我国花鲈养殖产业现状与种子工程研究进展
2016-12-02温海深张美昭李吉方
温海深,张美昭,李吉方,何 峰,李 昀
(中国海洋大学海水养殖教育部重点实验室,青岛 266003)
我国花鲈养殖产业现状与种子工程研究进展
温海深,张美昭,李吉方,何 峰,李 昀
(中国海洋大学海水养殖教育部重点实验室,青岛 266003)
花鲈(Lateolabraxmaculatus)是我国养殖产量最高的海产鱼种类之一,据中国渔业年鉴统计,2010~2014年连续五年其养殖产量超过10×104t。但是,近年来我国花鲈养殖产业正面临着空前挑战,花鲈种质退化、缺乏优良品种、病害增多;养殖成本上升、流通与加工产业滞后,活鱼价格偏低等。本文对我国花鲈种类、分布与主要生物学特征、花鲈养殖产业现状及存在的问题、我国花鲈种子工程相关研究进展进行了综述,并提出加强花鲈生殖调控与遗传基础研究,构建花鲈苗种规模化繁育基地,发展花鲈育种核心前沿技术,加快花鲈种质创新步伐等建议,以期为花鲈养殖产业可持续发展提供科学参考。
花鲈; 养殖产业; 种子工程
1 花鲈种类、分布与主要生物学特征
1.1 鲈鱼分类与地理分布
花鲈(Lateolabraxmaculatus),属于鲈形目,鮨科,花鲈属,分布于我国、日本和朝鲜半岛近海及河口地带,海水中繁殖,各类型水域均可生长,花鲈是我国重要的海产经济鱼类,在海水、半咸水、淡水或河口地区均可存活与生长。
1.2 花鲈种类与主要生物学特征
花鲈的英文名为spotted sea bass,又称中国鲈鱼、海鲈、寨花等,曾用拉丁学名为日本鲈鱼(Lateolabraxjaponicas)。Bleeker(1854~1857年)建立花鲈属Lateolabrax时,该属仅有一个物种,即Lateolabraxjaponicas[1],因此中国花鲈与日本鲈鱼长期被认为属于同一个物种。1995年日本学者提出两者在分类学角度存在差异[2],2000年中国学者也倾向于两者为不同物种[3],2006年中国学者Liu等发表论文提出了区分2个种的遗传学与分子生物学证据[4],后来花鲈Lateolabraxmaculatus逐渐得到国内外学者认同[5-8]。目前比较公认花鲈属有3个种,分别为高体鲈(L.latus)、日本鲈(L.japonicus)和花鲈(L.maculatus)。花鲈广布于中国沿岸,南到越南边界,北到朝鲜半岛西岸;日本鲈主要分布区域为除北海道外的日本沿岸水域和朝鲜半岛东南近岸水域;高体鲈则仅分布于日本静冈、长崎等南部近海海域(图1)。花鲈体侧有较多斑点(图2),日本鲈成体体侧无斑点。日本曾大量从香港、台湾、烟台等地进口中国花鲈鱼苗进行养殖,广东、浙江等地也大量利用北方苗种和亲鱼进行人工养殖和育苗,因此国内外鲈鱼种质资源比较混乱[3]。
有些学者认为在我国沿海花鲈包含北方种群和南方种群,前者主要包括黄渤海沿海和东海沿海,后者包括南海沿海和广西北海沿海,但是对具体的种群划分没有明确的定论[9-11]。国内有关花鲈生物学与生态学研究报道较多[12-18]。
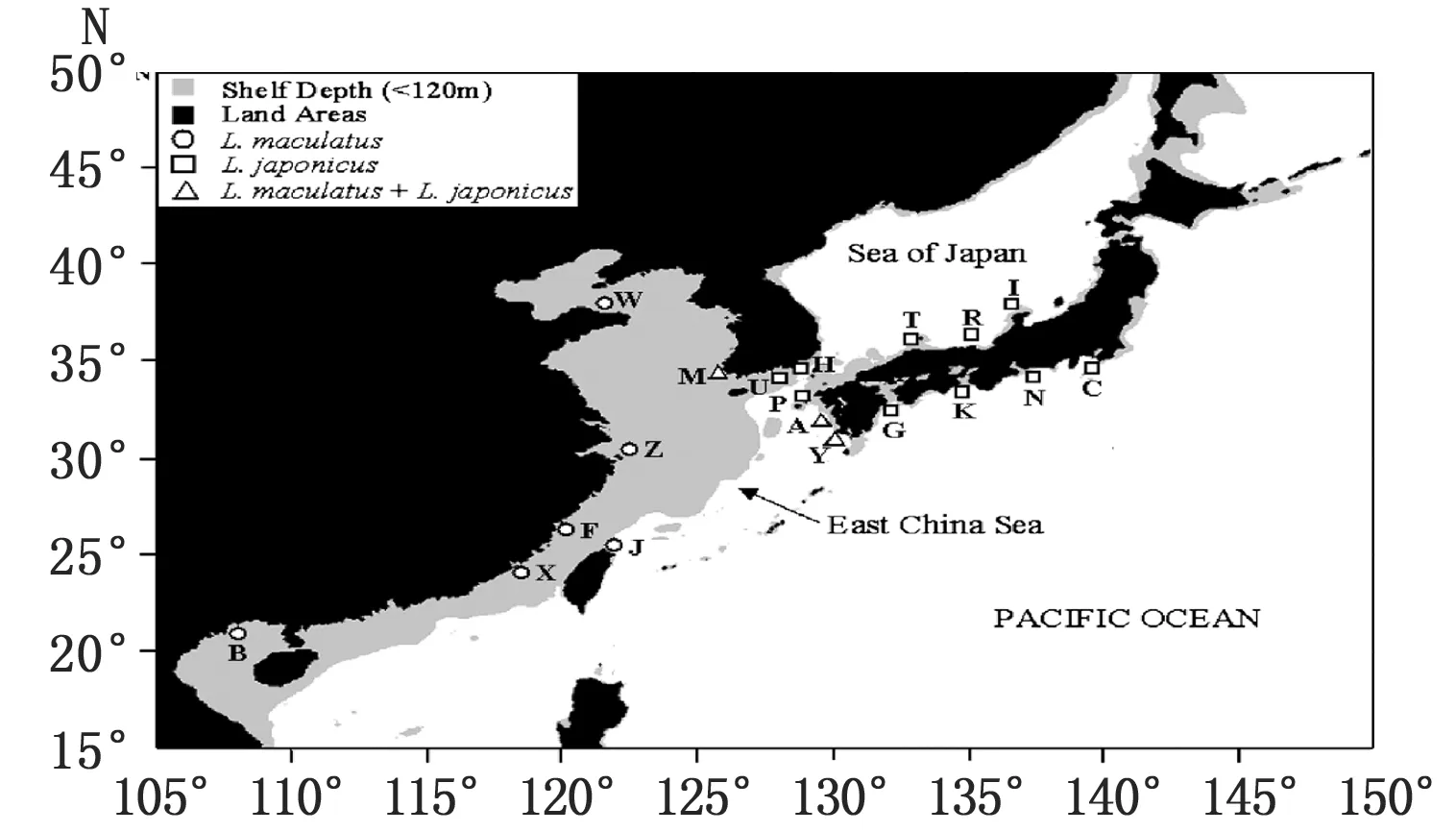
图1 花鲈属鱼类地理分布图(引自Liu 等,2006)Fig.1 The distribution map according to geographic distribution of the Lateolabrax genus (From Liu et al,2006)
2 花鲈养殖产业现状及存在的问题
2.1 花鲈养殖模式与产量
我国花鲈养殖区域较广,除了上海市外,其它10个沿海省区均有养殖,但主要集中在广东、山东、福建、浙江、广西等省区的沿海地区(图3)。目前,内陆养殖水域对花鲈苗种需求也在增加。花鲈在海水养殖鱼类中产量始终名列前茅,2012年达到12.58×104t(2013年统计数据),约为日本和韩国海鲈产量总和,年总产值近30亿元人民币。其中广东、福建、浙江、广西和山东的养殖产量均比较高。海水网箱、室内工厂化、海水池塘、内陆池塘、河口池塘和中小型水域等均适合花鲈规模化养殖。

图2 花鲈外部形态特征Fig.2 External morphological characteristics of Lateolabrax maculatus

图3 我国10省区海水养殖花鲈产量(2010~2014年)Fig.3 Production of spotted sea bass from ten provinces during 2010-2014
2.2 花鲈进出口情况
虽然全球鲈鱼类产品供不应求(中金企信,未发表数据),但我国花鲈市场已处于饱和状态,花鲈养殖企业应加大出口量以及出口渠道的开发。据不完全统计(2010),国内花鲈苗种市场需求量2亿尾,近年来基本稳定在这个规模,苗种主要需求在福建、广东和浙江等地。中国花鲈的主要出口贸易国家有:日本、韩国和新加坡,其中日本和韩国占总出口比例的90%以上。我国黄渤海花鲈种质优良,深受日本和韩国企业欢迎,他们每年从我国进口花鲈苗种约2 000多万尾。
2.3 花鲈养殖企业
2.3.1 福建省花鲈养殖业
福建闽威实业股份公司被确定为中国农业部福鼎花鲈良种场、中国农技协福建福鼎鲈鱼养殖专业技术交流中心、福建省闽威花鲈加工企业工程技术研究中心等。福建省漳州市成为鲈鱼繁育集散地,该地区有许多个体水产养殖企业从事花鲈人工繁殖、苗种培育与销售。据不完全统计,全市共有家庭式花鲈鱼苗孵化场近100家,年生产花鲈受精卵可高达1 000 kg左右。
2.3.2 广东省花鲈成鱼池塘养殖
广东省珠海市斗门区“白蕉海鲈”获“国家地理标志”保护产品称号,全市花鲈养殖农户达1 000多户,全年花鲈上市总产量占全国80%以上,形成了较完善的生产、加工、销售产业链。花鲈除了出口韩国和日本外,大部分运输到北方的山东省等地进行销售,青岛形成了全国主要的冰鲜花鲈消费市场。
2.3.3 浙江省花鲈出口产品基地
浙江奉化市宏泰水产养殖有限公司是宁波地区最大出口活鱼企业,该公司的养殖鱼场就在“中国第一渔村”奉化桐照村。该公司曾经有两个养殖基地,近5 000只网箱,为宁波地区首家“区域化管理”基地。该公司每年向韩国出口鲈鱼活鱼100 t以上,占韩国活鱼市场总额的10%以上,其中海水鲈鱼占韩国市场30%的份额。
2.3.4 山东省国家级花鲈原种场与良种场
山东威海裕隆水产开发有限公司为国家级花鲈原种场,该企业主要围绕花鲈原种保存、后备亲本培育和优质苗种生产技术等方面进行研究。山东省东营市利津县双瀛水产苗种有限责任公司主要进行开展花鲈繁育,2014年申请注册“利北花鲈”商标,该品种适合东营市盐碱水域及北部内陆盐碱地水域养殖。目前该公司与中国海洋大学共建花鲈省级良种场,实施国家科技支撑计划课题和东营市渔业科技项目,取得了较好成果。
2.4 花鲈养殖产业存在的问题
花鲈主养区养殖产业正面临着空前挑战,主要表现在种质退化、病害增多;养殖成本上升、流通与加工产业滞后,活鱼价格偏低。具体表现如下:(1)缺乏优良的花鲈品种:南北花鲈种群混杂,没有进行定向选育,尚未建立科学育种技术,导致花鲈生长和抗病性能下降;(2)缺乏大型花鲈苗种繁育企业:花鲈人工苗种主要由零散的个体生产户提供,种质质量难以保障,苗种生产处于无序状态;(3)池塘和网箱高密度养殖,对环境造成巨大压力,潜在养殖风险加大;(4)花鲈生殖调控与遗传育种等基础研究薄弱,关键技术未得到国家科技部门应有的重视,影响了花鲈养殖业的可持续发展。
2.5 花鲈养殖产业发展对策
(1) 拓宽销路:以打通流通渠道,扩大消费市场为根本,优化市场销售模式,加强冷链物流研究,扩大花鲈出口渠道和内陆水产品市场;(2)提高科技含量:增加科技经费投入,加强花鲈生殖调控与育种技术研发,确保花鲈苗种成活率、品种抗病能力和肌肉品质显著提升;(3)优化养殖模式:通过养殖品种的调整或养殖时间规划,与内陆淡水养殖优良品种进行混养,提高活鱼上市比例,维持相对稳定的养殖量或调整不同上市时间,保证合理的流通量,实现较高的养殖利润,保证产业的健康可持续发展。
随着花鲈苗种、养殖模式、饲料、流通等关键环节逐步完善,预计我国鲈鱼养殖产业将向着规模化、集约化、机械化、智能化的方向发展。
3 花鲈种子工程相关研究
种苗是海洋生物养殖产业的源头和必需物质基础,种质是海洋养殖业的核心问题。海水养殖产业“种子工程”包括“健康苗种繁殖”和“优良种质创制”两个部分,前者以实现原种和良种的人工繁殖为中心目标,包括鱼类原种开发、种子扩繁;后者则强调应用现代生物技术,不断改进和优化原种种质,提高产量和效益,是实现海水养殖业健康和可持续发展的有效保证[19]。
3.1 花鲈遗传育种研究进展
目前,关于花鲈遗传育种的系统研究尚未见报道,今后应针对我国海水和淡水养殖对花鲈良种的迫切需求,结合繁殖内分泌生理、基因组学、功能基因、遗传分子标记等研究领域,在育种技术上重点突破花鲈生理与遗传基础研究瓶颈。首要任务是提高生长率,缩短养殖周期。花鲈生殖周期4年,采用传统选择育种技术需要10年以上才能取得显著遗传进展,今后将重点结合现代基因组学信息、多性状复合选育、分子标记辅助选育等技术,提高育种进程和育种效率。其次,针对抗逆性状进行选育,使之能够更好地适应各类型水域,扩大养殖区域。第三是改善肌肉品质,提高经济附加值。
3.2 花鲈功能基因研究
对我国海水鱼类重要性状关键基因进行功能解析和开发利用一直是研究的重点。我国科学家在花鲈基因资源研发领域取得一些进展,获得了自主知识产权的功能基因20余个[20-28],涉及生长、发育与繁殖性状等(表1);进行了不同盐度条件下肝脏转录组测序分析,筛选盐度适应及渗透调节过程的关键基因,为解读花鲈盐度调控机制提供分子依据。但与其他经济鱼类相比,关于花鲈基础生理与遗传研究相对薄弱,功能基因研究成果尚未应用到渔业生产实际中。
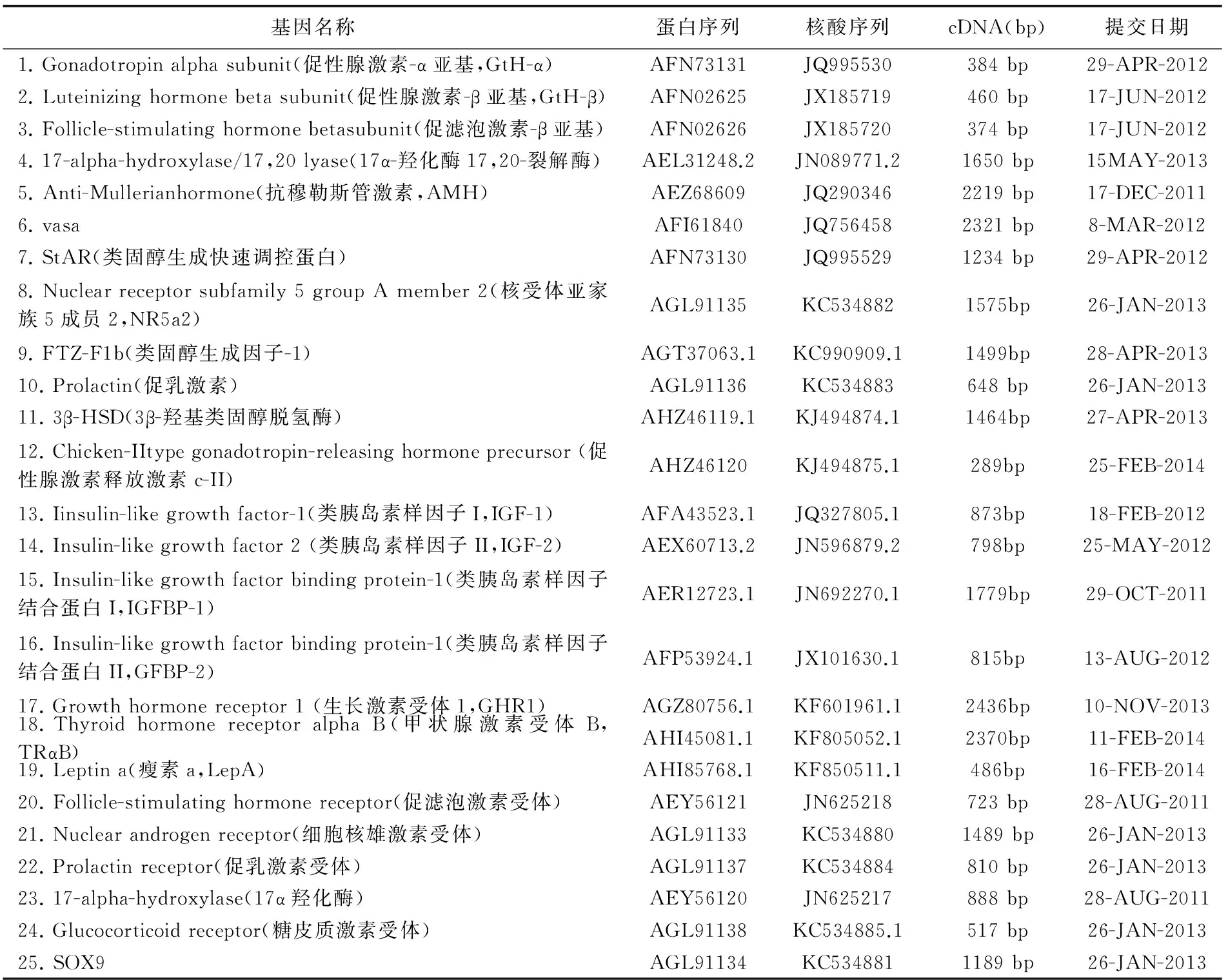
表1 花鲈生长发育与繁殖候选功能基因Tab.1 The functional genes related to growth,development and reproduction in spotted sea bass
3.3 花鲈生殖调控研究
花鲈繁殖生理与生殖调控的系统研究起步较晚,近年来侧重研究外源激素对雄性花鲈生殖内分泌机能的影响,以及不同盐度对雄性花鲈繁殖机能的影响等,取得了初步成果[23-25]。以后应侧重阐明花鲈生殖行为信号系统及转录组水平的分子机制,理解环境要素如光照、水温、盐度、营养等在促进花鲈生殖调控中的作用,建立生殖周期调控技术。
3.3.1 北部海区野生花鲈繁殖生物学比较研究
花鲈在黄渤海水域繁殖季节为9~11月,水温为16~18 ℃。生殖周期内花鲈雌鱼有2个月左右的产卵期,在此期间内分批产卵。网箱养殖条件下雌鱼和雄鱼均提早性成熟,雌鱼体重达到2.0 kg时可性成熟。绝对怀卵量30~200×104粒,相对怀卵量为 185.27~847.71粒/g体重,个体越大怀卵越多[11,29]。
3.3.2 网箱养殖花鲈繁殖生理功能评价
迟美丽[28]采集青岛沿海网箱养殖雄性花鲈150余尾,体重671.21±75.25 g,体长37.78±1.57 cm,研究网箱养殖花鲈对注射外源激素是否能产生有规律生理应答及不同盐度条件下繁殖生理功能变化,以判断能否作为后备亲鱼饲养。尽管花鲈能够在海水、淡水和半咸水中生活和生长,但是长期在淡水或半咸水中生存花鲈能否启动性腺发育,发育到什么程度,均没有实验数据支撑。实验结果提示,用于催产的后备亲鱼最好来自海水网箱养殖,不会影响激素正常分泌,淡水和半咸水中养殖的效果均不理想。在盐度不适宜的水体中血浆催乳素为硬骨鱼类渗透调节的主要激素,同时伴随血糖变化,为机体适应盐度变化提供能量,因此血糖监控也是盐度适应的主要生理指标[28]。
3.3.3 花鲈精液生理与保存
花鲈个体生殖力较大,又是分批产卵,对于精液需求量较大。由于人工养殖花鲈雄性个体均已经成熟,挤出的精液会有剩余,在这种情况下可以进行精液的超低温冷冻保存。具体操作步骤如下:将配好的稀释液与精液3:1比例装入冻存管中,摇匀后加入二甲基亚砜(抗冻剂)。逐步将冻存管放入液氮中保存:分别在液氮罐罐口、液氮面上10 cm、液氮面上6 cm、液氮面上1 cm,液氮面停留5 min左右,进行充分地预冷后再投入液氮罐中进行保存,在保存过程中要注意定时补充消耗的液氮[30]。
3.4 花鲈人工繁育研究
3.4.1 亲鱼培育
花鲈雌性亲鱼体重为3 kg以上,年龄为4龄以上,雄性亲鱼体重2 kg以上,年龄2龄以上;每年5~9月份,在室外网箱或土池塘中进行强化培养,主要投喂冰鲜鱼类,辅助以沙蚕等活饵,在北方地区人工繁殖结束后可在室内进行培育,12月下旬,北方冬季温度较低,采取升温措施,室内水温维持在12~13 ℃比较适宜[11,15]。
3.4.2 亲鱼性腺发育及调控
北方花鲈群体是每年9~11月份,水温在18~20 ℃时进行人工繁殖,此时性成熟系数最高。花鲈属于连续产卵鱼类,其性腺中生殖细胞发育不同步,自然条件下,野生花鲈只有在完全性成熟的情况下才开始排卵、排精,在人工养殖条件下,可选择一批发育程度相近的亲鱼,注射激素进行催熟和催产[31]。雌性亲鱼性腺发育状况差异较大,且成熟度鉴别比较困难。在北方海域可通过调节光周期、温度、盐度、营养等因素促进或延缓花鲈亲鱼性腺发育进程,达到反季节生产目的。
3.4.3 催产、受精及孵化
目前,南方大多数生产单位采取群体注射激素,自然产卵的方式进行繁殖,这种方法操作简单,易于管理,适合进行大规模花鲈苗种生产[12]。但是进行新品种培育和家系建设过程中,往往需要进行亲鱼催熟和催产,便于了解亲本系谱。由于花鲈雌性亲鱼存在成熟度不一致现象,如果亲鱼培养不当,造成脂肪在性腺附近积累,受精率较低。因此人工催产时进行成熟度鉴别非常关键,可通过观察腹部、挖卵鉴别卵径等方法检查成熟度,按量注射激素[12]。尽管花鲈成鱼能够在海水、半咸水、淡水中生存与生长,但是受精卵孵化必须在盐度较高的水体中进行。当盐度低于15时,孵化出的仔鱼畸形率大大提高;当盐度为25时,受精卵孵化率显著提高[17]。因此孵化过程中应当结合当地实际情况确保盐度在花鲈受精卵可接受的范围之内孵化。在水温为16~18 ℃,盐度为31时,孵化时间为56~68 h。
3.4.4 仔鱼稚鱼培养
花鲈苗种的早期培育死亡率较高,保证充足优质饵料、维持良好生活环境、维持合理密度、及时分池是保证苗种产量与质量的重要环节[11]。花鲈为肉食性鱼类,性凶猛,且具有残食性。在卵黄囊未消失时,个体发育差异较小;在仔鱼开口并自主摄食以后,随着发育进行,个体之间的发育差异开始出现,在苗种培育过程中注意及时根据发育程度进行分池,尽量避免残食现象。另外,花鲈的趋光性及争食性在苗种密度较大时也会对苗种的规模化生产造成一定的损害,及时分池可以将这些损害有效降低*此结论来自课题组其他研究成果,尚未公开发表。。
4 结论与展望
水产种业是凝聚生物高新技术最多的领域,良种的科技固化程度高,通过把复杂的高新技术成果凝聚到种子里,转化成为相对简单的技术,易被渔民接受和应用。水产良种是水产增产技术的核心,所有水产养殖技术最终依靠品种而实现其生产效益。掌握了水产良种,控制了种业,也就从顶端控制了水产养殖产业。针对花鲈已经成为当前及“十三五”期间海水养殖业发展的主要目标,开发花鲈育种核心前沿技术,加快花鲈种质创新步伐,是我国海水鱼类养殖业发展和科技竞争力提高的必然要求。
目前我国花鲈产业正面临着空前挑战,主要表现在种质退化,缺乏优良花鲈品种,北方地区仍然以捞取野生鱼苗作为苗种,导致野生花鲈资源遭到破坏,人工培育苗种成活率极低;没有进行定向选育,尚未建立科学育种技术,长期近亲繁殖导致生长和抗病性能下降;缺乏大型花鲈苗种繁育企业,花鲈人工苗种主要由零散的个体生产户提供,质量难以保障,苗种生产处于无序状态;花鲈生殖调控与遗传育种等基础研究薄弱,关键技术未得到国家科技部门的应有重视,影响了花鲈养殖业的可持续发展。未来5年必须推进花鲈养殖业良种工程建设,完善良种繁育机制,选育和推广生长快、抗逆能力强、肌肉品质好的花鲈优良品种,建立现代花鲈种子工程。
北部海区花鲈种质资源优异,是育种最好亲本来源,但是北部海域冬季水温较低增加了人工培育花鲈鱼苗的难度与成本,限制了规模化苗种生产和育种进程。主要原因是3~4个月冬季低温期造成活饵料供应难度加大,饲料成本高,生长速度慢;其次低温环境降低花鲈体质,苗种培育成活率不高,人工提高水温增加养殖成本;同时,花鲈繁殖期正值中国春节,劳动力成本偏高。可通过调控花鲈亲鱼生殖周期,延迟或提前花鲈亲鱼产卵时间来解决,但具体技术方案需要经过反复摸索才能实施;通过政府部门扶持并与大型企业合作,在北部海区构建专门的花鲈苗种规模化繁育基地或良种场,配套饵料培养、温度调控等基础设施;打通沿海与内陆花鲈苗种运输通道,建立南方与北方苗种及成鱼饲养的接力养殖模式。
[1] BLEEKER P. Nieuwe nalezingen op de ichthyologie van Japan [J]. Verh Batav Genootsch Kunst Wet, 1854-1857, 26: 1-132.
[2] YOKOGAWA K, SEKI S. Morphological and genetic differences between Japanese and Chinese sea bass of the genusLateolabrax[J]. Japanese Journal of Ichthyology, 1995, 41 (4): 437-445.
[3] 楼东, 高天翔, 张秀梅, 等. 花鲈种质资源的研究进展[J]. 浙江海洋学院学报(自然科学版), 2000, 19(2): 162-167.
[4] LIU J X, GAO T X, YOKOGAWA KOJI,etal. Differential population structuring and demographic history of two closely related Wsh species, Japanese sea bass (Lateolabraxjaponicus) and spotted sea bass (Lateolabraxmaculatus) in Northwestern Pacific[J]. Molecular Phylogenetics and Evolution, 2006, 39: 799-811.
[5] 胡自民, 高天翔, 韩志强, 等. 花鲈和鲈鱼群体的遗传分化研究[J]. 中国海洋大学学报(自然科学版), 2007, 37 (3) : 413-418
[6] 洪巧巧. 长江口中国花鲈的食性及分子生物学在食性分析上的应用[D]. 华东理工大学硕士学位论文, 2012.
[7] SUCK A H, LEE J W, KIM H Y,etal. Genetic differences between wild and hatchery populations of Korean spotted sea bass (Lateolabraxmaculatus) inferred from microsatellite markers[J]. Genes Genom, 2013, 35: 671-680
[8] 苏跃朋,等编著. 海鲈养殖新技术[M]. 北京: 中国农业出版社, 2014: 1-8.
[9] 王可玲, 张培军, 刘兰英, 等.中国近海带鱼种群生化遗传结构及其鉴别的研究[J]. 海洋学报. 1994, 16(l): 93-104.
[10] 李明云, 赵明忠, 钟爱华, 等. 山东日照和福建厦门沿海花妒同工酶的遗传变异分析[J]. 浙江海洋学院学报, 2003, 22(2): 121-124.
[11] 张美昭, 高天翔, 阮树会, 等. 花鲈亲鱼人工培育与催产技术研究[J]. 中国海洋大学学报(自然科学版), 2001, 31(2): 195-200.
[12] 雷霁霖主编. 海水鱼类养殖理论与技术[M]. 北京: 中国农业出版社, 2005: 746-749,753-754.
[13] 孙帼英, 朱云云, 周忠良, 等. 长江口及浙江沿海花鲈的繁殖生物学[J]. 水产学报, 1994, 18(1): 18- 236.
[14] 陈大刚, 高天翔, 曾晓起, 等. 莱州群体花鲈渔业生物学特征的研究[J]. 海洋学报, 2001, 23 (4): 81- 86.
[15] 张春丹, 李明云. 花鲈繁殖生物学及繁育技术研究进展[J]. 宁波大学学报(理工版), 2005, 18(3): 400-403.
[16] 胡先成, 曹双俊, 周忠良, 等. 花鲈胚胎发育的研究[J].重庆师范学院学报(自然科学版), 1997, 14(2): 51-56.
[17] 黄杰斯. 几种水环境理化因子对花鲈孵化与生长发育的影响及毒性试验研究[D]. 中国海洋大学硕士学位论文, 2015.
[18] 苏跃朋, 朱建洪, 梁健文. 广东珠海市斗门区白蕉海鲈养殖技术介绍[J]. 海洋与渔业, 2013(5): 74-76.
[19] 孟振, 刘新富, 雷霁霖. 略论我国海水鱼类苗种繁育和种质改良的研究[J]. 渔业信息与战略, 2012, 27(3): 224-231.
[20] 钱焜, 温海深, 迟美丽, 等. 花鲈类胰岛素生长因子-1基因的全长cDNA分离与表达分析[J]. 中国海洋大学学报(自然科学版), 2014, 44(2): 27-34.
[21] 钱焜, 温海深, 迟美丽, 等. 海产花鲈IGFBP-1,2 基因克隆及表达分析[J]. 中国海洋大学学报(自然科学版), 2014, 44(9): 37-45.
[22] 钱焜, 温海深, 迟美丽, 等. 外源激素对花鲈(Lateolabraxjaponicus)血清IGF-1含量及肝脏IGF-1和IGFBP-1 mRNA表达的影响[J]. 渔业科学进展, 2014, 35(4): 39-44.
[23] CHI M L, NI M, WEN H S. Molecular identification of genes involved in testicular steroid synthesis and characterization of the responses to hormones stimulation in testis of Japanese sea bass (Lateolabraxjaponicas) [J]. Steriod, 2014, 84: 92-102.
[24] CHI M L, NI M, WEN H S. Molecular cloning and characterization of gonadotropin subunits (GTHa, FSHb and LHb) and their regulation by hCG and GnRHa in Japanese sea bass (Lateolabraxjaponicas) in vivo[J]. Fish Physiology and Biochemistry, 2015, 41: 587-601.
[25] CHI M L, NI M, WEN H S. Cloning and characterization of vasa gene and expression changes analysis after hormone injection in male Japanese sea bass (Lateolabraxjaponicas)[J]. J. Ocean Univ. China, 2015, 14(4): 717-723.
[26] 张沛, 迟美丽, 温海深, 等. 盐度调控对花鲈(Lateolabraxjaponicus)生长相关基因表达的影响[J]. 海洋与湖沼, 2015, 46(2): 446-453.
[27] 张沛, 迟美丽, 温海深, 等. 花鲈瘦素基因克隆及盐度调控下的表达分析[J]. 大连海洋大学学报, 2016, 31(1): 13-18.
[28] 迟美丽. 花鲈繁殖相关基因的克隆及其在雄性个体中的表达调控分析[D]. 中国海洋大学博士学位论文, 2015.
[29] 谢凤才, 温海深. 我国花鲈人工繁育关键技术浅析[J]. 现代农业科技, 2012(20): 300,302.
[30] 柴森浩. 花鲈仔鱼发育与精液低温冷冻保存研究[D]. 中国海洋大学硕士学位论文, 2014.
[31] 温海深, 钱焜, 张美昭, 等. 花鲈繁殖生理及人工养殖模式探讨[J]. 现代农业科技, 2013 (5): 281-282.
[32] 韩枫, 温海深, 张美昭, 等. 人工繁育花鲈早期发育形态特征与仔鱼培育技术研究[J].《海洋与湖沼通报》接受待发表.
Research progress of aquaculture industry and its seed engineering in spotted sea bass (Lateolabraxmaculatus) of China
WEN Hai-shen, ZHANG Mei-zhao, LI Ji-fang, HE Feng, LI Yun
(KeyLaboratoryofMariculture,MinistryofEducation,OceanUniversityofChina,Qingdao266003,China)
Spotted sea bass (Lateolabraxmaculatus) has the highest fishery production among all marine cultured fish species in aquaculture industry in China. According to the report of China Fisheries Yearbook from 2011 to 2014, the annual production of spotted sea bass has exceeded 100,000 tons. However, this industry is suffered from several challenges including degeneration of genetic characterization, shortage of good varieties, outbreak of diseases, rising of cultivation costs, lagging behind of circulation and processing sectors and underpriced live fish. This review will focus on the distribution and biological characteristics, current situation and problems with seed engineering industry of spotted sea bass in our country, this paper would like to provide a scientific reference for sustainable development of spotted sea bass aquaculture industry. It is suggested that more efforts should be concentrated on the fundamental studies of reproductive regulation and genetic basis, construction of large-scale cultural bases, development of cutting-edge technologies for breeding and quickening the pace of germplasm innovation of spotted sea bass.
spotted sea bass (Lateolabraxmaculatus); aquaculture industry,;seed engineering.
2095-3666(2016)02-0105-07
10.13233/j.cnki.fishis.2016.02.005
2016-02-25
2016-05-03
国家“十二五”科技支撑计划重大项目课题(2011BAD13B03);东营市渔业科技项目(20150217)
温海深(1963-),男,博士,教授,研究方向:鱼类生理与繁育。E-mail:wenhaishen @ouc.edu.cn
S 917.4
A
