Manipulating extrinsic and intrinsic obstacles to axonal regeneration after spinal cord injury
2016-12-02EugeneMironets,DiWu,VeronicaJ.Tom
PERSPECTIVE
Manipulating extrinsic and intrinsic obstacles to axonal regeneration after spinal cord injury
Spinal cord injury (SCI) is a traumatic event that can lead to permanent motor and sensory deficits. After the initial trauma, axons of surviving neurons rapidly retract. While there may be a small degree of abortive sprouting, virtually all attempts at robust regrowth across the lesion site ultimately fail. Thus, neurons below the level of the injury are permanently disconnected from their normal input, resulting in persistent loss of function.
While developing central nervous system (CNS) neurons are capable of robust growth during normal development and following injury, this capacity sharply declines after a critical period due to the loss of a supportive milieu, the deposition of inhibitory matrix molecules in the glial scar, such as chondroitin sulfate proteoglycans (CSPG), by reactive astrocytes, and the presence of myelin-associated growth inhibitors, such as Nogo, myelin-associated glycoprotein (MAG), and oligodendrocyte-myelin glycoprotein (OMgp) (Silver et al., 2014). Additionally, neurons express proteins needed for growth in an age-dependent fashion - younger neurons express higher levels of these proteins than mature neurons [see Figure 1A; (Blackmore, 2012)].
This is in stark contrast to the repair response following injury to the peripheral nervous system (PNS). Dorsal root ganglion cells retain the capacity to regrow their axons after injury. Additionally, Schwann cells, the myelinating cell type in peripheral nerve, align with each other and upregulate trophic and tropic molecules, providing an environment that is much more conducive for supporting axon extension.
In the 1980’s, Albert Aguayo and colleagues re-visited pioneering work by Santiago Ramon y Cajal and his student Francisco Tello in which segments of peripheral nerve were grafted into mature brain or spinal cord. Aguayo and colleagues nicely demonstrated that some adult CNS axons do retain the limited ability to regrow if provided a suitable, supportive environment, such as grafts of growth factor-rich, pre-degenerated peripheral nerve (PNG). Unfortunately, these axons largely fail to exit the graft and re-integrate into CNS tissue because of the presence of the potent growth-inhibitors within the glial scar at the interface. We took advantage of the fact that some bacteria express an enzyme that is capable of cleaving the inhibitory moieties from CSPGs, rendering them far less growth-inhibitory. We demonstrated that treatment of the PNG-spinal cord interface with the bacterial enzyme chondroitinase (ch’ase) increases the ability of mature axons to grow across the scar (Houle et al., 2006). However, there are still many axons that grew into the graft that fail to re-emerge into spinal cord, indicating that solely addressing the environmental, scar-associated obstacles to regeneration is not optimal.
Recently, several labs have begun to elucidate what factors may mediate the intrinsic, growth deficiency in mature CNS neurons. Interestingly, increasing levels of several developmentally-regulated transcription factors, such as several members of the Krüppel-like factor (KLF) family and Sox11, in adult neurons improves their ability to re-grow or sprout. Elegant work by Zhigang He and colleagues showed that the tumor suppressor phosphatase and tensin homolog (PTEN), is upregulated in mature neurons. PTEN inhibits the activation of the protein synthesis regulator mammalian target of rapamycin (mTOR). MTOR activation increases the synthesis of proteins that are associated with growth and survival. Moreover, genetic deletion of PTEN increases mTOR activation and results in substantial axon growth after injury. However, while this approach addresses the intrinsic limitations to growth after injury, the inhibitory barriers within the glial scar remain intact.
Recently, we hypothesized that simultaneously addressing intrinsic neuronal deficits in axon regrowth and extrinsic, scar-associated impediments to regeneration would result in significant regeneration after SCI (see Figure 1B). To test this hypothesis, we used a multi-pronged approach. We grafted PN into the complete, thoracic level SCI to provide a growth-promoting environment to span the cavity. We injected adeno-associated virus (AAV) encoding for green fluorescent protein (GFP; control vector) or constitutively active Rheb (caRheb; Ras homolog enriched in brain) rostral to the lesion to transduce and drive mTOR activation in neurons above the SCI. Lastly, we treated the distal graft-host interface with ch’ase to diminish the effects of the inhibitory extracellular matrix in the glial scar (Wu et al., 2015).
Typically, very few supraspinal neurons regenerate into a PN grafted into lower level SCI sites; in such an instance, primarily propriospinal interneurons extend axons into the transplant. In our recent study, we found that expressing caRheb did not alter the population that grew into the PNG. Though caRheb was expressed in both supraspinal and propriospinal neurons rostral to the transection, axons in the PNG were mainly of propriospinal origin, though we did find that some brainstem neurons also extended into the transplants. Furthermore, as demonstrated through retrograde labeling of axons within the graft, similar numbers of neurons grew in, regardless of treatment group. Intriguingly, though, we did find significantly more fibers in the grafts in the caRheb-treated animals, suggesting that caR-heb enhances the ability of these neurons to collateralize. Along these lines, we observed more fibers from transduced neurons in cervical spinal cord, far above the thoracic level transection in the rats treated with AAV-caRheb than AAV-GFP, further indicating that caRheb expression enhances the ability to sprout.
A primary question we set out to answer was whether this combinatorial approach enhances growth across a distal graft interface. As our previous publications indicate, we found that ch’ase digestion of the scar at the distal graft interface significantly increased the numbers of axons that grew out of the PNG and into host tissue. Though we found that expressing caRheb alone (and without ch’ase) was not sufficient to improve growth across the interface, expressing caRheb did further enhance the ability of axons to cross a ch’ase-treated scar. Excitingly, in this instance, significantly more axons traversed the interface into distal cord and they extended for longer distances (see Figure 2). Furthermore, we injected the retrograde tracer Fluorogold (FG) several millimeters caudal to the lesion site to identify which and how many neurons extended these axons in caudal spinal cord. We found significantly more labeled propriospinal neurons in the caRheb+ch’ase animals than in the GFP+ch’ase animals, strongly supporting the notion that caRheb expression increases the regenerative ability of propriospinal neurons across a ch’ase-treated, thoracic spinal transection site.
A large question then is, are these newly extended axons capable of forming synapses? We saw instances that regenerated axons expressed the presynaptic marker synaptophysin, we also found examples of regenerated axons that expressed the presynaptic protein vesicular glutamate transporter 2 (VGLUT2) in extremely close proximity to post-synaptic density-95 (PSD-95), two critical components of functional glutamatergic synapses. While these data suggest that the regenerated axons formed putative synapses with appropriate pre-and post-synaptic machinery for a functional synapse, it is not known if this anatomical regeneration has functional significance. We and others have published that ch’ase-improved axon regeneration improves function (Tom et al., 2009; Alilain et al., 2011). We are currently investigating if enhancing the intrinisic growth potential in addition to ch’ase further promotes functional recovery.
Understanding the mechanism for axon growth through a graft has become increasingly important.Though activation of the mTOR pathway is thought to enhance axon growth via protein synthesis, which specific proteins may mediate this has not been well studied. It is known that growth-associated protein-43 (GAP-43) is heavily correlated with axon extension. We found that neurons expressing caRheb had higher levels of expression of GAP-43 than neurons expressing just GFP. This finding elucidates a protein that may actually underlie better growth capacity in these mature neurons. Identifying what other growth-associated proteins may mediate enhanced growth will be crucial moving forward.
In our study, we found that propriospinal neurons were the ones that appeared to respond the most robustly to caRheb expression. This may be due to the fact that these neurons have a greater propensity to grow into the graft, given their proximity to the lesion; the distance a neuron is from the lesion is inversely correlated with its ability to regenerate axons into grafts. Thus, activating mTOR may enhance a growth response only in those neurons that coincidentally express other factors needed for growth. It may not be sufficient to enhance long-distance, regenerative growth of neurons whose cell bodies are further from the lesion site (i.e., neurons within the brain). A variety of transcription factors and related proteins in addition to mTOR may need to be targeted, in this instance (Sun et al., 2011).
Because the effects of mTOR on growth is thought to be protein-synthesis dependent, a limiting factor to this approach is mRNA availability - if transcripts for growth-associated proteins are not present in the first place, there will be translation, regardless of whether protein syntheticmachinery is activated or not. Thus, an important area of investigation for future studies will need to focus on epigenetic regulation of proteins necessary for growth.
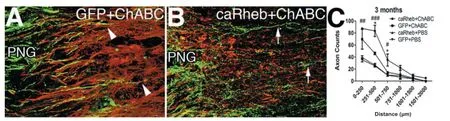
Figure 2 Expression of caRheb enhances axonal regeneration beyond a ChABC-treated PNG interface into host spinal cord tissue.

Figure 1 Extrinsic and intrinsic therapeutic targets after spinal cord injury (SCI).
Another aspect to consider when discussing functional regeneration is that since descending tracts that originate relatively far from a spinal cord injury at any level, getting these tracts to regrow back to its original target is a daunting task - not only because of the distance that the axons may need to traverse but also because guidance cues present during development to help target axons to appropriate targets are largely absent in adulthood. Thus, when devising a treatment strategy, it becomes apparent that we need to better understand the capabilities of the remaining circuitry and how to promote plasticity to tap into it. Interestingly, in an incomplete injury setting, injured axons that are disconnected from their original target can spontaneously sprout rostral to the injury and synapse upon a new target - propriospinal neurons whose projections caudal to the injury were spared. These interneurons can then relay the descending signals to neurons below the lesion to possibly result in functional recovery (Courtine et al., 2008). Because we and others have demonstrated that activating mTOR enhances axons’ intrinsic ability to sprout, this may be one way to enhance the formation of a compensatory relay.
Because multiple facets will need to be considered to achieve functional repair of the SCI, combinatorial strategies focused on manipulating intrinsic neuronal growth potential while simultaneously reducing the extrinsic inhibitors of axon extension will likely be a staple for future therapies in SCI.
This work was funded by NIH R01 NS085426 (VJT) and the Craig H. Neilsen Foundation (DW).
Eugene Mironets, Di Wu, Veronica J. Tom*
Department of Neurobiology and Anatomy, Drexel University College of Medicine, Philadelphia, Pennsylvania, PA, USA
*Correspondence to: Veronica J. Tom, Ph.D., veronica.tom@drexelmed.edu.
Accepted: 2015-12-14
orcid: 0000-0002-3369-7575 (Veronica J. Tom)
Alilain WJ, Horn KP, Hu H, Dick TE, Silver J (2011) Functional regeneration of respiratory pathways after spinal cord injury. Nature 475:196-200.
Blackmore MG (2012) Molecular control of axon growth: insights from comparative gene profiling and high-throughput screening. Int Rev Neurobiol 105:39-70.
Courtine G, Song B, Roy RR, Zhong H, Herrmann JE, Ao Y, Qi J, Edgerton VR, Sofroniew MV (2008) Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat Med 14:69-74.
Houle JD, Tom VJ, Mayes D, Wagoner G, Phillips N, Silver J (2006) Combining an autologous peripheral nervous system “bridge” and matrix modification by chondroitinase allows robust, functional regeneration beyond a hemisection lesion of the adult rat spinal cord. J Neurosci 26:7405-7415.
Silver J, Schwab ME, Popovich PG (2014) Central nervous system regenerative failure: role of oligodendrocytes, astrocytes, and microglia. Cold Spring Harb Perspect Biol 7:a020602.
Sun F, Park KK, Belin S, Wang D, Lu T, Chen G, Zhang K, Yeung C, Feng G, Yankner BA, He Z (2011) Sustained axon regeneration induced by co-deletion of PTEN and SOCS3. Nature 480:372-375.
Tom VJ, Sandrow-Feinberg HR, Miller K, Santi L, Connors T, Lemay MA, Houle JD (2009) Combining peripheral nerve grafts and chondroitinase promotes functional axonal regeneration in the chronically injured spinal cord. J Neurosci 29:14881-14890.
Wu D, Klaw MC, Connors T, Kholodilov N, Burke RE, Tom VJ (2015) Expressing constitutively active rheb in adult neurons after a complete spinal cord injury enhances axonal regeneration beyond a chondroitinase-treated glial scar. J Neurosci 35:11068-11080.
10.4103/1673-5374.177719 http://www.nrronline.org/
How to cite this article: Mironets E, Wu D, Tom VJ (2016) Manipulating extrinsic and intrinsic obstacles to axonal regeneration after spinal cord injury. Neural Regen Res 11(2):224-225.
杂志排行

中国神经再生研究(英文版)的其它文章
- Tissue-type plasminogen activator is a modulator of the synaptic vesicle cycle
- Impaired consciousness caused by injury of the lower ascending reticular activating system: evaluation by diffusion tensor tractography
- Considering calcium-binding proteins in invertebrates: multi-functional proteins that shape neuronal growth
- Cardiovascular dysfunction following spinal cord injury
- Practical application of the neuroregenerative properties of ketamine: real world treatment experience
- Exergames: neuroplastic hypothesis about cognitive improvement and biological effects on physical function of institutionalized older persons