Glucose metabolism and neurogenesis in the gerbil hippocampus after transient forebrain ischemia
2016-12-01DaeYoungYooKwonYoungLeeJoonHaParkHyoYoungJungJongWhiKimYeoSungYoonMooHoWonJungHoonChoiInKooHwangDepartmentofAnatomyandCellBiologyCollegeofVeterinaryMedicineandResearchInstituteforVeterinaryScienceSeoulNationalU
Dae Young Yoo, Kwon Young Lee, Joon Ha Park, Hyo Young Jung Jong Whi Kim Yeo Sung Yoon Moo-Ho Won, Jung Hoon Choi, In Koo Hwang Department of Anatomy and Cell Biology, College of Veterinary Medicine, and Research Institute for Veterinary Science, Seoul National University, Seoul, South Korea2 Department of Anatomy, College of Veterinary Medicine and Institute of Veterinary Science, Kangwon National University, Chuncheon, South Korea Department of Neurobiology, School of Medicine, Kangwon National University, Chuncheon, South Korea
Glucose metabolism and neurogenesis in the gerbil hippocampus after transient forebrain ischemia
Dae Young Yoo1,#, Kwon Young Lee2,#, Joon Ha Park3, Hyo Young Jung1, Jong Whi Kim1, Yeo Sung Yoon1, Moo-Ho Won3, Jung Hoon Choi2,*, In Koo Hwang1,*
1 Department of Anatomy and Cell Biology, College of Veterinary Medicine, and Research Institute for Veterinary Science, Seoul National University, Seoul, South Korea
2 Department of Anatomy, College of Veterinary Medicine and Institute of Veterinary Science, Kangwon National University, Chuncheon, South Korea
3 Department of Neurobiology, School of Medicine, Kangwon National University, Chuncheon, South Korea
How to cite this article: Yoo DY, Lee KY, Park JH, Jung HY, Kim JW, Yoon YS, Won MH, Choi JH, Hwang IK (2016) Glucose metabolism and neurogenesis in the gerbil hippocampus after transient forebrain ischemia. Neural Regen Res 11(8)∶1254-1259.
Funding: This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, No. NRF-2013R1A1A2059364, NRF-2015R1D1A3A01020635); by 2013 Research Grant from Kangwon National University, and also partially supported by the Research Institute for Veterinary Science, Seoul National University.
Jung Hoon Choi, D.V.M., Ph.D., or In Koo Hwang, D.V.M,
Ph.D., jhchoi@kangwon.ac.kr or vetmed2@snu.ac.kr.
#These authors contributed equally to this article.
orcid:
0000-0002-3725-4907
(Jung Hoon Choi)
0000-0002-0533-4638
(In Koo Hwang)
Accepted: 2016-06-21
Recent evidence exists that glucose transporter 3 (GLUT3) plays an important role in the energy metabolism in the brain. Most previous studies have been conducted using focal or hypoxic ischemia models and have focused on changes in GLUT3 expression based on protein and mRNA levels rather than tissue levels. In the present study, we observed change in GLUT3 immunoreactivity in the adult gerbil hippocampus at various time points after 5 minutes of transient forebrain ischemia. In the sham-operated group, GLUT3 immunoreactivity in the hippocampal CA1 region was weak, in the pyramidal cells of the CA1 region increased in a time-dependent fashion 24 hours after ischemia, and in the hippocampal CA1 region decreased significantly between 2 and 5 days after ischemia, with high level of GLUT3 immunoreactivity observed in the CA1 region 10 days after ischemia. In a double immunofluorescence study using GLUT3 and glial-fibrillary acidic protein (GFAP), we observed strong GLUT3 immunoreactivity in the astrocytes. GLUT3 immunoreactivity increased after ischemia and peaked 7 days in the dentate gyrus after ischemia/reperfusion. In a double immunofluorescence study using GLUT3 and doublecortin (DCX), we observed low level of GLUT3 immunoreactivity in the differentiated neuroblasts of the subgranular zone of the dentate gyrus after ischemia. GLUT3 immunoreactivity in the sham-operated group was mainly detected in the subgranular zone of the dentate gyrus. These results suggest that the increase in GLUT3 immunoreactivity may be a compensatory mechanism to modulate glucose level in the hippocampal CA1 region and to promote adult neurogenesis in the dentate gyrus.
nerve regeneration; transient forebrain ischemia; glucose transporter 3; pyramidal cells; astrocytes; neuroblasts; neural regeneration
Introduction
Glucose is the primary energy source for the brain and is essential for the maintenance of neuronal functions. Glucose can be delivered across the blood-brain barrier into glia and neurons by a family of structurally related membrane-spanning glycoproteins called glucose transporters (GLUTs) (Simpson et al., 2007). In particular, the more selective GLUT3 is primarily expressed by neuronal cells and meets the energy requirements necessary for synaptic neurotransmission (Maher et al., 1996).
Interruption of blood flow to the brain results in neuronal depletion of glucose and oxygen, which impairs ion transport, alters the resting potential of the membrane, and finally leads to the depolarization of neurons (Hwang et al., 2008; Wang et al., 2010). Accumulating evidence has indicated a compensatory increase in neuronal expression of GLUT3 in various forms of brain damage including ischemia (Urabe et al., 1996; Vannucci et al., 1998; Zovein et al., 2004; Huang et al., 2010), traumatic brain injury (Hamlin et al., 2001), glucose deprivation by insulin (Uehara et al., 1997) and hyperglycemia as a result of diabetes mellitus (Vannucci et al., 1994). Homozygous GLUT3-deficient mice show prenatal lethality, but heterozygous mice are born normally. However, they display autism spectrum-like behaviors such as abnormal cognitive flexibility with intact motor ability, perturbed social behavior with reduced vocalization and stereotypies, seizures, and decreased contextual memory (Zhao et al., 2010). However, most studies have been conducted using focal or hypoxic-ischemia models and have focused on changes in GLUT3 expression based on protein and mRNA levels rather than tissue levels.
To better understand the cellular localization of GLUT3 in the hippocampus at various time points after ischemia, we investigated the temporal and spatial changes of GLUT3expression in the hippocampal CA1 region and dentate gyrus of the Mongolian gerbil.
Materials and Methods
Experimental animals
Male Mongolian gerbils (Meriones unguiculatus, n = 49, 3-month-old, 50-60 g) were purchased from Japan SLC Inc. (Shizuoka, Japan). The animals were randomly divided into two groups; sham-operated (n = 5) and ischemia (n = 44) groups. They were housed under standard conditions with adequate temperature (22°C), humidity (60%), a 12-hour reversible light/dark cycle, and free access to food and water. The handling and care of the animals conformed to guidelines compliant with current international laws and policies (NIH Guide for the Care and Use of Laboratory Animals, NIH Publication No. 85-23, 1985, revised 1996). Animal procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Seoul National University. All experiments were conducted with an effort to minimize the number of animals used and the suffering caused by the procedures employed in the present study as previously described (Yoo et al., 2015).
Induction of transient forebrain ischemia
As previously described (Yoo et al., 2015), the animals were anesthetized with a mixture of 2.5% isoflurane (Baxtor, Deerfield, IL, USA) in 33% oxygen and 67% nitrous oxide. Common carotid arteries from both sides were isolated and occluded using non-traumatic aneurysm clips. The complete interruption of blood flow was confirmed by observing the obstruction of central artery in retinae using an ophthalmoscope (HEINE K180®, Heine Optotechnik, Herrsching, Germany). After 5 minutes of occlusion, the aneurysm clips were removed. Body temperature under free-regulating or normothermic (37 ± 0.5°C) conditions was monitored with a rectal temperature probe (TR-100; Fine Science Tools, Foster City, CA, USA) and maintained using a thermometric blanket before, during, and after the surgery until the animals completely recovered from anesthesia. Thereafter, animals were kept in a thermal incubator (Mirae Medical Industry, Seoul, South Korea) to maintain body temperature until the animals were euthanized. Sham-operated animals were subjected to the same surgical procedures except that the common carotid arteries were not occluded. Four animals were excluded due to incomplete occlusion of the common carotid arteries or uncontrolled body temperature.

Figure 1 Immunohistochemical staining for GLUT3 in the hippocampal CA1 region in sham-operated (A) and ischemia (B-I) groups.
Immunohistochemistry
For histology, sham-operated animals were anesthetized with 1 g/kg urethane (Sigma-Aldrich, St. Louis, MO, USA) 4 days after sham operation and ischemia-operated animals (n = 5 at each time point) were anesthetized at 6, 12, 24 hours, 2, 3, 5, 7, and 10 days after ischemia/reperfusion. The animals were perfused transcardially with 0.1 M phosphate-buffered saline (PBS, pH 7.4), followed by 4% paraformaldehyde in 0.1 M phosphate-buffer (pH 7.4), as previously described (Yoo et al., 2012b). The brains were removed and postfixed in the same fixative for 12 hours. The brain tissues were cryoprotected by infiltration with 30% sucrose overnight. Coronal brain sections (30 μm thick) were serially cut using a cryostat (Leica, Wetzlar, Germany). The sections were collected into 6-well plates containing PBS for further process.
To ensure that the immunohistochemical data were comparable between groups, the sections were carefully prounderthe same conditions as previously described (Jung et al., 2016). The tissue sections were selected between 1.4 and 2.0 mm posterior to the bregma in reference to a gerbil atlas (Loskota et al., 1974) for each animal. Five sections, 90 μm apart from each other, were sequentially treated with 0.3% hydrogen peroxide (H2O2) in PBS for 30 minutes and 10% normal goat serum in 0.05 M PBS for 30 minutes at room temperature. They were then incubated with rabbit anti-GLUT3 antibody (diluted 1:50, Santa Cruz Biotechnology, Santa Cruz, CA, USA) overnight at room temperature and subsequently treated with biotinylated goat anti-rabbit IgG and a streptavidin-peroxidase complex (1:200, Vector, Burlingame, CA, USA) for 2 hours at room temperature. Sections were then visualized by reaction with 3,3′-diaminobenzidine tetrachloride (Sigma) in 0.1 M Tris-HCl buffer (pH 7.2) and mounted on gelatin-coated slides. Sections were mounted in Canada Balsam (Kanto, Tokyo, Japan) after dehydration.
Data analysis
Analysis of the hippocampal CA1 region and dentate gyrus was performed using an image analysis system and ImageJ software v. 1.5 (National Institutes of Health, Bethesda, MD, USA), according to a previously described method (Jung et al., 2016). Digital images of the mid-point of the hippocampal CA1 region were captured with a BX51 light microscope (Olympus, Tokyo, Japan) equipped with a digital camera (DP72, Olympus) connected to a computer monitor. Images were calibrated into an array of 512 × 512 pixels corresponding to a tissue area of 1,200 μm × 900 μm (100× primary magnification). Each pixel resolution was 256 gray levels, and the intensity of GLUT3 immunoreactivity was evaluated by relative optical density (ROD), which was obtained after transformation of the mean gray level using the formula: ROD = log(256/mean grayscale level). ROD of background staining was determined in unlabeled portions of the sections using Photoshop CC software (Adobe Systems Inc., San Jose, CA, USA), and this value was subtracted to correct for nonspecific staining, using ImageJ v. 1.50 software (National Institutes of Health). Data are expressed as a percentage of the sham-operated group values (set to 100%).
Double immunofluorescence
To confirm the colocalization of GLUT3 and glial fibrillary acidic protein (GFAP) or GLUT3 and doublecortin (DCX) in the brain, the sections at 7 days after ischemia were processed by double immunofluorescence staining under the same conditions according to a previously described method (Jung et al., 2016). Double immunofluorescence staining for rabbit anti-GLUT3 (1:20) / goat anti-DCX (diluted 1:25; Santa Cruz Biotechnology) or mouse anti-GFAP (diluted 1:500, Millipore, Temecula, CA, USA) was performed. The sections were incubated in the mixture of antisera overnight at room temperature. After washing three times for 10 minutes each with PBS, they were then incubated in a mixture of both Cy3-conjugated donkey anti-rabbit IgG (1:600; Jackson ImmunoResearch, West Grove, PA, USA) and FITC-conjugated donkey anti-goat IgG (1:600; Jackson ImmunoResearch) or FITC-conjugated donkey anti-mouse IgG (1:600; Jackson ImmunoResearch) for 2 hours at room temperature. The immunoreactions were observed under the confocal microscope (LSM510 META NLO, Carl Zeiss, Göttingen, Germany).
Statistical analysis
As previously described (Yoo et al., 2015; Jung et al., 2016), the data are shown as the mean ± SEM. Differences among the means were statistically analyzed by one-way analysis of variance followed by a Bonferroni’s post-hoc test, using GraphPad Prism 5.01 software (GraphPad Software, Inc., La Jolla, CA, USA). Statistical significance was considered at P < 0.05.
Results
Changes in GLUT3 immunoreactivity in the hippocampal CA1 region
In the sham-operated group, very weak GLUT3 immunoreactivity was detected in the hippocampal CA1 region (Figure 1A). GLUT3 immunoreactivity in the CA1 region increased significantly compared to that in the sham-operated group 6 hours after ischemia (Figure 1B, J). A significant increase in GLUT3 immunoreactivity was detected in the pyramidal neurons of the CA1 region 12-24 hours after ischemia (Figure 1C, D, J). Two days after ischemia/reperfusion, GLUT3 immunoreactivity in the stratum pyramidale decreased significantly while an increase in GLUT3 immunoreactivity was found in the strata oriens and radiatum (Figure 1E). Three days after ischemia, GLUT3 immunoreactivity was seen in the degenerating cells of the stratum pyramidale, while GLUT3 immunoreactivity was not observed in other regions (Figure 1F). Five days after ischemia, only faint GLUT3 immunoreactivity was detected in the CA1 region (Figure 1G). Seven to ten days after ischemia, abundant GLUT3 immunoreactivity was detected in the strata oriens and radiatum (Figure 1H, I). In the ischemia group, GLUT3 immunoreactivity was significantly increased in the hippocampal CA1 region 6, 12, 24 hours, 2, 3, 7, 10 days after ischemia than 5 days after ischemia (Figure 1J). GLUT3 immunoreactive structures also showed GFAP immunoreactivity with the activated form (hypertrophied cytoplasm) in the strata oriens and radiatum of the hippocampal CA1 region (Figure 3A-C).
Changes in GLUT3 immunoreactivity in the dentate gyrus In the sham-operated group, a moderate level of GLUT3 immunoreactivity was observed in the subgranular zone of the dentate gyrus (Figure 2A). Unlike what we observed in the hippocampal CA1 region, GLUT3 immunoreactivity did not change spatially in the dentate gyrus although the GLUT3 immunoreactivity did. GLUT3 immunoreactivity in the dentate gyrus increased significantly compared to that observed in the sham-operated group 6 hours after ischemia (Figure 2B, J). GLUT3 immunoreactivity 12 hours after ischemia was similar to that observed 6 hours after ischemia, althougha significant increase was observed 24 hours after ischemia compared to 12 hours after ischemia (Figure 2C, D, J). Two to five days after ischemia, GLUT3 immunoreactivity in the dentate gyrus decreased compared to the levels observed 24 hours after ischemia, although it was significantly greater in the ischemia group at these time points compared to the levels observed in the sham-operated group (Figure 2J). GLUT3 immunoreactivity remained significantly increased 7 days after ischemia, and clear GLUT3 immunoreactive band-like structures were found in the subgranular zone of the dentate gyrus. These band-like structures were not present in either the sham-operated group or at 5 days after ischemia (Figure 2H, J). The results of the double immunofluorescence study indicated partial colocalization of GLUT3 immunoreactive structures such as cytosol and processes in DCX-immunoreactive neuroblasts (Figure 3D-F). Ten days after ischemia, GLUT3 immunoreactivity in the subgranular zone of the dentate gyrus showed a decrease compared to the level observed 7 days after ischemia (Figure 2I and J). However, GLUT3 immunoreactivity in the ischemia group 7 days after ischemia was significantly greater than that in the sham-operated group (Figure 2J).
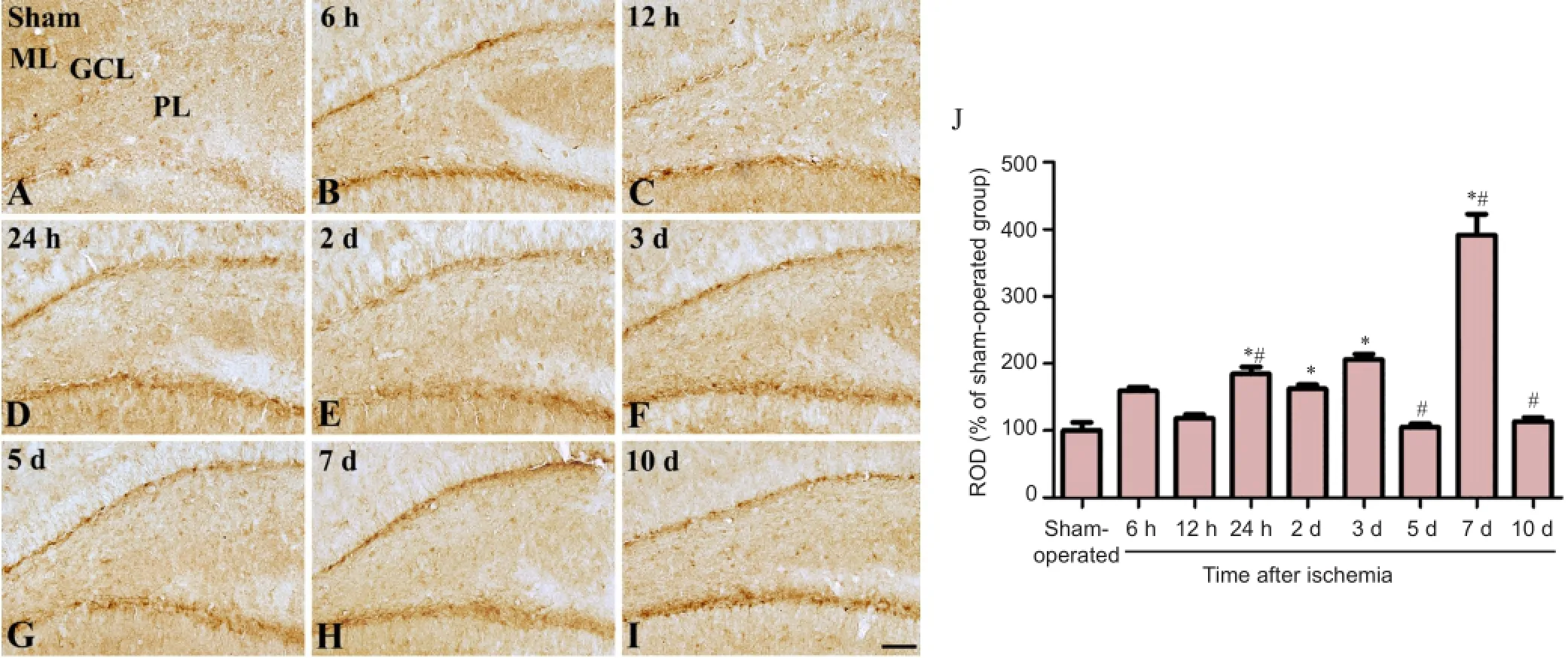
Figure 2 Immunohistochemical staining for GLUT3 in the dentate gyrus in sham-operated (A) and ischemia (B-I) groups.

Figure 3 Double immunofluorescence staining for GLUT3 (A, green), GFAP (B, red), and merged images (C, yellow) in the hippocampal CA1 region as well as GLUT3 (D, green), DCX (E, red), and merged images (F, yellow) in the dentate gyrus 7 days after ischemia.
Discussion
Glucose is the primary energy source for brain metabolism, and the passage of glucose across the cell membrane is facilitated by a family of integral membrane transporter proteins known as GLUTs (Simpson et al., 2007). GLUT3 is the predominant isoform that fuels ATP generation and thereby energy metabolism in neurons (Simpson et al., 2007). Cerebral ischemia has been associated with changes in both glucose utilization and cerebral metabolic rate (Pulsinelli et al., 1982; Suzuki et al., 1983). In the present study, we observed GLUT3 in the pyramidal neurons and subgranular zone of the dentate gyrus in the Mongolian gerbil. This observation is supported by previous rat/mouse studies reporting GLUT3 immunoreactivity that is primarily localized in neuronal components (Nagamatsu et al., 1994; Vannucci et al., 1997; Choeiri et al., 2002; Simpson et al., 2008).
A significant increase in GLUT3 immunoreactivity was observed in the pyramidal neurons of the hippocampal CA1 region 24 hours after ischemia. This result coincides with those of previous studies reporting increased GLUT3 levels in response to ischemia in the brain (Gerhart et al., 1994; Urabe et al., 1996; Zovein et al., 2004; Fung et al., 2010; Huang et al., 2010). However, in the present study, we observed biphasic increases in GLUT3 immunoreactivity in pyramidal neurons, followed by increases in immunoreactivity in the astrocytes of the hippocampal CA1 region. The switch from neurons to astrocytes may be associated with neuronal death in the hippocampus as the steady glucose supply provided by astrocytes (Danbolt, 2001) becomes the main source of energy after neuronal damage in the hippocampal CA1 region (Vilchez et al., 2007). In a model of focal cerebral ischemia using gerbils, GLUT3 expression increased 10-fold 24 hours following ischemia (Huang et al., 2010). In addition, in transient focal ischemia after 3 hours of middle cerebral artery occlusion, GLUT3 mRNA and protein levels increased in the rat brain 48 hours after reperfusion (Urabe et al., 1996). After 45 minutes of hypoxic ischemia, a significant increase in GLUT3 expression was observed in both the ipsilateral and contralateral sides of the brain 4 hours after re-oxygenation, returning to baseline levels 24 hours after re-oxygenation (Fung et al., 2010).
In the dentate gyrus, we observed increased levels of GLUT3 immunoreactivity after transient forebrain ischemia, and these GLUT3 immunoreactive cells were identified as differentiated neuroblasts 7 days after ischemia. The increase in GLUT3 expression may be a compensatory mechanism intended to induce neurogenesis in the dentate gyrus. This observation is supported by our previous study in which GLUT3 immunoreactive cells were found to overlap with DCX-immunoreactive neuroblasts in the postnatal brain (Jung et al., 2016). In addition, GLUT3 immunoreactivity in the dentate gyrus peaked 7 days after ischemia. This result may be closely related with the chronological changes observed in DCX immunoreactive neuroblasts (Choi et al., 2012). Several lines of evidence show that cAMP response element-binding protein (CREB) is an important transcription factor of the GLUT3 gene (Rajakumar et al., 2004), potentially binding to the promoter region of the gene and regulating GLUT3 expression (Reddy et al., 2010; Jin et al., 2013). Phosphorylation of CREB protein on serine 133 is implicated in activity-dependent neuronal plasticity, neurotrophin-mediated neuronal survival, and adult neurogenesis (Ghosh and Greenberg, 1995; Riccio et al., 1999). The genetic down-regulation of GLUT3 expression accelerates neuronal damage (Fung et al., 2010) and reduces preconditioning effects on ischemic tolerance (Iwabuchi and Kawahara, 2011).
In conclusion, our findings suggest that the transient increases in GLUT3 expression following forebrain ischemia may be a compensatory mechanism aimed at reducing the neuronal damage in the hippocampal CA1 region and promoting endogenous neurogenesis in the dentate gyrus.
Acknowledgments: The authors would like to thank to Seung-Hae Kwon from Chuncheon Center, Korea Basic Science Institute, South Korea for technical assistance with the confocal image analyses (LSM 510 META NLO).
Author contributions: All authors conceived the experiment and paper. DYY, KYL, JHC, and IKH designed the experiment. DYY and KYL wrote the paper and JHC and IKH edited the paper. DYY, KYL, HYJ, and JWK conducted the immunohistochemistry and analyzed the data. JHP conducted double immunohistochemistry. DYY and KYL made the animal model for ischemia. YSY and MHW advised the design of experiment and edited the paper. All authors approved the final version of this paper.
Conflicts of interest: None declared.
Plagiarism check: This paper was screened twice using Cross-Check to verify originality before publication.
Peer review: This paper was double-blinded and stringently reviewed by international expert reviewers.
References
Choeiri C, Staines W, Messier C (2002) Immunohistochemical localization and quantification of glucose transporters in the mouse brain. Neuroscience 111:19-34.
Choi JH, Yoo KY, Lee CH, Park JH, Yan BC, Kwon SH, Seo JY, Cho JH, Hwang IK, Won MH (2012) Comparison of neurogenesis in the dentate gyrus between the adult and aged gerbil following transient global cerebral ischemia. Neurochem Res 37:802-810.
Danbolt NC (2001) Glutamate uptake. Prog Neurobiol 65:1-105.
Fung C, Evans E, Shin D, Shin BC, Zhao Y, Sankar R, Chaudhuri G, Devaskar SU (2010) Hypoxic-ischemic brain injury exacerbates neuronal apoptosis and precipitates spontaneous seizures in glucose transporter isoform 3 heterozygous null mice. J Neurosci Res 88:3386-3398.
Gerhart DZ, Leino RL, Taylor WE, Borson ND, Drewes LR (1994) GLUT1 and GLUT3 gene expression in gerbil brain following brief ischemia: an in situ hybridization study. Brain Res Mol Brain Res 25:313-322.
Ghosh A, Greenberg ME (1995) Calcium signaling in neurons: molecular mechanisms and cellular consequences. Science 268:239-247.
Hamlin GP, Cernak I, Wixey JA, Vink R (2001) Increased expression of neuronal glucose transporter 3 but not glial glucose transporter 1 following severe diffuse traumatic brain injury in rats. J Neurotrauma 18:1011-1018.
Huang CY, Liou YF, Chung SY, Pai PY, Kan CB, Kuo CH, Tsai CH, Tsai FJ, Chen JL, Lin JY (2010) Increased expression of glucose transporter 3 in gerbil brains following magnesium sulfate treatment and focal cerebral ischemic injury. Cell Biochem Funct 28:313-320.
Hwang IK, Yoo KY, An SJ, Li H, Lee CH, Choi JH, Lee JY, Lee BH, Kim YM, Kwon YG, Won MH (2008) Late expression of Na+/H+exchanger 1 (NHE1) and neuroprotective effects of NHE inhibitor in the gerbil hippocampal CA1 region induced by transient ischemia. Exp Neurol 212:314-323.
Iwabuchi S, Kawahara K (2011) Inducible astrocytic glucose transporter-3 contributes to the enhanced storage of intracellular glycogen during reperfusion after ischemia. Neurochem Int 59:319-325.
Jin N, Qian W, Yin X, Zhang L, Iqbal K, Grundke-Iqbal I, Gong CX, Liu F (2013) CREB regulates the expression of neuronal glucose transporter 3: a possible mechanism related to impaired brain glucose uptake in Alzheimer’s disease. Nucleic Acids Res 41:3240-3256.
Jung HY, Yim HS, Yoo DY, Kim JW, Chung JY, Seong JK, Yoon YS, Kim DW, Hwang IK (2016) Postnatal changes in glucose transporter 3 expression in the dentate gyrus of the C57BL/6 mouse model. Lab Anim Res 32:1-7.
Loskota WA, Lomax P, Verity MA (1974) A stereotaxic atlas of the Mongolian gerbil brain (Meriones unguiculatus). Ann Arbor: Ann Arbor Science Publishers Inc.
Maher F, Davies-Hill TM, Simpson IA (1996) Substrate specificity and kinetic parameters of GLUT3 in rat cerebellar granule neurons. Biochem J 315:827-831.
Nagamatsu S, Sawa H, Inoue N, Nakamichi Y, Takeshima H, Hoshino T (1994) Gene expression of GLUT3 glucose transporter regulated by glucose in vivo in mouse brain and in vitro in neuronal cell cultures from rat embryos. Biochem J 300:125-131.
Pulsinelli WA, Levy DE, Duffy TE (1982) Regional cerebral blood flow and glucose metabolism following transient forebrain ischemia. Ann Neurol 11:499-502.
Rajakumar A, Thamotharan S, Raychaudhuri N, Menon RK, Devaskar SU (2004) Trans-activators regulating neuronal glucose transporter isoform-3 gene expression in mammalian neurons. J Biol Chem 279:26768-26779.
Reddy AB, Srivastava SK, Ramana KV (2010) Aldose reductase inhibition prevents lipopolysaccharide-induced glucose uptake and glucose transporter 3 expression in RAW264.7 macrophages. Int J Biochem Cell Biol 42:1039-1045.
Riccio A, Ahn S, Davenport CM, Blendy JA, Ginty DD (1999) Mediation by a CREB family transcription factor of NGF-dependent survival of sympathetic neurons. Science 286:2358-2361.
Simpson IA, Carruthers A, Vannucci SJ (2007) Supply and demand in cerebral energy metabolism: the role of nutrient transporters. J Cereb Blood Flow Metab 27:1766-1791.
Simpson IA, Dwyer D, Malide D, Moley KH, Travis A, Vannucci SJ (2008) The facilitative glucose transporter GLUT3: 20 years of distinction. Am J Physiol Endocrinol Metab 295:E242-253.
Suzuki R, Yamaguchi T, Kirino T, Orzi F, Klatzo I (1983) The effects of 5-minute ischemia in Mongolian gerbils: I. Blood-brain barrier, cerebral blood flow, and local cerebral glucose utilization changes. Acta Neuropathol 60:207-216.
Uehara Y, Nipper V, McCall AL (1997) Chronic insulin hypoglycemia induces GLUT-3 protein in rat brain neurons. Am J Physiol 272:E716-E719.
Urabe T, Hattori N, Nagamatsu S, Sawa H, Mizuno Y (1996) Expression of glucose transporters in rat brain following transient focal ischemic injury. J Neurochem 67:265-271.
Vannucci SJ, Maher F, Simpson IA (1997) Glucose transporter proteins in brain: delivery of glucose to neurons and glia. Glia 21:2-21.
Vannucci SJ, Maher F, Koehler E, Simpson IA (1994) Altered expression of GLUT-1 and GLUT-3 glucose transporters in neurohypophysis of water-deprived or diabetic rats. Am J Physiol 267:E605-E611.
Vannucci SJ, Reinhart R, Maher F, Bondy CA, Lee WH, Vannucci RC, Simpson IA (1998) Alterations in GLUT1 and GLUT3 glucose transporter gene expression following unilateral hypoxia-ischemia in the immature rat brain. Brain Res Dev Brain Res 107:255-264.
Vilchez D, Ros S, Cifuentes D, Pujadas L, Vallès J, García-Fojeda B, Criado-García O, Fernández-Sánchez E, Medraño-Fernández I, Domínguez J, García-Rocha M, Soriano E, Rodríguez de Córdoba S, Guinovart JJ (2007) Mechanism suppressing glycogen synthesis in neurons and its demise in progressive myoclonus epilepsy. Nat Neurosci 10:1407-1413.
Wang Y, Denisova JV, Kang KS, Fontes JD, Zhu BT, Belousov AB (2010) Neuronal gap junctions are required for NMDA receptor-mediated excitotoxicity: implications in ischemic stroke. J Neurophysiol 104:3551-3556.
Yoo DY, Jung HY, Nam SM, Kim JW, Choi JH, Kwak YG, Yoo M, Lee S, Yoon YS, Hwang IK (2015) Valeriana officinalis extracts ameliorate neuronal damage by suppressing lipid peroxidation in the gerbil hippocampus following transient cerebral ischemia. J Med Food 18:642-647.
Zhao Y, Fung C, Shin D, Shin BC, Thamotharan S, Sankar R, Ehninger D, Silva A, Devaskar SU (2010) Neuronal glucose transporter isoform 3 deficient mice demonstrate features of autism spectrum disorders. Mol Psychiatry 15:286-299.
Zovein A, Flowers-Ziegler J, Thamotharan S, Shin D, Sankar R, Nguyen K, Gambhir S, Devaskar SU (2004) Postnatal hypoxic-ischemic brain injury alters mechanisms mediating neuronal glucose transport. Am J Physiol Regul Integr Comp Physiol 286:R273-282.
Copyedited by Jackson C, Li HF, Liu ZFSong LP, Liu WJ, Zhao M
10.4103/1673-5374.189189
*Correspondence to:
杂志排行

中国神经再生研究(英文版)的其它文章
- Secondary parkinsonism induced by hydrocephalus after subarachnoid and intraventricular hemorrhage
- Prospects for bone marrow cell therapy in amyotrophic lateral sclerosis: how far are we from a clinical treatment?
- Uncoupling protein 2 in the glial response to stress: implications for neuroprotection
- Selective neuronal PTEN deletion: can we take the brakes off of growth without losing control?
- TRPV1 may increase the effectiveness of estrogen therapy on neuroprotection and neuroregeneration
- Tamoxifen: an FDA approved drug with neuroprotective effects for spinal cord injury recovery