New insight into curcumin-based therapy in spinal cord injuries: CISD2 regulation
2016-12-02Chai-ChingLin,Muh-ShiLin
PERSPECTIVE
New insight into curcumin-based therapy in spinal cord injuries: CISD2 regulation
Multiple protective effects of curcumin in cases of spinal cord injuries (SCIs): Curcumin [1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione] is a nonsteroidal, naturally occurring compound commonly utilized as a dietary pigment as well as a spice in India. It is obtained from curcuma longa in the Ginger family. Due to its anti-inflammatory, anti-oxidant, and immunomodulatory activities, this extract has been used to treat a number of inflammatory and neurodegenerative conditions, including Alzheimer’s disease, multiple sclerosis, and trauma to the central nervous system (Menon and Sudheer, 2007). Curcumin antagonizes several processes involved in the inflammatory cascade, including the activation of nuclear factor kappa B (NF-κB) and inducible nitric oxide synthase (iNOS) (Weber et al., 2006). It has also been shown to scavenge NO, decrease lipid peroxidation, and attenuate mitochondrial dysfunction (Wang et al., 2005).
Previous studies have shown that curcumin exerts protective effects against neuroinflammation in SCI rats and improves functional outcomes through the inhibition of astrocyte reactivation and the prevention of apoptosis and neuronal cell death (Lin et al., 2011a). In primary astrocyte cultures, curcumin has been shown to decrease the formation of glial fibrillary acid protein, which suggests the ability to attenuate glial activation. The anti-inflammatory effects of curcumin (glial deactivation) can be attributed to the protective benefits for neurons. Specifically, the anti-inflammatory effects of curcumin are associated with the regulation of astrocyte reactivation, particularly in cases of acute SCI with injury-induced RANTES (Regulated on Activation, Normal T Cell Expressed and Secreted) from astrocytes. The neuroprotective effects of curcumin have been demonstrated in a rat SCI model as well as in an astrocyte culture model via a reduction in RANTES production by injury-reactivated astrocytes (Lin et al., 2011b). Our research team has demonstrated novel findings that pharmacological effects of curcumin include the upregulation of CDGSH Iron Sulfur Domain 2 (CISD2) associated with SCI (Lin et al., 2015).
CISD2 is involved in the pathological response associated with SCIs: CDGSH iron sulfur domain 2 (CISD2) (synonyms: NAF-1, Eris, Miner1 and Noxp70) was originally regarded as a survival gene, based on its roles in calcium metabolism, anti-apoptosis, and longevity. Specifically, CISD2 enhances BCL2-BECN1 interaction and the binding of CISD2 to BCL2 regulates autophagy/ apoptosis in response to stress (Chang et al., 2010). Furthermore, CISD2 has been shown to inhibit an increase in excitotoxic Ca2+at the endoplasmic reticulum, via a combination between BCL2 and the inositol 1,4,5-triphosphate receptor (Chang et al., 2012). CISD2 helps to prevent mitochondrial dysfunction and subsequent cell death (Chen et al., 2009).
The authors previously identified the injury-induced down-regulation of CISD2 expression in an animal model of SCI as well as in a cell culture model of astrocyte reactivation (Lin et al., 2015). Using primary impact and hemisection to adjust injury severity, animals subjected to SCI presented a significant upregulation of proinflammatory mediators, including iNOS and RANTES as well as a downregulation of CISD2 mRNA and protein expression. In the cell culture model, primary cultured astrocytes were challenged using lipopolysaccharide (LPS) in order to mimic the conditions of glial reactivation following injury in vivo. CISD2 mRNA expression levels were significantly lower in the 1 μg/ mL LPS-challenged group than in the control group. It has been postulated that SCI causes inflammatory responses and a drop in CISD2 levels. Furthermore, CISD2 inactivity has been proposed as a mechanism underlying secondary damage following SCI.
Acute SCI has been shown to prevent the expression of CISD2 and induce neuroinflammation in vivo. This begs the question as to what reaction occurs in the signaling cascade following the down-regulation of CISD2. Answering this question requires an understanding of the physiological role of CISD2 and its association with secondary damage following SCI. To confirm a causal relationship between CISD2 and injury-induced secondary insults, we performed a loss-of-function study using siRNA to knockdown the expression of CISD2 in a SH-SY5Y cell culture of LPS-induced injury. A loss of CISD2 function caused by siCISD2 resulted in a pronounced increase in iNOS as well as a decrease in BCL2 expression. We postulate that CISD2 serves a novel role in the suppression of NO signaling. A deficiency of CISD2 increases iNOS expression, which may elevate the production of NO and the neurotoxic nitration of proteins. The subsequent attenuation of BCL2 expression promotes neuronal apoptosis.
Our research team has determined that the inactivation of CISD2 expression increases NO production in LPS-challenged SH-SY5Y cells (Lin et al., 2015). Injury-induced excessive production of NO promotes tissue damage associated with SCI and contributes to apoptotic cell death. The novel role of CISD2 in the antagonism of NO signaling is beneficial to neuronal survival. The presence of NO in glia cells may have proinflammatory effects resulting from the stimulation of proinflammatory cytokine release. Injury-activated astrocytes and microglias produce NO and reactive oxygen species (ROS), which can amplify a vicious inflammatory cascade. The presence of NO in neurons may also be an indication of longterm potentiation (LTP), an increase in neurotransmitter release, an increase in ROS formation, and the negation of anti-apoptotic BCL-2 protein. It has been surmised that CISD2 may suppress NO signaling and prevents apoptosis in neuronal cells. It has also been postulated that CISD2 serves as a neuroinflammatory suppressor in glial cells. iNOS-elevation may be associated with astrocytes and microglia following a decrease in CISD2 after SCI. This putative mechanism in glia cells has yet to be investigated. Figure 1 presents a schematic illustration of the proposed sequence of events associated with CISD2 gene expression in SCI.
Curcumin attenuates the downregulation of CISD2 in SCI animals and LPS-treated astrocytes: The authors have found that SCI downregulates CISD2, the effects of which can be prevented by curcumin (Lin et al., 2015). In an animal model of SCI as well as in a cell culture model of injury-induced astrocyte reactivation, curcumin has been shown to confer considerable benefits by attenuating the downregulation of CISD2 expression following injury. Among curcumin-treated animals, those in the SCI group presented a significant increase in CISD2 expression and a significant decrease in RANTES and iNOS mRNA expression. Animals with SCI that underwent treatment with curcumin showed increased the protein levels of CISD2 and a significantly higher percentage of CISD2 cells in the injured region of the spinal cord than did the untreated SCI group.
In primary cultures of astrocytes, CISD2 expression levels in the 1 μg/mL LPS-challenged group were significantly lower than those in the control group. LPS-reactivated cells treated with 1 μM curcumin had CISD2 expression levels far higher than those in the LPS-challenged group without curcumin treatment. in vivo and in vitro analyses indicate that the administration of curcumin led to an upregulation of CISD2 expression and a decrease in the expression of inflammatory mediators following SCI.
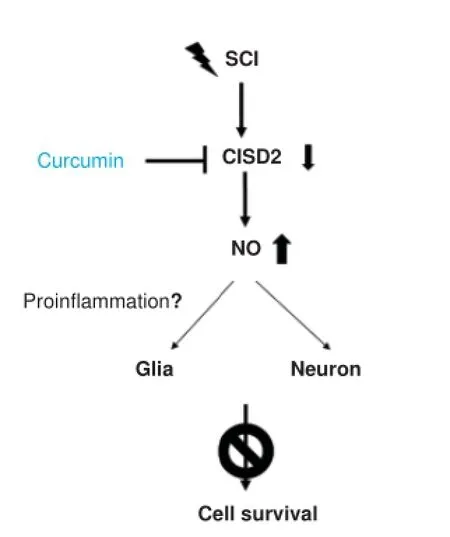
Figure 1 Downregulation of CISD2 in cases of spinal cord injury (SCI) leading to an increase in nitric oxide (NO) production.
Curcumin has been shown to suppress interleukin-1 beta (IL-1β) and tumor necrosis factor alpha (TNF-α) through the inhibition of NF-κB. NF-κB transcription factors regulate iNOS gene expression as well as a number of proinflammatory genes. It is likely that curcumin attenuates NF-κB activation and the expression of proinflammatory mediators such as iNOS, thereby decreasing glial reactivation and quenching neuronal apoptosis associated with injury-induced neuroinflammation. It appears that the upregulation of CISD2 following the administration of curcumin helps to attenuate the loss of CISD2 expressing cells as well as the direct effects related to the targeting of CISD2 by curcumin.
Curcumin-based therapy in CISD2 regulation - a potential therapeutic target in SCI-induced neuroinflammation and apoptosis: The therapeutic target of SCI is the amelioration of secondary damage to the spinal cord following traumatic SCI, which may otherwise lead to long-term disability. Methylprednisolone is the only recognized therapy widely used in the treatment of SCI. Unfortunately, the side effects of this treatment can be severe, including gastrointestinal bleeding, gastritis, and Cushing’s syndrome. Numerous pharmacological therapies for SCI have been evaluated in clinical trials; however, the therapeutic benefits remain inconclusive.
The survival-enhancing effects of CISD2 and its neuroprotective role in SCI are well established. CISD2 has been shown to exert anti-apoptotic and anti-inflammatory effects in neural cells (Lin et al., 2015). A persistent increase in the expression of CISD2 was found to improve survival rates in an in vivo transgenic mouse model (Wu et al., 2012). Thus, we surmise that any treatment modality capable of enhancing the production of CISD2 as a means of decreasing NO production and reducing neuroinflammation/ apoptosis would represent a considerable advancement in SCI therapy.
Curcumin exhibits multiple pharmacologic effects, such as anti-inflammatory, anticarcinogenic, anti-infection, antioxidant, and hypocholesterolemic activities. The neuroprotective benefits stemming from the anti-inflammatory effects of curcumin have been demonstrated in cases of SCI (Lin et al., 2011b; Lin et al., 2011a). The pharmacological effects of curcumin clearly involve the regulation of CISD2. Briefly, injury to the spinal cord downregulates the expression of CISD2; however, curcumin has been shown to attenuate the downregulation of CISD2 in SCI and LPS-treated astrocytes (Lin et al., 2015). The ability of curcumin to prolong the expression of CISD2 could be of therapeutic value in SCI therapy. The anti-apoptosis and attenuation of NO production by CISD2 may account for the neuroprotective effects of curcumin in the treatment of SCI.
This work was supported by grants from the Taipei Institute of Pathology, Taiwan (TIP10102A), the Taipei City Hospital, Taiwan (TPCH-102-061 and TPCH-104-043) and the Department of Health, Taipei City Government (10401-62-038).
Chai-Ching Lin, Muh-Shi Lin*
Department of Neurosurgery, Taipei City Hospital, Zhong Xiao Branch, Taipei, Taiwan, China (Lin MS)
Department of Surgery, Faculty of Medicine, School of Medicine, National Yang-Ming University, Taipei, Taiwan, China (Lin MS) Department of Biotechnology and Animal Science, College of Bioresources, National Ilan University, Yilan, Taiwan, China (Lin CC, Lin MS)
*Correspondence to: Muh-Shi Lin, M.D., Ph.D., LL.M., neurosurgery2005@yahoo.com.tw.
Accepted: 2015-12-25
orcid: 0000-0002-9798-8160 (Muh-Shi Lin)
Chang NC, Nguyen M, Shore GC (2012) BCL2-CISD2: An ER complex at the nexus of autophagy and calcium homeostasis? Autophagy 8:856-857.
Chang NC, Nguyen M, Germain M, Shore GC (2010) Antagonism of Beclin 1-dependent autophagy by BCL-2 at the endoplasmic reticulum requires NAF-1. EMBO J 29:606-618.
Chen YF, Kao CH, Chen YT, Wang CH, Wu CY, Tsai CY, Liu FC, Yang CW, Wei YH, Hsu MT, Tsai SF, Tsai TF (2009) Cisd2 deficiency drives premature aging and causes mitochondria-mediated defects in mice. Genes Dev 23:1183-1194.
Lin CC, Chiang TH, Chen WJ, Sun YY, Lee YH, Lin MS (2015) CISD2 serves a novel role as a suppressor of nitric oxide signalling and curcumin increases CISD2 expression in spinal cord injuries. Injury 46:2341-2350.
Lin MS, Lee YH, Chiu WT, Hung KS (2011a) Curcumin provides neuroprotection after spinal cord injury. J Surg Res 166:280-289.
Lin MS, Sun YY, Chiu WT, Hung CC, Chang CY, Shie FS, Tsai SH, Lin JW, Hung KS, Lee YH (2011b) Curcumin attenuates the expression and secretion of RANTES after spinal cord injury in vivo and lipopolysaccharide-induced astrocyte reactivation in vitro. J Neurotrauma 28:1259-1269.
Menon VP, Sudheer AR (2007) Antioxidant and anti-inflammatory properties of curcumin. Adv Exp Med Biol 595:105-125.
Wang Q, Sun AY, Simonyi A, Jensen MD, Shelat PB, Rottinghaus GE, Mac-Donald RS, Miller DK, Lubahn DE, Weisman GA, Sun GY (2005) Neuroprotective mechanisms of curcumin against cerebral ischemia-induced neuronal apoptosis and behavioral deficits. J Neurosci Res 82:138-148.
Weber WM, Hunsaker LA, Gonzales AM, Heynekamp JJ, Orlando RA, Deck LM, Vander Jagt DL (2006) TPA-induced up-regulation of activator protein-1 can be inhibited or enhanced by analogs of the natural product curcumin. Biochem Pharmacol 72:928-940.
Wu CY, Chen YF, Wang CH, Kao CH, Zhuang HW, Chen CC, Chen LK, Kirby R, Wei YH, Tsai SF, Tsai TF (2012) A persistent level of Cisd2 extends healthy lifespan and delays aging in mice. Hum Mol Genet 21:3956-3968.
10.4103/1673-5374.177718 http://www.nrronline.org/
How to cite this article: Lin CC, Lin MS (2016) New insight into curcumin-based therapy in spinal cord injuries: CISD2 regulation. Neural Regen Res 11(2):222-223.
杂志排行

中国神经再生研究(英文版)的其它文章
- Tissue-type plasminogen activator is a modulator of the synaptic vesicle cycle
- Impaired consciousness caused by injury of the lower ascending reticular activating system: evaluation by diffusion tensor tractography
- Considering calcium-binding proteins in invertebrates: multi-functional proteins that shape neuronal growth
- Cardiovascular dysfunction following spinal cord injury
- Practical application of the neuroregenerative properties of ketamine: real world treatment experience
- Exergames: neuroplastic hypothesis about cognitive improvement and biological effects on physical function of institutionalized older persons