Volume transmission and receptor-receptor interactions in heteroreceptor complexes: understanding the role of new concepts for brain communication
2016-12-01KjellFuxeDasielBorrotoEscuela1DepartmentofNeuroscienceKarolinskaInstitutetStockholmSweden2ObservatorioCubanodeNeurocienciasGrupoBohEstudioYaguajayCuba
Kjell Fuxe, Dasiel O. Borroto-Escuela1 Department of Neuroscience, Karolinska Institutet, Stockholm, Sweden2 Observatorio Cubano de Neurociencias, Grupo Bohío-Estudio, Yaguajay, Cuba
Volume transmission and receptor-receptor interactions in heteroreceptor complexes: understanding the role of new concepts for brain communication
Kjell Fuxe1,*, Dasiel O. Borroto-Escuela1,2,*
1 Department of Neuroscience, Karolinska Institutet, Stockholm, Sweden
2 Observatorio Cubano de Neurociencias, Grupo Bohío-Estudio, Yaguajay, Cuba
How to cite this article: Fuxe K, Borroto-Escuela DO (2016) Volume transmission and receptor-receptor interactions in heteroreceptor complexes∶understanding the role of new concepts for brain communication. Neural Regen Res 11(8)∶1220-1223.
Funding: The authors are supported by grants from the Swedish Medical Research Council (04X-715) to KF, by AFA Försäkring (130328) to KF and DOBE, and by Hjärnfonden to DOBE. DOBE belong to Academia de Biólogos Cubanos.
Kjell Fuxe, M.D. or
Dasiel O. Borroto-Escuela,
Ph.D., kjell.fuxe@ki.se or
dasiel.borroto.escuela@ki.se.
orcid: 0000-0002-5736-373X
(Dasiel O. Borroto-Escuela)
Accepted: 2016-07-18
The discovery of the central monoamine neurons not only demonstrated novel types of brain stem neurons forming global terminal networks all over the brain and the spinal cord, but also to a novel type of communication called volume transmission. It is a major mode of communication in the central nervous system that takes places in the extracellular fluid and the cerebral spinal fluid through diffusion and flow of molecules, like neurotransmitters and extracellular vesicles. The integration of synaptic and volume transmission takes place through allosteric receptor-receptor interactions in heteroreceptor complexes. These heterocomplexes represent major integrator centres in the plasma membrane and their protomers act as moonlighting proteins undergoing dynamic changes and their structure and function. In fact, we propose that the molecular bases of learning and memory can be based on the reorganization of multiples homo and heteroreceptor complexes into novel assembles in the post-junctional membranes of synapses.
volume transmission; receptor-receptor interaction; heteroreceptor complexes; oligomerization; brain communication; learning; memory; central monoamine neurons
Central Monoamine Neurons
The central dopamine (DA), noradrenaline (NA) and 5-hydroxytryptamine (5-HT) neurons in the central nervous system (CNS) were discovered using the Falck-Hillarp technique and immunohistochemistry (Dahlstroem and Fuxe, 1964; Fuxe, 1965). Amazingly, these brainstem monoamine neurons were found to have extensive monosynaptic projections and collaterals forming global terminal networks all over the brain and the spinal cord. The parcellation of the monoamine cell groups in the brainstem was found to be phylogenetically highly conserved (Fuxe et al., 2009). They represent the foundation for our modern understanding of cellular communication in the brain, how drugs act on the brain and how this communication is altered in CNS disorders like Parkinson’s disease, depression, schizophrenia, addiction, etc. It gave the beginning of a cellular basis for neuropsychopharmacology.
Volume Transmission
The discovery of the central monomine neurons also gave volume transmission (VT) to the wired brain (Fuxe et al., 2009). It was the discovery of the DA, NA and 5-HT neurons in the mammalian CNS in the 1960s and especially their pharmacological characterization that gave the essence of the experimental support for the introduction of the volume transmission theory by Fuxe and Agnati (1986). Since the first international symposium on “Volume transmission in the brain” organized in 1990 in Stockholm (Fuxe and Agnati, 1991), VT began to be considered a major mode of communication in the brain, which also is used by neuropsychoactive drugs to reach their targets (Nicholson and Sykova, 1998). VT represents a widespread mode of intercellular communication that occurs in the extracellular fluid of the brain and in the cerebrospinal fluid. VT signals are molecules like neurotransmitters, trophic factors, ions and gases (e.g., NO) and move from the source cells to the target cells as a consequence of energy gradients leading to diffusion and flow (Descarries and Mechawar, 2000; Trueta and De-Miguel, 2012).
DA, NA and 5-HT neurons were shown to operate mainly via short distance (extrasynaptic) VT in the micrometers range in the local circuits of the CNS where the monoamine receptors are mainly extrasynaptically located. Long-distance VT over millimeters mainly involve peptide/proteintransmission like beta-endorphin, oxytocin, prolactin-like and interleukin-1beta transmission (Fuxe et al., 2009). The release of neuropeptides may allow the monoamine neurons to send VT signals to cellular networks further away from the monoamine terminal networks.
Recently, we proposed that extracellular vesicles (exosomes and shedding vesicles) mediate a special form of VT in the CNS (Borroto-Escuela et al., 2015) based on the fundamental work of Simons and Raposo (2009) in peripheral tissue. In this novel type of VT extracellular vesicles are safe vesicular carriers for targeted intercellular communication of proteins, including receptors and homeoproteins, mtDNA and different forms of RNA in the CNS migrating in the extracellular fluid along energy gradients to reach adjacent target cells.
Knowing the integration of synaptic transmission and VT will be fundamental for the understanding of brain function and the concept is introduced that a major site of this integration is represented by heteroreceptor complexes in which inter alia ion channel receptors / G protein coupled receptors (GPCRs) and GPCR/GPCR integrate their activity by direct receptor-receptor interactions through allosteric mechanisms (Fuxe et al., 2014b; Figure 1). The same neural-glial network can give a different balance of its outputs. This happens when the diffusing VT-signals (glially and/or neuronally formed) by up-regulating or down-regulating synaptic contacts mainly through receptor-receptor interactions which can change the integrative action of the network and differences in their balance of the outputs are achieved (Fuxe et al., 2013; Borroto-Escuela et al., 2015). This will have a major impact on the flow of information through the brain circuits regulated by this cellular network and thus on brain function and behaviors. Within the neural-glial network we postulated the existence of the CNS trophic unit. It consists of: neurons; glial cells; pericytes; blood vessels with endothelial cells; extracellular matrix (Borroto-Escuela et al., 2015). This term was used to indicate the smallest set of cells within the neural-glial networks which act in a complementary way to support the trophism of one another and includes the neurovascular unit. The neural network is postulated to survive and function through integration of volume transmission and wiring transmission in the trophic unit (Borroto-Escuela et al., 2015). This trophism gives the energy necessary for the neuronal network and its brain circuits to operate.
Receptor-Receptor Interactions in Heteroreceptor Complexes
In 1980-1983, we obtained, for the first time, indications for the existence of direct interactions between different types of GPCRs potentially forming heteroreceptor complexes with allosteric receptor-receptor interactions based on the ability of neuropeptides to modulate the binding characteristics of subtypes of monoamine receptors in membrane preparations (Fuxe et al., 1983). It is now concluded that GPCR heteroreceptor complexes with allosteric receptor-receptor interactions operating through the receptor interface have become major integrative centers at the molecular level and their receptor protomers act as moonlighting proteins (Fuxe et al., 2014a, b). The GPCR heteroreceptor complexes in the CNS have become exciting new targets for neurotherapeutics in Parkinson’s disease, schizophrenia, drug addiction, anxiety and depression opening a new field in neuropsychopharmacology (Fuxe et al., 2014b).
Based on a mathematical approach, Tarakanov and Fuxe in 2010 deduced a set of triplet amino acid homologies that may importantly participate in the receptor interface of heteromers. They show how such protriplets of amino acid residues and their ‘teams’ may be utilized to construct a kind of code that determines (and/or predicts) which receptors should or should not form heterodimers (Borroto-Escuela et al., 2012). Based on the obtained results, they proposed a ‘guide-and-clasp’ manner for receptor-receptor interactions where ‘adhesive guides’ might be the triplet homologies.
Over many years evidence for the existence of antagonistic receptor-receptor interactions in A2A-D2 heteroreceptor complexes was obtained in the ventral and dorsal striatum and was validated with proximity ligation assay. In 2011, indications were obtained for the existence of a possible A2A-D2-β-Arrestin2 complex. The results suggested that the antagonistic A2AR-D2R allosteric receptor-receptor interaction in A2AR-D2R heteromers favors β-arrestin2 recruitment versus Gi/o coupling to the D2R protomer with subsequent cointernalization associated with a reduced time onset of Akt phosphorylation followed by a rapid dephosphorylation. Thus, a moonlighting phenomenon with a change of D2 receptor function takes place (Fuxe et al., 2014b; George et al., 2014).
Increased support was obtained for the view that A2A agonists at the A2A protomer in the A2A-D2 heteroreceptor complex are novel atypical antipsychotic drugs for treatment of schizophrenia as well as drugs for treatment of cocaine use disorder (Fuxe et al., 2014b; Figure 1). A2A receptor antagonists can instead act as blockers of the A2A protomer in the A2A-D2 heteroreceptor complex are antiparkinson drugs for the treatment of Parkinson’s disease. It represents a continuation of our previous work starting already in 1974 on effects of methylxanthines in hemiparkinson rats. This is a rational continuation of our search for novel antiantiparkisson drugs since the late 1960s and 1970s, which resulted in the development of a number of DA receptor agonists like apomorphine, ET495 and pravidel (bromocriptine) for treatment of Parkinson’s disease.
Our demonstration of facilitatory allosteric D2R-OTR interactions in the accumbens heteroreceptor complexes may represent at least a significant part of the molecular mechanism for oxytocin-induced changes in social and emotional behavior. D2R-OTR heteroreceptor complexes may become a new target for drug development and treatment of dysfunctions in the emotional networks of the brain taking place e.g., in schizophrenia as seen from the negative symptoms. Ourwork demonstrated that D2R is in fact a hub receptor capable of forming heteromers with many other types of GPCRs in cellular models and likely also in the neuronal-glial networks of the brain (Borroto-Escuela et al., 2014).
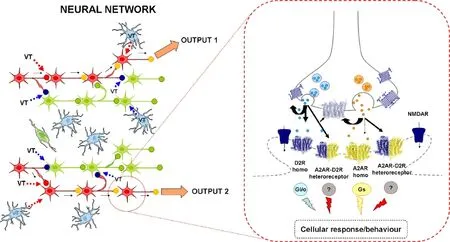
Figure 1 Balancing and integration of volume transmission (VT) and synaptic transmission through receptor-receptor interactions.
Our findings indicate that neurotrophic and antidepressant effects of 5-HT in brain may be mediated by activation of the 5-HT1A receptor protomer in the hippocampal FGFR1-5-HT1A receptor complex enhancing the FGFR1 signaling (Borroto-Escuela et al., 2012, 2016). Thus, a molecular neurotrophic mechanism exists in 5-HT nerve cell communication the activation of which may cause a relief from depression. A new strategy for treatment of depression is given. It represents an exciting continuation of the work of our group in this field which started with the introduction by Carlsson, Fuxe and Ungerstedt in 1968 of a novel target for antidepressants, the serotonin reuptake mechanism in the serotonin neurons (Carlsson et al., 1968). It led to the development of the selective serotonin re-uptake inhibitors (SSRIs).
Novel Concept on the Molecular Basis of Learning and Memory (Molecular Engram)
Molecular basis of learning and memory (molecular engram) is proposed to be based on the reorganization of multiple homo- heteroreceptor complexes into novel molecular assemblies in the postjunctional membrane of synapses with changes in the prejunctional receptor complexes to facilitate the pattern of transmitter release to be learned (Fuxe et al., 2014a; Borroto-Escuela et al., 2015).
Learning. A new temporal pattern of release of multiple transmitters in the synapse is learnt through a transient reorganization of sets of homo- and heteroreceptor complexes (receptors, ion channels, adapter proteins) in the postsynaptic and adjacent perisynaptic membranes. This results in novelallosteric receptor-receptor interactions altering receptor protomer functions. In this way a short-term memory is created from the novel pattern of transmitter release to be learned leading to a novel transient bar-code representing a molecular engram of short term memory (Fuxe et al., 2014a; Borroto-Escuela et al., 2015). The novel pattern of transmitter release to be learned can be stabilized by reorganization of pre-synaptic and associated perisynaptic homo-heteroreceptor complexes. This can involve agonist dependent processes, release of soluble molecules like neuropeptides and trophic factors and exosomes from the post-perisynaptic membranes as a result of the new bar-code.
Long-term memory may be created by the transformation of parts of the heteroreceptor complexes into unique transcription factors which can lead to the formation of specific adapter proteins which can consolidate the heteroreceptor complexes into long-lived complexes with conserved allosteric receptor-receptor interactions (Fuxe et al., 2014a; Borroto-Escuela et al., 2015). Thus, new types of “barcodes”can be formed based on reorganized homo-and heteroreceptor complexes and on novel receptor complexes formed in the post and perisynaptic membranes of synapses through allosteric mechanisms:
1. Production of unique transcription factors from the new barcode.
2. Unique adapter proteins formed through trough these transcription factors. Thus, protein synthesis is needed.
3. Consolidation of the receptor complexes themselves and also to their link to the cytoskeleton.
4. Volume transmission signals from adjacent terminals of emotional pathways reaching the postjunctional membrane leads to life-long memories through exceptional formation of special adapter proteins.
Author contributions: KF wrote most of the text of the article and assisted with editing figures and figure legends and organizing references; DOBE proposed the original version of the figures designed the figures and provided critical commentary, text and references. All authors participated in the organization of the article and topics to be covered and approved the final version of the article.
Conflicts of interest: None declared.
References
Agnati LF, Fuxe K, Zoli M, Ozini I, Toffano G, Ferraguti F (1986) A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta Physiol Scand 128:201-207.
Borroto-Escuela DO, Agnati LF, Bechter K, Jansson A, Tarakanov AO, Fuxe K (2015) The role of transmitter diffusion and flow versus extracellular vesicles in volume transmission in the brain neural-glial networks. Philos Trans R Soc Lond B Biol Sci 370(1672). pii: 20140183.
Borroto-Escuela DO, Brito I, Romero-Fernandez W, Di Palma M, Oflijan J, Skieterska K, Duchou J, Van Craenenbroeck K, Suarez-Boomgaard D, Rivera A, Guidolin D, Agnati LF, Fuxe K (2014) The G protein-coupled receptor heterodimer network (GPCR-Het-Net) and its hub components. Int J Mol Sci 15:8570-8590.
Borroto-Escuela DO, Romero-Fernandez W, Mudo G, Perez-Alea M, Ciruela F, Tarakanov AO, Narvaez M, Di Liberto V, Agnati LF, Belluardo N, Fuxe K (2012) Fibroblast growth factor receptor 1-5-hydroxytryptamine 1A heteroreceptor complexes and their enhancement of hippocampal plasticity. Biol Psychiatry 71:84-91.
Borroto-Escuela DO, Tarakanov AO, Fuxe K (2016) FGFR1-5-HT1A heteroreceptor complexes: implications for understanding and treating major depression. Trends Neurosci 39:5-15.
Carlsson A, Fuxe K, Ungerstedt U (1968) The effect of imipramine on central 5-hydroxytryptamine neurons. J Pharm Pharmacol 20:150-151.
Dahlstroem A, Fuxe K (1964) Evidence for the existence of monoamine-containing neurons in the central nervous system. I. demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol Scand Suppl 232:231-255.
Descarries L, Mechawar N (2000) Ultrastructural evidence for diffuse transmission by monoamine and acetylcholine neurons of the central nervous system. Prog Brain Res 125:27-47.
Fuxe K (1965) Evidence for the existence of monoamine neurons in the central nervous system. Iv. distribution of monoamine nerve terminals in the central nervous system. Acta Physiol Scand Suppl 247:237+.
Fuxe K, Agnati LF (1991) Volume transmission in the brain, novel mechanisms for neural transmission. New York: Raven Press.
Fuxe K, Agnati LF, Benfenati F, Celani M, Zini I, Zoli M, Mutt V (1983) Evidence for the existence of receptor--receptor interactions in the central nervous system. Studies on the regulation of monoamine receptors by neuropeptides. J Neural Transm Suppl 18:165-179.
Fuxe K, Borroto-Escuela DO, Ciruela F, Guidolin D, Agnati LF (2014a) Receptor-receptor interactions in heteroreceptor complexes: a new principle in biology. Focus on their role in learning and memory. Neurosci Discov doi: 10.7243/2052-6946-2-6.
Fuxe K, Borroto-Escuela DO, Romero-Fernandez W, Palkovits M, Tarakanov AO, Ciruela F, Agnati LF (2014b) Moonlighting proteins and protein-protein interactions as neurotherapeutic targets in the G protein-coupled receptor field. Neuropsychopharmacology 39:131-155.
Fuxe K, Borroto-Escuela DO, Tarakanov A, Romero Fernandez R, Manger P, Rivera A, van Craenenbroeck K, Skieterska K, Diaz-Cabiale Z, Filip M, Ferraro L, Tanganelli S, Guidolin D, Cullheim S, Perez de la Mora M, Agnati LF (2013) Understanding the balance and integration of volume and synaptic transmission. Relevance for psychiatry. Neurol Psychiatry Brain Res 19:141-158.
Fuxe K, Dahlstrom AB, Jonsson G, Marcellino D, Guescini M, Dam M, Manger P, Agnati L (2009) The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog Neurobiol 90:82-100.
George SR, Kern A, Smith RG, Franco R (2014) Dopamine receptor heteromeric complexes and their emerging functions. Prog Brain Res 211:183-200.
Nicholson C, Sykova E (1998) Extracellular space structure revealed by diffusion analysis. Trends Neurosci 21:207-215.
Simons M, Raposo G (2009) Exosomes--vesicular carriers for intercellular communication. Curr Opin Cell Biol 21:575-581.
Trueta C, De-Miguel FF (2012) Extrasynaptic exocytosis and its mechanisms: a source of molecules mediating volume transmission in the nervous system. Front Physiol 3:319.
10.4103/1673-5374.189168
*Correspondence to:
杂志排行

中国神经再生研究(英文版)的其它文章
- Secondary parkinsonism induced by hydrocephalus after subarachnoid and intraventricular hemorrhage
- Prospects for bone marrow cell therapy in amyotrophic lateral sclerosis: how far are we from a clinical treatment?
- Uncoupling protein 2 in the glial response to stress: implications for neuroprotection
- Selective neuronal PTEN deletion: can we take the brakes off of growth without losing control?
- TRPV1 may increase the effectiveness of estrogen therapy on neuroprotection and neuroregeneration
- Tamoxifen: an FDA approved drug with neuroprotective effects for spinal cord injury recovery