Tissue-type plasminogen activator is a modulator of the synaptic vesicle cycle
2016-12-02ManuelYepes
PERSPECTIVE
Tissue-type plasminogen activator is a modulator of the synaptic vesicle cycle
Tissue-type plasminogen activator (tPA) is a serine proteinase that catalyzes the conversion of plasminogen into plasmin. For a long time after its discovery it was thought that tPA’s main source was the endothelial cell and that therefore its chief role was to generate plasmin in the vascular system. However, this concept was challenged four decades later when a group of investigators detected tPA mRNA in well-defined areas of the brain, namely the hippocampus, hipothalamus, thalamus, neocortex and cerebellum; and tPA-catalyzed proteolysis in the hippocampus, the amygdala, the thalamus and the hypothalamus (Sappino et al., 1993). Remarkably, subsequent work indicated that neurons but not endothelial cells are the main reservoir of tPA in the central nervous system, and that in the brain tPA has many functions that do not require plasminogen as a substrate (Yepes et al., 2003). Interestingly, while most of the studies on neuronal tPA have been performed either with cells cultured from the hippocampus or with hippocampal slices, fewer have investigated whether tPA has a function in cerebral cortical neurons (Wu et al., 2012, 2015).
Research from our laboratory has found that in the cerebral cortex tPA is expressed mainly in axons from clusters of parvalbumin-positive neurons, and that membrane depolarization induces the rapid release of this tPA (Echeverry et al., 2010). These observations are of particular importance because parvalbumin is found predominantly in interneurons that play a central role integrating information from different areas of the brain and modulating circuit function (Kaiser et al., 2015). To further characterize the axonal expression of tPA, wild-type cerebral cortical neurons were co-stained with antibodies against tPA and bassoon, a protein abundantly expressed in the presynaptic terminal. These studies revealed that approximately one-third of all axonal tPA is found in the axonal bouton (Wu et al., 2015). Remarkably, this proportion remained constant throughout all examined axons, suggesting the presence of a mechanism that regulates the translocation of tPA from the axonal shaft into the presynaptic terminal.
In the presynaptic terminal of chemical synapses neurotransmitters are stored in synaptic vesicles of ~40 nm of diameter that participate in a cycle that permits their repeated use during sustained synaptic activity. Synaptic vesicles are distributed in three groups known as readily releasable, recycling, and reserve pools (Rizzoli and Betz, 2004). The synaptic vesicles of the readily releasable pool are docked to an electron dense thickening of the presynaptic membrane, known as the active zone, where exocytosis takes place. In contrast, synaptic vesicles from the recycling and reserve pools are not docked to the active zone and to release their load of neurotransmitters they need to translocate to the readily releasable pool. The readily releasable pool is rapidly depleted following membrane depolarization. Hence, the mobilization of synaptic vesicles from the recycling and reserve pools to replenish the readily releasable pool is of pivotal importance to maintain neurotransmitter release during sustained synaptic activity.
To study the expression of tPA in the presynaptic terminal, we prepared synaptoneurosomes from wild-type cerebral cortical neurons and subjected them to sucrose density fractionation to isolate the synapse assembled by the presynaptic membrane, synaptic vesicles docked to the active zone, and the attached post-synaptic density of the post-synaptic membrane. Then each gradient fraction was immunoblotted with antibodies against tPA, synaptophysin (an integral transmembrane protein found in synaptic vesicles), syntaxin I (a transmembrane protein found in the presynaptic plasma membrane), and post-synaptic protein-95 (detects the post-synaptic density). Our data indicate that in the presynaptic terminal tPA is stored outside the active zone and in line with these observations, electron microscopy studies detected tPA-containing vesicles either intermixed with small clear-core vesicles, or in direct contact with the presynaptic membrane, but always outside the active zone. This finding is of significant importance because it indicates that tPA is released at extra-synaptic sites and suggest that as it has been described for presynaptic neuropetides released outside the active zone, tPA may also be able to regulate synaptic function in a large number of neurons, even those located at a long-distance within the brain. More importantly, because the presynaptic release of these neuropeptides is proportional to the magnitude of the stimulus, it is plausible to postulate that the presynaptic release of tPA is a mechanism that matches the intensity of the depolarizing stimulus with a proportional release of excitatory neurotransmitters.
To study whether the release of tPA has an effect on the presynaptic terminal we performed mass spectrometry in preparations from the active zone isolated from wild-type cerebral cortical neurons previously treated with tPA. Surprisingly, we found that tPA increases the abundance of the cytoskeletal protein βII-spectrin in the active zone. Additionally, our Western blot analyses not only confirmed these observations but also indicated that this effect does not require tPA’s ability to catalyze the conversion of plasminogen into plasmin.
Spectrin, a cytoskeletal protein critical for membrane structural integrity, is a heterotetramer composed of two α and two β subunits (Bennett and Lambert, 1991). It was first discovered in erythrocytes where it plays a pivotal role maintaining the shape and flexibility of the cell membrane by forming complexes with actin and other intracellular proteins. Subsequent studies indicated that one member of the spectrin family, βII-spectrin, is found in the presynaptic terminal (Phillips et al., 2001) where it plays an important role in synaptic transmission (Sikorski et al., 2000). Our studies indicate that tPA induces the recruitment of βII-spectrin to the active zone of cerebral cortical neurons, and that by doing so it also increases the size of the synaptic release site.
Synapsin I is a member of the synapsin family of phosphoproteins that plays a central role in clustering synaptic vesicles of the reserve pool. Accordingly, synapsin I is associated with the cytoplasmic surface of synaptic vesicles where it serves as a linker with other synaptic vesicles and the actin cytoskeleton of the presynaptic terminal. However, following membrane depolarization synapsin I is phosphorylated in a Ca2+-dependent manner, leading to its dissociation from synaptic vesicles, which then are free to translocate to the active zone to release their content of neurotransmitters into the synaptic cleft (Chi et al., 2001). βII-spectrin has a synapsin I-binding site (Sikorski et al., 1991); therefore it is plausible to postulate that tPA not only induces the recruitment of βII-spectrin to the active zone but also promotes its binding to synapsin I-expressing synaptic vesicles. This hypothesis was corroborated by our immunoprecipitation studies with synapse-enriched fractions from cerebral cortical neurons treated with tPA. In summary, our data suggest a model where the presynaptic release of tPA induces the recruitment of βII-spectrin to the active zone and by doing so not only enlarges its size but also promotes βII-spectrin binding to synaptic vesicles.
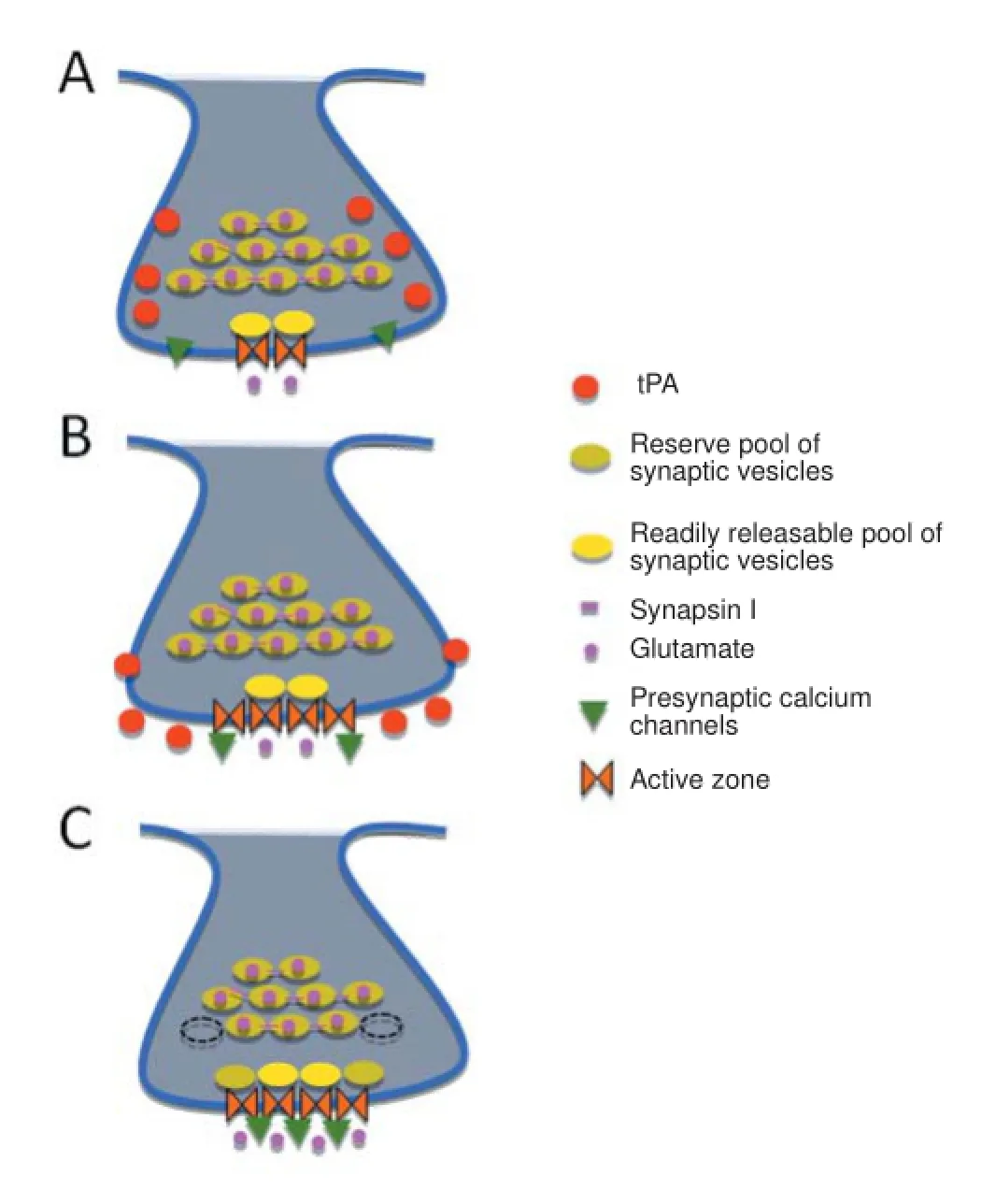
Figure 1 Tissue-type plasminogen activator activates the synaptic vesicle cycle.
Under resting conditions synapsin I clusters synaptic vesicles and tethers them to the cytoskeleton. However, during synaptic activity synapsin I phosphorylation at Serine 9 leads to its dissociation from synaptic vesicles of the reserve pool freeing them to move to the active zone. Interestingly, phosphorylation of the enzyme glycogen synthase kinase-3β at Serine 9 reduces its activity (Grimes and Jope, 2001) and this has been proposed to induce the clustering of synapsin I in the presynaptic terminal and the development of synaptic plasticity (Zhu et al., 2007). Data from our laboratory indicate that tPA induces the rapid phosphorylation of synapsin I at Serine 9 and our electrophysiology studies demonstrate that this is followed by an increase in the frequency of miniature excitatory postsynaptic currents, a postsynaptic marker of the quantal release of neurotransmiters from the presynaptic terminal.
In summary, the available data indicate that tPA plays a central role as a regulator of synaptic function. According to this model, the release of tPA from cerebral cortical neurons has a neuromodulatory effect on presynaptic function by inducing the anatomical and biochemical changes in the synaptic release site required for the release of excitatory neurotransmitters (Figure 1). Further studies are needed to understand whether this effect requires a receptor for tPA on the surface of the presynaptic terminal. Health Grants NS-079331(to MY) and NS-091201 (to MY).
Manuel Yepes*
Department of Neurology & Center for Neurodegenerative Disease, Emory University School of Medicine, Atlanta, GA, USA; Department of Neurology, Veterans Affairs Medical Center, Atlanta, GA, USA
*Correspondence to: Manuel Yepes, M.D., myepes@emory.edu.
Accepted: 2015-11-23
Bennett V, Lambert S (1991) The spectrin skeleton: from red cells to brain. J Clin Invest 87:1483-1489.
Chi P, Greengard P, Ryan TA (2001) Synapsin dispersion and reclustering during synaptic activity. Nat Neurosci 4:1187-1193.
Echeverry R, Wu J, Haile WB, Guzman J, Yepes M (2010) Tissue-type plasminogen activator is a neuroprotectant in the mouse hippocampus. J Clin Invest 120:2194-2205.
Grimes CA, Jope RS (2001) The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling. Prog Neurobiol 65:391-426.
Kaiser T, Ting JT, Monteiro P, Feng G (2015) Transgenic labeling of parvalbumin-expressing neurons with tdTomato. Neuroscience doi: 10.1016/j.neuroscience.2015.08.036.
Phillips GR, Huang JK, Wang Y, Tanaka H, Shapiro L, Zhang W, Shan WS, Arndt K, Frank M, Gordon RE, Gawinowicz MA, Zhao Y, Colman DR (2001) The presynaptic particle web: ultrastructure, composition, dissolution, and reconstitution. Neuron 32:63-77.
Rizzoli SO, Betz WJ (2004) The structural organization of the readily releasable pool of synaptic vesicles. Science 303:2037-2039.
Sappino AP, Madani R, Huarte J, Belin D, Kiss JZ, Wohlwend A, Vassalli JD (1993) Extracellular proteolysis in the adult murine brain. J Clin Invest 92:679-685.
Sikorski AF, Terlecki G, Zagon IS, Goodman SR (1991) Synapsin I-mediated interaction of brain spectrin with synaptic vesicles. J Cell Biol 114:313-318.
Sikorski AF, Sangerman J, Goodman SR, Critz SD (2000) Spectrin (betaSpIIsigma1) is an essential component of synaptic transmission. Brain Res 852:161-166.
Wu F, Wu J, Nicholson AD, Echeverry R, Haile WB, Catano M, An J, Lee AK, Duong D, Dammer EB, Seyfried NT, Tong FC, Votaw JR, Medcalf RL, Yepes M (2012) Tissue-type plasminogen activator regulates the neuronal uptake of glucose in the ischemic brain. J Neurosci 32:9848-9858.
Wu F, Torre E, Cuellar-Giraldo D, Cheng L, Yi H, Bichler EK, Garcia PS, Yepes M (2015) Tissue-type plasminogen activator triggers the synaptic vesicle cycle in cerebral cortical neurons. J Cereb Blood Flow Metab doi: 10.1038/ jcbfm.2015.155.
Yepes M, Sandkvist M, Moore EG, Bugge TH, Strickland DK, Lawrence DA (2003) Tissue-type plasminogen activator induces opening of the bloodbrain barrier via the LDL receptor-related protein. J Clin Invest 112:1533-1540.
Zhu LQ, Wang SH, Liu D, Yin YY, Tian Q, Wang XC, Wang Q, Chen JG, Wang JZ (2007) Activation of glycogen synthase kinase-3 inhibits long-term potentiation with synapse-associated impairments. J Neurosci 27:12211-12220.
This work has been supported in part by National Institutes of
10.4103/1673-5374.177712 http://www.nrronline.org/
How to cite this article: Yepes M (2016) Tissue-type plasminogen activator is a modulator of the synaptic vesicle cycle. Neural Regen Res 11(2):212-213.
杂志排行

中国神经再生研究(英文版)的其它文章
- Gas6-Tyro3 signaling is required for Schwann cell myelination and possible remyelination
- Impaired consciousness caused by injury of the lower ascending reticular activating system: evaluation by diffusion tensor tractography
- Considering calcium-binding proteins in invertebrates: multi-functional proteins that shape neuronal growth
- Cardiovascular dysfunction following spinal cord injury
- Practical application of the neuroregenerative properties of ketamine: real world treatment experience
- Exergames: neuroplastic hypothesis about cognitive improvement and biological effects on physical function of institutionalized older persons