黄、东海陆架2011年秋季活体浮游有孔虫生态分布
2016-11-30吕红红向荣
吕红红,向荣
(1.中国科学院南海海洋研究所中国科学院边缘海地质重点实验室,广东广州510301;2.中国科学院大学地球科学学院,北京100049)
黄、东海陆架2011年秋季活体浮游有孔虫生态分布
吕红红1,2,向荣1*
(1.中国科学院南海海洋研究所中国科学院边缘海地质重点实验室,广东广州510301;2.中国科学院大学地球科学学院,北京100049)
现代活体浮游有孔虫的生态研究是其古环境重建应用的重要基础。根据黄、东海陆架2011年秋季采集的20个垂直浮游拖网样品,分析了该海域浮游有孔虫的秋季生态分布特征。结果表明,黄海秋季基本上没有浮游有孔虫的出现。东海共发现13种活体浮游有孔虫,主要优势属种依次为Globigerinoides sacculifer、Pulleniatina obliquiloculata、Globigerina bulloides、Neogloboquadrina dutertrei和Globigerinoides ruber。浮游有孔虫丰度整体上呈现东南高,西北低的分布格局,这种分布格局反映了浮游有孔虫在黄、东海陆架区的分布主要受外海水影响强弱控制。浮游有孔虫主要属种在东海陆架呈现明显的区域分布差异:暖水种G.sacculifer是秋季陆架海区的主要优势种,其分布格局与总丰度基本一致,相对含量从南至北、从东至西逐渐降低,主要受区域表层海水温度变化的控制。G.bulloides与G.sacculifer呈相反的含量分布变化,其高含量主要出现在东海中陆架,从北往南逐渐降低,此外,在闽浙沿岸也有较高含量,表明了温度和生产力是影响G.bulloides在黄、东海陆架分布的主要因素。秋季P.obliquiloculata和N.dutertrei的高含量主要出现在东海南部中陆架区和济州岛西南黄海暖流影响区,可能受暖水与生产力的共同制约。
黄海;东海;浮游拖网;浮游有孔虫;区域分布;海洋环境
1 引言
浮游有孔虫是全球海洋中广泛分布的一类钙质原生动物,其钙质壳体能够在海洋沉积物中大量保存,作为表层海洋环境变化的一种主要记录载体,被广泛应用于古海洋环境重建研究,具有不可替代的重要作用。浮游有孔虫的区域分布特征、栖息深度习性以及季节性变化等生态资料是进行海洋古环境重建的重要基础。随着高分辨率古海洋研究的不断开展,古海洋学者越发意识到浮游有孔虫生态特征对于区域古海洋环境记录解释的重要性。由于不同海区地理位置不同所导致的海洋物理参数的差异,势必会造成浮游有孔虫在不同海区的生物地理区系或生态差异,因此有必要针对不同海区开展相应的生态调查研究。近年来在全球许多海区都开展了有关现代浮游有孔虫生态的调查研究[1—5]。
黄、东海陆架现代浮游有孔虫生态方面的资料迄今为止基本空白。除了Yamasaki和Oda[6]曾通过沉积物捕获器对东海陆架进行过3个站位的短期调查研究外,其他对黄、东海陆架浮游有孔虫的认识主要来自于表层沉积物的分析[7—10]。然而,沉积物中的有孔虫组合只能反映区域分布变化,无法详细了解主要浮游有孔虫属种的季节和深度变化信息。此外由于陆架残留沉积的存在,导致表层沉积物中的浮游有孔虫不一定是现代海洋环境下的产物。因此黄、东海陆架现代浮游有孔虫生态调查方面的工作亟待开展。本文拟通过黄、东海陆架秋季20个站位23个垂直浮游拖网样品的浮游有孔虫分析,结合现代水文观测资料,探讨秋季陆架海区现代浮游有孔虫的分布规律及其影响因素,为浮游有孔虫在陆架古环境重建应用提供基础生态资料。
2 材料与方法
本文研究材料来自于“我国陆架海生态环境演变过程、机制及未来变化趋势预测”973项目2011年黄、东海陆架秋季调查航次。样品采用浮游有孔虫垂直拖网采集,网口直径65 cm,网眼直径63μm,每个站位按一层采集,个别站位按两层采集,为保证获取足够的有孔虫,部分站位样品为多网合并样品。采集样品在船上用5%福尔马林溶液进行固定,并加少量四硼酸钠缓冲保存。在20个站位(图1)共获取了23个拖网样品(表1)。

表1 拖网样品采样站点、水深、拖网层位以及取样时间Tab.1 Location,water depth,tow depth and collecting dates of the samples
拖网样品在实验室采用125μm的标准网筛进行冲洗,去除样品中盐分,然后将湿样放置在体视显微镜下将粒级大于125μm的浮游有孔虫挑出,鉴定。根据统计结果计算了各站位的浮游有孔虫丰度及浮游有孔虫属种的相对百分含量。
3 海洋环境背景
黄、东海是中国边缘海重要组成部分,有着广阔的陆架。黄、东海陆架同时受陆地输入和外海大洋的影响。在其西侧,两条世界主要的河流(长江、黄河)为黄、东海陆架提供巨量的淡水与陆源物质供给。在其东侧,强劲的西边界流为其带来了高温、高盐的热带水团。这两种主要水体端元在黄东海环流系统作用下的相互混合形成了现今黄东海的水团体系[13]。黄、东海环流体系东侧由自南而北的黑潮、台湾暖流、对马暖流以及黄海暖流组成,在西侧以自北而南的黄海沿岸流、长江冲淡水、闽浙沿岸流等沿岸流系为主,整体构成了该海区的气旋式环流系统(图1)。黑潮表层水主要位于东海外陆架海区,包括陆坡、冲绳海槽,琉球半岛两侧等。低温低盐的黄海沿岸流、闽浙沿岸流系主要呈带状分布于内陆架海区[13]。台湾暖流与黄海暖流均呈现出明显的季节变化特性,台湾暖流秋季势力相对较弱[14];黄海暖流为一具有补偿性质的洋流,在秋冬季明显增强[15]。

图1 浮游拖网取样站位及环流示意图[11—12]Fig.1 Location of plankton tow samples and circulation sketch map in the study area[11-12]
根据同步观测的温盐资料,表层海水温度(SST)变化范围为18~27℃,整体上呈东南高、西北低的格局,南黄海和东海的分界大体上与21℃等温线一致。表层海水盐度(SSS)变化范围为30~34.5,分布格局与温度变化大体一致(图2)。这种温、盐分布格局反映了外海水团与陆架水团的一个混合状况。
叶绿素浓度被广泛用作指示海洋生产力的变化。前人研究发现黄、东海秋季海水垂直混合较好,叶绿素浓度相对较高,整体呈现由沿岸向东部开放海域逐渐降低趋势。叶绿素高值区(大于1 mg/m3)主要出现在沿岸水和济州岛西南区,低值区为东海中部,浓度低于0.5 mg/m3;东海南部叶绿素浓度大于0.5 mg/m3[16—17]。
4 结果
4.1浮游有孔虫群落组成
通过对黄、东海陆架海区20个站位23个拖网样品分析发现,黄海基本上没有浮游有孔虫出现,东海共鉴定出13种,其中百分含量超过5%的属种有5种,分别为:Globigerinoides sacculifer(34.2%)、Pul-leniatina obliquiloculata(18.7%)、Globigerina bulloides(16.4%)、Neogloboquadrina dutertrei(12.1%)和Globigerinoidesruber(9.9%)。以上5个优势属种占本海区浮游有孔虫群落的90%以上。为了更好对比分析浮游有孔虫在陆架区的分布规律,在研究区域选取了4个断面进行分析,其中A、P、E断面主要观察从外海到陆架浮游有孔虫的群落变化,M断面主要观察从南到北浮游有孔虫群落沿纬向的变化规律。本文计算了研究区及各断面各属种相对含量(图3),其中A断面浮游有孔虫以G.bulloides为主,且该属种在群落中占明显优势;P断面以G.sacculifer为主,此外G.ruber相对含量较研究区及其他断面高;E断面以G.sacculifer、P.obliquiloculata为主,而M断面以G.sacculifer、G.bulloides为主。
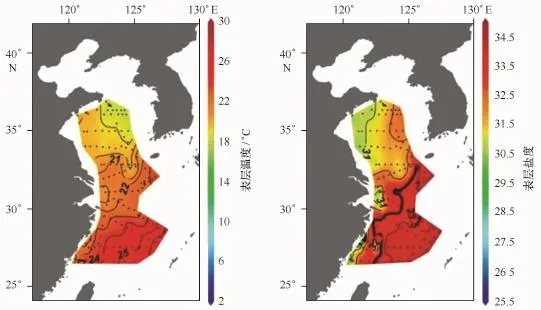
图2 黄、东海陆架海秋季表层温、盐度布变化[18]Fig.2 Distribution of autumn sea surface temperature and sea surface salinity in the Yellow Sea and the East China Sea shelf[18]
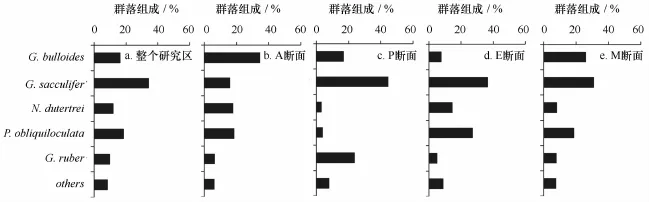
图3 黄、东海陆架研究区及各断面秋季浮游有孔虫群落组成Fig.3 Community compositions of planktonic foraminifera for the whole study area and four sections in the Yellow Sea and the East China Sea shelf
此外,本文计算了黄东海陆架各站位浮游有孔虫简单分异度及体积丰度(图4)。结果表明,本海区各站位浮游有孔虫简单分异度(图4a)在0~12之间,分异度值超过5的区域局限于外陆架海区与30°N以南的中陆架海区;内陆架及30°N以北中陆架海区浮游有孔虫种数寥寥无几,34°N以北海区未见浮游有孔虫分布。浮游有孔虫分异度大致呈现由东南向西北递减的分布格局,黄海及东海内陆架分异度值较低,中外陆架南部分异度值较大。黄东海陆架各站位浮游有孔虫丰度(体积丰度,图4b)与简单分异度分布基本一致,整体呈现东南高,西北低的分布格局。由于离岸的远近和纬度的差异,浮游有孔虫丰度存在明显的差异。丰度超过10枚/m3的区域局限于中陆架外缘及以东的陆架海区;中陆架至内陆架浮游有孔虫丰度逐渐降低,直至为0;黄、东海陆架浮游有孔虫分异度与丰度分布规律在研究区4个断面亦有较好的体现(图4)。其中,A断面自东南向西北分异度值由10逐渐降至为零;丰度大体也呈现东南高,西北低的分布格局,东南部站位丰度均超过10枚/m3,其中A10站位丰度近24枚/m3,而西北部站位丰度为0。P断面分异度与丰度呈现明显的自外海向陆架递减的趋势。E断面东部陆坡区浮游有孔虫丰度较低,该断面陆架区浮游有孔虫分异度与体积丰度自外陆架向内陆架递减趋势。M断面自南至北分异度逐渐减小,但丰度变化不明显。由此可以看出,黄、东海陆架海区及各断面分异度及丰度呈外海高于近岸,南面高于北面的分布格局。
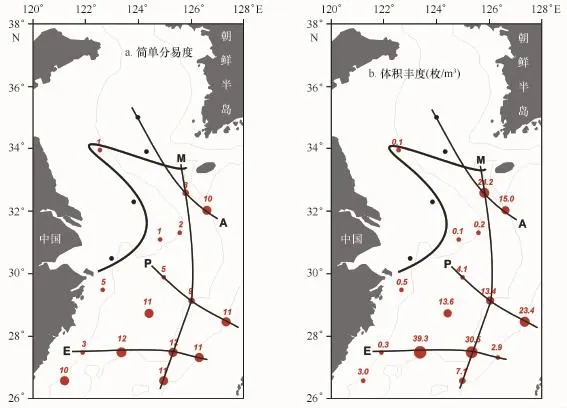
图4 浮游有孔虫简单分异度(a)与体积丰度(b)分布Fig.4 Distributions of species diversity(a)and abundances(b)of planktonic foraminifera
4.2优势属种分布特征
黄、东海陆架海区浮游有孔虫主要属种分布存在区域性差异。图5展现了秋季群落组成以及5个主要优势属种的相对丰度分布情况。G.sacculifer是秋季陆架海区主要优势属种,该属种在东海陆架广泛分布(图5b),东海大部分站位该属种相对丰度大于30%,其中外陆架边缘海区该属种相对丰度超过50%,仅济州岛泥质区相对丰度为10%~20%,陆架海区G.sacculifer相对含量分布格局呈东南向西北逐渐降低趋势。G.ruber含量较G.sacculifer普遍要低,G.ruber含量在10%左右,是该区优势属种之一。高含量区为东海中部中外陆架海区,最高值达50%以上,沿岸流与暖流影响区域该属种相对丰度均较低。
典型冷水种G.bulloides占研究区群落的16%左右,研究区大部分站位该属种含量低于10%(图5e),其高含量区主要出现在东海中陆架,可高达近50%,此外在闽浙沿岸也有较高含量。其丰度分布整体呈现从北往南逐渐降低的分布格局,与G.sacculifer分布格局相反。
P.obliquiloculata与N.dutertrei为次表层水种,主要生活在50~100 m水深[19]。秋季陆架P.obliquiloculata与N.dutertrei分布格局基本一致,高含量区主要出现在东海南部中陆架区和济州岛西南黄海暖流影响区。研究区秋季P.obliquiloculata含量较高(图5d),相对含量高的可达30%以上,个别站位相对含量低于10%。而N.dutertrei秋季含量相对较低,仅占群落12%左右。除了济州岛泥质区与东海南部中陆架区个别站位相对丰度超过10%以外,其他区域含量均较低。
除此之外,A、P、E与M断面(图3 b-e,图5)亦很好反应了有孔虫优势属种分布状况。从E-P-A断面,随着纬度的升高,SST逐渐降低,热带-亚热带种G.sacculifer含量呈减少趋势;冷水种G.bulloides含量逐渐增大。从外海到近岸,冷水种逐渐增加,热带-亚热带种逐渐减少。生产力较高的A断面与E断面中部N.dutertrei、P.obliquiloculata含量较高。位于沿岸水与暖流水之间的P断面G.ruber(23.6%)含量最高。这与整个研究区分布状况基本一致。与此同时,M断面自南向北,浮游有孔虫G.sacculifer的含量逐渐降低,而G.bulloides含量逐渐增加。
5 浮游有孔虫分布与海洋环境
黄、东海陆架海区秋季浮游有孔虫具明显的区域分布特征。浮游有孔虫分布与海洋环境关系密切[20]。本文根据同步实测的CTD温盐资料探讨黄东海陆架海区现代浮游有孔虫与海洋环境的关系。从图6可以看出,浮游有孔虫丰度分布与表层温、盐变化呈正相关(相关系数分别为R2=0.4与R2= 0.56),这可能表明来自外海的高温高盐水相比于陆架低盐水更有利于浮游有孔虫生存。从外陆架到内陆架,外海水影响强度逐渐降低,由外陆架外海水,到陆架水与外海水的混合水,直至内陆架低温低盐陆架水,海水温、盐逐渐下降,浮游有孔虫丰度逐渐降低。黄海为半封闭海域,海水与外界交流较少,自黄海向渤海,外海水影响逐渐减小。黄海34°N以北海区未见浮游有孔虫,黄海其他海域浮游有孔虫较少(图4),这比郑执中和郑守仪[8]在黄、东海表层沉积物中浮游有孔虫最北出现纬度(35°30′N)要低,之所以造成这种差异,有可能是因为本文仅对秋季一个季节数据的研究,有待其他季节的数据加以验证;亦有可能是表层沉积物中有残留沉积物,残留沉积物中的浮游有孔虫非现代浮游有孔虫,从而使得出现浮游有孔虫的纬度比现代浮游有孔虫纬度要高。东海内浮游有孔虫丰度整体呈现东南高,西北低,自东南向西北递减的分布格局(图4)。这一分布格局与表层海水温、盐分布格局(图2)基本一致,可能反应了浮游有孔虫在黄东海陆架区的分布主要受外海水影响强弱的控制。
与总丰度相似,黄、东海陆架浮游有孔虫优势属种的分布与海洋环境亦有较好的相关性。具体表现为,暖水种G.sacculifer主要生活在50 m以上表层海水,其最佳生活水温为20~29℃[21]。该属种是黄、东海秋季陆架海区主要优势属种,其分布与总丰度基本一致。琉球半岛两侧浮游有孔虫捕获器样品中,G.sacculifer在秋季丰度最高,此外海水温度较高的夏季、春季表层该属种丰度亦较高,且在该海区G.sacculifer最佳生活SST为25~26℃,该属种在琉球半岛两侧的分布与SST密切相关[10]。G.sacculifer相对含量分布与表层海水温度分布格局基本一致,整体呈现由东南向西北逐渐降低的趋势。从图6可以看出,该属种相对含量与海水温、盐变化呈正相关,且相关性较好,相关系数分别为R2=0.7和R2=0.44,这可能表明G.sacculifer相对含量随表层海水温度升高而升高(图6),可能反映了该属种分布受表层海水温度变化控制,与捕获器[6]和表层沉积物[9]研究结果较为一致。
G.bulloides为典型冷水种,栖息环境水温相对较低,最佳SST为(13.4±7.8)℃[22—23],喜生活在高营养盐和生产力环境。该属种在北大西洋[24]、阿拉伯海以及中赤道太平洋[25]等海区被视为高生产力的上升流指示种。张在秀等[26]将G.bulloides作为沿岸环境指示种对东海泥质沉积区古环境进行相关研究;赵泉鸿等[27]在东海陆架泥质区全新世古环境研究中,把G.bulloides作为上升流指示种对东海陆架古上升流进行研究。本文研究区域G.bulloides相对含量呈现自北至南逐渐降低趋势,与G.sacculifer分布相反,从图6可看出,G.bulloides相对含量分布与SST和SSS变化呈负相关(图6,相关系数分别为R2=0.39和R2=0.38),这可能表明G.bulloides相对含量随表层海水温度升高明显降低。但闽浙沿岸温度较高,G.bulloides相对含量却较高,研究表明,闽浙沿岸秋季受沿岸上升流影响,初级生产力较高[28],G.bulloides相对含量较高,这表明该属种可能还受生产力等因素控制。由此可知,黄、东海陆架区G.bulloides分布可能主要受表层海水温度和生产力共同控制。
次表层水中P.obliquiloculata与N.dutertrei常被作为黑潮指示种[29—30]。然而从我们秋季活体浮游有孔虫的分布看,除了受黄海暖流影响的济州岛西南区这两个属种有较高的含量分布外,在东部黑潮主流轴区P.obliquiloculata与N.dutertrei丰度均较小,这表明以上两个属种除了受黑潮暖水影响以外,可能还受其他因素控制。研究表明,P.obliquiloculata与N.dutertrei在高生产力区含量较大。Watkins等[25]对中赤道太平洋区拖网样品中浮游有孔虫研究发现在热带沿岸上升流区,P.obliquiloculata与N.dutertrei较多;Fairbanks等[31]在巴拿马海盆活体浮游有孔虫调查研究中发现生产高的海区N.dutertrei丰度较大;在北太平洋西北部,Kuroyanagi等[23]发现N.dutertrei主要出现在高生产力的上升流区。在我国南海南沙海区,P.obliquiloculata与N.dutertrei高通量时期,叶绿素浓度较高,而SST并非高值期[32]。可见,该属种分布不仅受温度控制,还受生产力等因素控制。秋季济州岛西南海区受黄海暖流影响,东海南部中陆架受台湾暖流影响,海水温盐较高,同时这两个区域秋季叶绿素a浓度反映的生产力也较高[16,33],有利于两个属种的生长。因此,我们推断研究区P.obliquiloculata与N.dutertrei的分布可能受黑潮相关暖水与生产力的共同制约。

图6 浮游有孔虫丰度与陆架海温、盐的相关性Fig.6 Relationship between the planktonic foraminiferal abundance and water temperature and salinity
广温性属种G.ruber主要生活在50 m以上表层海水,平均生活水温为23.8℃[19]。东海捕获器样品中,该属种主要出现在SST较高的夏秋季节[6],在南海G.ruber夏季丰度较高[34]。在5°N以北和5°S以南温暖、贫营养盐海区G.ruber含量较高[25]。G.ruber分布与营养盐高低、食物多少、光照强弱等因素息息相关[31,35]。G.ruber在研究区主要分布在中部中外陆架,沿岸流影响区以及暖流影响区相对含量均较低。这表明,表层海水温度过高或过低均不适合G.ruber生长,陆架水与暖流水混合的中部中外陆架区秋季SST为(23±1)℃(图2),该水温状况有利于G.ruber生长,其相对含量较大。而陆架水区海水温度较低,且营养盐较高,可能不利于该属种的生长。此外广温性属种在两水团混合区竞争力更大,相对含量较大。
由于本文仅根据秋季1个航次的样品来研究陆架海区的浮游有孔虫分布,因此所得的认识肯定不全面,与表层沉积物中的有孔虫组合肯定会存在明显的差异,还需要其他不同季节的调查结合起来进行研究。
6 结语
通过对黄、东海陆架海区秋季浮游有孔虫垂直拖网样品的分析,获得以下认识:
(1)黄海秋季基本没有浮游有孔虫出现。东海陆架海区秋季共鉴定出13个属种,其中主要优势属种依次为:Globigerinoides sacculifer、Pulleniatina obliquiloculata、Globigerina bulloides、Neogloboquadrina dutertrei和Globigerinoides ruber。A断面浮游有孔虫以G.bulloides为主,且该属种在群落中占明显优势;P断面以G.sacculifer为主,此外G.ruber相对含量较研究区及其他断面高;E断面以G.sacculifer、P.obliquiloculata为主,而M断面以G.sacculifer、G.bulloides为主。
(2)黄、东海陆架海区浮游有孔虫丰度整体呈现东南高,西北低分布格局,这种分布格局可能反应了浮游有孔虫在黄、东海陆架海区的分布主要受外海水影响强弱控制。自外海至近岸,受外海水影响逐渐减小,浮游有孔虫分异度与丰度亦相应降低。黄海为半封闭海域,海水与外界交流较少,自黄海向渤海,外海水影响逐渐减小。黄海34°N以北海区未见浮游有孔虫,黄海其他海域浮游有孔虫寥寥无几。
(3)浮游有孔虫主要属种在东海陆架呈现明显的区域分布差异:暖水种G.sacculifer是秋季陆架海区的主要优势属种,其分布格局与总丰度一致,相对含量从东至西,从南至北逐渐降低,主要受区域表层海水温度变化的控制。G.bulloides与G.sacculifer呈相反的含量分布变化,其含量主要出现在东海中陆架,从北往南逐渐降低,此外在闽浙沿岸也有较高含量,表明温度和生产力是影响G.bulloides在黄、东海陆架分布的主要因素。P.obliquiloculata与N.dutertrei的高含量主要出现在东海南部中陆架区和济州岛西南黄海暖流影响区,可能受黑潮相关暖水与生产力共同制约。广温性属种G.ruber高含量主要出现在研究区的中部中外陆架。
致谢:衷心感谢国家重点基础研究发展计划(973计划)“我国陆架海生态环境演变过程、机制及未来变化趋势预测”秋季航次获取的温盐数据。感谢杜恕环老师为本研究采集了浮游有孔虫拖网样品。
[1]刘芳,向荣.现代浮游有孔虫生态研究进展[J].微体古生物学,2010,27(4):366-375.Liu Fang,Xiang Rong.Advances in the study of modern planktonic foraminifera[J].Acta Micropalaeontologica Sinica,2010,27(4):366-375.
[2]Oda M,Yamasaki M.Sediment trap results from the Japan trench in the Kuroshio domain:Seasonal variations in the planktonic foraminifera flux[J].The Journal of Foraminiferal Research,2005,35(4):315.
[3]Wilke I,Meggers H,Bickert T.Depth habitats and seasonal distributions of recent planktonic foraminifers in the Canary Islands region(29°N)based on oxygen isotopes[J].Deep-Sea ResearchⅠ,2009,10:1-18.
[4]Eguchi N O,Ujiie H,Kawahata H,et al.Seasonal variations in planktonic foraminifera at three sediment traps in the subarctic,transition and subtropical zones of the central north Pacific Ocean[J].Marine Micropaleontology,2003,48(1/2):149-163.
[5]丁旋,王汝建,张海峰,等.北冰洋马克洛夫海盆现代浮游有孔虫深度分布及其生态与碳氧同位素特征[J].科学通报.2014,59(13):1230-1241.Ding Xuan,Wang Rujian,Zhang Haifeng,et al.Distribution,ecology,and oxygen and carbon isotope characteristics of modern planktonic foraminifers in the Makarov Basin of the Arctic Ocean[J].Chinese Science Bulletin,2014,59(13):1230-1241.
[6]Yamasaki M,Oda M.Sedimentation of planktonic foraminifera in the East China Sea:evidence from a sediment trap experiment[J].Micropaleontology,2003,49:3-20.
[7]郑执中,郑守仪.黄海和东海浮游有孔虫[J].海洋与湖沼,1960,3:125-164.Cheng Tsi-chung,Cheng Sau-yee.The planktonic foraminifera of the Yellow Sea and the East China Sea[J].Oceanologia et Limnologia Sinica,1960,3:125-164.
[8]郑执中,郑守仪.黄海和东海浮游有孔虫生态的研究[J].海洋与湖沼,1962,4:60-86.Cheng Tsi-chung,Cheng Sau-yee.On the ecology of the planktonic foraminifera of the Yellow Sea and the East China Sea[J].Oceanologia et Limnologia Sinica,1962,4:60-86.
[9]汪品先,张纪军,赵泉鸿,等.东海低质中的有孔虫和介形虫[M].北京:海洋出版社,1988:1-438.Wang Pinxian,Zhang Jijun,Zhao Quanhong,et al.Foraminifera and Ostracoda in Bottom Sediments of the East China Sea[M].Beijing:China O-cean Press,1988:1-438.
[10]Xu X D,Yamasaki M,Oda M,et al.Comparison of seasonal flux variations of planktonic foraminifera in sediment traps on both sides of the Ryukyu Islands,Japan[J].Marine Micropaleontology,2005,58:45-55.
[11]王利波,杨作升,张荣平,等.南黄海中部泥质区ZY2孔6200年以来的海表温度记录及黄海暖流变化的影响[J].科学通报,2011,56(15):1213-1220.Wang Libo,Yang Zuosheng,Zhang Rongping,et al.Sea surface temperature records of core ZY2 from the central mud area in the South Yellow Sea during last 6200 years and related effect of the Yellow Sea Warm Current[J].Chinese Science Bulletin,2011,56(15):1213-1220.
[12]Zhou C Y,Dong P,Li G X.Hydrodynamic processes and their impacts on the mud deposit in the Southern Yellow Sea[J].Marine Geology,2015,360:1-16.
[13]Li G,Han X,Yue S,et al.Monthly variations of water masses in the East China Seas[J].Continental Shelf Research,2006,26:1954-1970.
[14]Qi J F,Yin B S,Zhang Q L,et al.Analysis of seasonal variation of water masses in East China Sea[J].Chinese Journal of Oceanology and Limnology,2014,32(4):958-971.
[15]Xu L,Wu D,Lin X,et al.The study of the Yellow Sea warm current and its seasonal variability[J].J Hydrodyn Ser B,2009,21:159-165.
[16]宁修仁,刘子琳,史君贤.渤、黄、东海初级生产力和潜在渔业生产量的评估[J].海洋学报,1995,17(3):73-84.Ning Xiuren,Liu Zilin,Shi Junxian.Estimation of ocean primary productivity and the potential fisheries production of the Bohai Sea,the Yellow Sea and the East China Sea[J].Haiyang Xuebao,1995,17(3):73-84.
[17]李国胜,王芳,梁强,等.东海初级生产力遥感反演及其时空演化机制[J].地理学报,2003,58(4):483-493.Li Guosheng,Wang Fang,Liang Qiang,et al.Estimation of ocean primary productivity by remote sensing and introduction to spacialtemperal variation mechenism for the East China Sea[J].Journal of Geographical Sciences,2003,58(4):483-493.
[18]Quan Q,Mao X Y,Yang X D,et al.Seasonal variations of several main water masses in the southern Yellow Sea and East China Sea in 2011[J].Oceanic and Coastal Sea Research,2013,12(4):524-536.
[19]BéA W H.An ecological,zoogeographic and taxonomic review of recent planktonic foraminifera[G]//Ramsay A T S.Oceanic Micropaleontology.Academic press,London,1977:1-100.
[20]向荣,陈木宏,张兰兰,等.南海北部秋季活体浮游有孔虫的组成与分布[J].中国地质大学学报,2010,35(1):1-10.Xiang Rong,Chen Muhong,Zhang Lanlan,et al.Compositions and distribution of living planktonic foraminifera in autumn waters of the northern South China Sea[J].Earth Science,Journal of China University of Geosciences,2010,35(1):1-10.
[21]Fabien L,Laurent L,Elisabeth M,et al.Modelling the temperature dependent growth rates of planktonic foraminifera[J].Marine Micropaleontology,2009,70:1-7.
[22]BéHutson.Ecology of planktonic foraminifera and biogeographic patterns of life and fossil assemblages in the Indian Ocean[J].Micropaleontology,1977,23:369-414.
[23]Kuroyanagi A,Kawahata H,Nishi H,et al.Seasonal changes in planktonic foraminifera in the northwestern North Pacific Ocean:sediment trap experiments from subarctic and subtropical gyres[J].Deep-sea ResearchⅡ,2002,49:5627-5645.
[24]Gupta A K,Anderson D M,Overpeck J T.Abrupt changes in the Asian southwest monsoon during the Holocene and their links to the North Atlantic Ocean[J].Nature,2003:354-357.
[25]Watkins J M,Mix A C,Wilson J.Living planktic foraminifera in the central tropical Pacific Ocean:articulating the equatorial‘cold tongue'during La Nifia,1992[J].Marine Micropaleontology,1998,33:157-174.
[26]张在秀,赵泉鸿,陈荣华,等.东海泥质沉积区8千年来的有孔虫Mg/Ca和δ18O记录及其古环境意义[J].海洋地质与第四纪地质,2010,30(3):79-86.Zhang Zaixiu,Zhao Quanhong,Cheng Ronghua,et al.Foraminiferal Mg/Ca andδ18O records in the mud aera of the East China Sea during the last 8000 years and their palaeoenvironmental implications[J].Marine Geology&Quaternary Geology,2010,30(3):79-86.
[27]赵泉鸿,翦知湣,张在秀,等.东海陆架泥质沉积区全新世有孔虫和介形虫及其古环境应用[J].微体古生物学报,2009,26(2):117-128.Zhao Quanhong,Jian Zhimin,Zhang Zaixiu,et al.Holocene Benthic Foraminifera and Ostracoda from the shelf mud aera of the East China Sea and their palaeoenvironmental implications[J].Acta Micropalaeontologica Sinica,2009,26(2):117-128.
[28]经志友,齐义泉,华祖林.闽浙沿岸上升流及其季节变化的数值研究[J].河海大学学报,2007,35(4):464-470.Jing Zhiyou,Qi Yiquang,Hua Zulin.Numerical study on upwelling and its seasonal variation along Fujian and Zhejiang coast[J].Journal o f Hohai University,2007,35(4):464-470.
[29]Xu X D,Oda M.Surface-water evolution of the eastern East China Sea during the last 36,000 years[J].Marine Micropaleontology,1999,156:285 -304.
[30]Jian Z,Wang P,Saito Y,et al.Holocene variability of Kuroshio Current in the Okinawa Trough,northwestern Pacific Ocean[J].Earth and Planetary Science Letters,2000,184:305-319.
[31]Fairbanks R G,Sverdlove M,Free R,et al.Vertical distribution and isotopic fractionation of living planktonic foraminifera from the Panama Basin[J].Nature,1982,298:841-844.
[32]万随,翦知湣,成鑫荣,等.南沙海区浮游有孔虫通量及其壳体化学性质的季节变化[J].中国科学:地球科学,2010,40(7):881-892.Wan Sui,Jian Zhimin,Cheng Xinrong,et al.Seasonal variations in planktonic foraminiferal flux and the chemical properties of their shells in the southern South China Sea[J].Science China:Earth Science,2010,40(7):881-892.
[33]Li G S,Gao P,Wang F et al.Estimation of ocean primary productivity and its spatio-temporal variation mechanism for East China Sea based on VGPM model[J].Journal of Geographical Sciences,2004,14(1):32-40.
[34]Lin H L,Hsieh H Y.Seasonal variations of modern planktonic foraminifera in the South China Sea[J].Deep-Sea Research PartⅡ,2007,54:1634 -1644.
[35]Kuroyanagi A,Kawahata H.Vertical distribution of living planktonic foraminifera in the seas around Japan[J].Marine Micropaleontology,2004,53:173-196.
Distribution of living planktonic foraminifera on the shelf of the Yellow Sea and the East China Sea in autumn,2011
Lyu Honghong1,2,Xiang Rong1
(1.Key Laboratory of Marginal Sea Geology,South China Sea Institute of Oceanology,Chinese Academy of Sciences,Guangzhou 510301,China;2.College of Earth Science,University of Chinese Academy of Sciences,Beijing 100049,China)
The knowledge of ecology of modern living planktonic foraminifera is very important for their application in paleoenvironmental reconstruction.The distribution of living planktonic foraminifers has been analyzed on 20 plankton tow samples collected from the shelf of the Yellow Sea and the East China Sea in Autumn,2011.The results indicate that planktonic foraminifera is almost absent in autumn in the Yellow Sea.Thirteen planktonic foraminiferal species occurred in the East China Sea,dominated by Globigerinoides sacculifer,Pulleniatina obliquiloculata,Globigerina bulloides,Neogloboquadrina dutertrei and Globigerinoides ruber.The abundance of planktonic foraminifera shows a general pattern,with high abundance in the southeast and low values in the northwest of the study area,which may indicate that the distribution of foraminifers in the study area is mainly controlled by the influence of the open ocean water.The major species of planktonic foraminifera shows apparent different distribution in the East China shelf seas.G.sacculifer,the preponderant species of the shelf seas in autumn,shows a similar distribution pattern with the total abundance.Its relative percentage gradually decreased from south to north,and from east to west,suggesting the control of the variation by regional surface sea temperature.In contrast to the distribution of G.sacculifer,high percentage of G.bulloides mainly occurred in the middle shelf of the East China Sea,and its relative abundance gradually decreased from north to south,and high percentage also occurred in the inner shelf off Min-Zhe coast,suggesting that both temperature and productivity are principal factors controlling the distribution of G.bulloides in the study area.High abundance of P.obliquiloculata and N.dutertrei mainly appeared in the middle shelf of the southern East China Sea and southwest off Cheju Island,possibly modulated by the joint influence of warm water and productivity.
the Yellow Sea;the East China Sea;plankton tow;planktonic foraminifera;spatial distribution;marine environment
吕红红,向荣.黄、东海陆架2011年秋季活体浮游有孔虫生态分布[J].海洋学报,2016,38(2):93—103,
10.3969/j.issn.0253-4193.2016.02.009
Lyu Honghong,Xiang Rong.Distribution of living planktonic foraminifera on the shelf of the Yellow Sea and the East China Sea in autumn,2011[J].Haiyang Xuebao,2016,38(2):93—103,
10.3969/j.issn.0253-4193.2016.02.009
Q958
A
0253-4193(2016)02-0093-11
2015-09-22;
2015-11-16。
国家重点基础研究发展计划973项目(2010CB428901);国家自然科学基金项目(41476040,91228207)。
吕红红(1989—),女,江西省乐平市人,主要从事海洋沉积古环境研究。E-mail:lvhonghong13@mails.ucas.ac.cn
向荣(1972—),男,湖南省洞口县人,研究员,主要从事微体古生物与海洋沉积古环境研究。E-mail:rxiang@scsio.ac.cn
