoplankton nutrient limitation in subarctic lakes. Global Change Biology, 2013, 19(8): 2557-2568.
[3] 李文娇, 刘红梅, 赵建宁, 等. 氮素和水分添加对贝加尔针茅草原植物多样性及生物量的影响. 生态学报, 2015, 35(19): 6460-6469.
[6] 黄菊莹, 余海龙, 袁志友, 等. 长期 N 添加对典型草原几个物种叶片性状的影响. 生态学报, 2012, 32(5): 1419-1427.
[7] 万宏伟, 杨阳, 白世勤, 等. 羊草草原群落 6 种植物叶片功能特性对氮素添加的响应. 植物生态学报, 2008, 32(3): 611-621.
[14] 李博. 温度升高和氮沉降对松嫩草地羊草种群生理与生态特性的影响[D]. 长春: 东北师范大学, 2015.
[15] 宾振钧, 王静静, 张文鹏, 等. 氮肥添加对青藏高原高寒草甸6个群落优势种生态化学计量学特征的影响. 植物生态学报, 2014, 38(3): 231-237.
[16] 胡钧宇, 朱剑霄, 周璋, 等. 氮添加对4种森林类型林下植物多样性的影响. 北京大学学报, 2014, 50(5): 904-910.
[18] 赵帅, 张静妮, 赖欣, 等. 放牧与围栏内蒙古针茅草原土壤微生物生物量碳、氮变化及微生物群落结构 PLFA 分析. 农业环境科学学报, 2011, 30(6): 1126-1134.
[20] 邓钰, 柳小妮, 辛晓平, 等. 不同放牧强度下羊草的光合特性日动态变化. 草业学报, 2012, 21(3): 308-313.
[21] 鲍士旦. 土壤农化分析[D]. 第三版. 北京:中国农业出版社, 2000.
[29] 许大全. 光合作用气孔限制分析中的一些问题. 植物生理学通讯, 1997, 33(4): 241-244.
[30] 肖胜生, 董云社, 齐玉春, 等. 内蒙古温带草原羊草叶片功能特性与光合特征对外源氮输入的响应. 环境科学学报, 2010, 30(12): 2535-2543.
[32] 杨浩, 罗亚晨. 糙隐子草功能性状对氮添加和干旱的响应. 植物生态学报, 2015, 39(1): 32-42.
[36] 窦晶鑫, 刘景双, 王洋, 等. 小叶章对氮沉降的生理生态响应. 湿地科学, 2009, 7(1): 40-46.
[38] 李林芝, 张德罡, 辛晓平. 等. 呼伦贝尔草甸草原不同土壤水分梯度下羊草的光合特性. 生态学报, 2009, 29(10): 5271-5279.
Effects of long-term nitrogen addition on photosynthetic characteristics and leaf traits ofStipabaicalensisin Inner Mongolia, China
LIU Hong-Mei**, LI Jie**, HUANGFU Chao-He, CHEN Xin-Wei, YANG Dian-Lin
Agro-EnvironmentalProtectionInstitute,MinistryofAgriculture,Tianjin300191,China
Atmospheric nitrogen deposition is an important component phenomenon of global change, and induces a series of ecological problems. We conducted a field manipulation experiment to study the photosynthetic response and change in leaf functional traits after long-term nitrogen addition in an S.baicalensis grassland in Inner Mongolia. This study was conducted from 2010-2015. Five treatments including N0(0 kg N/ha), N30(30 kg N/ha), N50(50 kg N/ha), N100(100 kg N/ha) and N150(150 kg N/ha) were set up and forty-eight plots, sized 8 m×8 m, were established with 2 m strips between each plot. The objective was to determine how photosynthetic characteristics and leaf traits vary with nitrogen addition and what causes these differences. Photosynthetic traits and leaf traits differed among different nitrogen addition treatments. Net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), photosynthetic nitrogen use efficiency (PNUE), photosynthetic energy use efficiency (PEUE) under N30, N50, N100, and N150treatments were lower or significantly lower than those under N0. Specific leaf area (SLA), leaf N content (Nmass), leaf construction cost (CCmass), leaf N∶P under N30, N50, N100, N150treatments tended to be higher or were significantly higher than those under N0. Correlation analysis showed thatPnwas positively correlated withGs,Tr, PNUE, PEUE, leaf P content (Pmass),Nmassand soil N, P whilePnwas negatively correlated with SLA,Nmass,CCmass, soil water content and soil pH;Nmasswas positively correlated with SLA,CCmassand soil N, P, andNmasswas negatively correlated with PNUE, PEUE,Pmass, soil water content and soil pH. In conclusion, long-term nitrogen addition induced decreases inPnand nutrient use efficiency ofS.baicalensis, and increases in leafCCmassand N∶P. Soil water content and pH decreased with nitrogen addition. This suggests that water content and soil pH are two important factors affecting photosynthetic characteristics and leaf traits of S.baicalensis under different nitrogen addition regimes.
nitrogen addition;Stipabaicalensis; photosynthetic characteristics; leaf traits
10.11686/cyxb2016110
http://cyxb.lzu.edu.cn
2016-03-14;改回日期:2016-06-06
国家自然科学基金项目(31170435)资助。
刘红梅(1976-),女,河北沧州人,副研究员。E-mail:liuhongmei@caas.cn。李洁(1986-),女,河北保定人,助理研究员。E-mail:lijie@caas.cn。**共同第一作者These authors contributed equally to this work.
刘红梅, 李洁, 皇甫超河, 陈新微, 杨殿林. 贝加尔针茅光合特征与叶片功能特性对长期氮添加的响应. 草业学报, 2016, 25(11): 76-85.
LIU Hong-Mei, LI Jie, HUANGFU Chao-He, CHEN Xin-Wei, YANG Dian-Lin. Effects of long-term nitrogen addition on photosynthetic characteristics and leaf traits ofStipabaicalensisin Inner Mongolia, China. Acta Prataculturae Sinica, 2016, 25(11): 76-85.
