有氧运动对胰岛素抵抗小鼠visfatin表达的影响
2016-11-24李晶董荣娜于德民
李晶 董荣娜 于德民
·论著·
有氧运动对胰岛素抵抗小鼠visfatin表达的影响
李晶 董荣娜 于德民
目的探讨高脂饮食诱导的胰岛素抵抗(IR)小鼠visfatin表达的改变及有氧运动对visfatin表达及脂代谢的影响。方法雄性C57BL/6小鼠24只,按随机数字表随机分为普通饮食组(n=12)和高脂饮食组(n=12),后者喂养8周高脂饲料建立IR模型,再各自分为普通饮食对照组(n=6)、普通饮食运动组(n=6)、高脂饮食对照组(n=6)、高脂饮食运动组(n=6)。两个运动组进行6周跑台训练,各组饮食不变。14周后ELISA检测小鼠血清visfatin,Western印迹检测腓肠肌、肝脏及睾周脂肪visfatin表达。结果8周后,与普通饮食组相比,高脂饮食组空腹胰岛素水平升高,血糖曲线下面积增加(t=6.32,17.44,P均<0.01)。14周后,高脂饮食运动组血清visfatin水平低于高脂饮食对照组,而普通饮食运动组血清visfatin水平高于普通饮食对照组(F=43.59,P<0.05)。高脂饮食运动组骨骼肌、肝脏、内脏脂肪组织中visfatin 蛋白表达水平较高脂饮食对照组降低(F=29.33~85.17,P均<0.05),普通饮食运动组骨骼肌visfatin表达较普通饮食对照组升高(F=52.79,P<0.05),但肝脏、内脏脂肪组织visfatin蛋白表达在普通饮食组间无明显变化(P均>0.05)。结论高脂饮食可以增加IR模型动物visfatin水平,有氧运动能有效下调visfatin的表达,改善脂代谢紊乱。
有氧运动;胰岛素抵抗;内脂素;脂代谢
胰岛素抵抗(IR)是诸多代谢性疾病共同的病理生理基础。改善不良的饮食习惯和进行积极地有氧运动能够有效的预防IR的发生[1]。Visfatin是一种脂肪细胞因子,有类胰岛素的作用,在机体内的变化与IR相关[2]。有研究提示高脂饮食可以导致肥胖及IR的发生,同时visfatin水平有升高趋势。而有氧运动对肥胖及IR有一定的改善作用,但目前有关visfatin在IR状态下的变化及有氧运动干预对其影响的相互作用关系仍不完全清楚。本研究探讨有氧运动对高脂饮食诱导的C57BL/6 IR小鼠血清、骨骼肌、肝脏及内脏组织中visfatin水平的影响。
1 材料与方法
1.1 动物饲养与分组 选用24只6周龄雄性C57BL/6小鼠,体重(24.60±0.75)g[购自军事医学科学院实验动物中心,许可证号SCXK-(军)2012-0004,批号0026037],温度22~25℃,湿度30%~40%,光照模拟昼夜交替养殖。1周适应性喂养后,按动物体重编号,应用随机数字表将动物随机分为高脂饮食组(n=12)和普通饮食组(n=12只),分别给予高脂饲料和普通饲料喂养8周。8周后通过检测空腹胰岛素水平和口服葡萄糖耐量试验(OGTT)来评判IR成模情况,所有动物模型均成功造模。随后将两组动物再根据运动情况分别分为两个亚组:高脂饮食对照组(n=6),继续高脂饮食;高脂饮食运动组(n=6),在高脂饮食喂养同时每天进行60 min的跑台训练;普通饮食对照组(n=6),持续普通饮食喂养;普通饮食运动组(n=6),每天进行60 min的跑台训练,再观察6周。
1.2 饲料配方 普通饮食组小鼠喂以普通饲料(购自天津实验动物中心);高脂饮食组喂以成分为蛋白质20%、碳水化合物35%、脂肪45%配方的高脂饲料(购于中国医学科学院实验动物研究所)。
1.3 运动方案 适应训练1周后,逐渐增加强度,使动物适应,直至达到预定强度。为期6周,强度为75%VO2max,时间为60 min/次,频次为5次/周。
1.4 标本采集 在所有干预结束后,将小鼠禁食12 h,乙醚麻醉,内眦静脉取血留取血样,分离血清,置于-20℃保存。随后,断颈处死动物,分离腓肠肌、肝脏及睾周脂肪组织,置于液氮保存。
1.5 检测指标
1.5.1 OGTT及胰岛素的测定 喂养8周后,将小鼠禁食14~16 h,尾静脉取血。采用血糖仪(罗氏公司)测定小鼠空腹(0 min)血糖。然后给小鼠(10 μl/g)胃内灌注20%葡萄糖溶液(Sigma公司),分别在15,30,60,90,150,180 min进行尾静脉取血检测血糖。采用ELISA法检测胰岛素水平(R&D公司)。
1.5.2 血甘油三酯、胆固醇及visfatin水平测定 在所有干预结束后将小鼠禁食14~16 h,鼠内眦静脉取血,离心后取血清应用酶法测定血甘油三酯、低密度脂蛋白-胆固醇(LDL-C)、高密度脂蛋白-胆固醇(HDL-C)。采用ELISA法检测visfatin水平(R&D公司),将标本和不同浓度标准品加入相应孔中,37℃孵育90 min,洗板4次,加入抗体工作液,孵育60 min,洗板4次,加入酶结合物工作液,孵育30 min,洗板4次,加入显色剂和终止液,混匀后即刻测量OD450值(5 min内),生成标准曲线, 根据OD值由标准曲线换算visfatin含量。
1.5.3 Western 印迹检测骨骼肌、肝脏、脂肪visfatin蛋白表达 提取腓肠肌、肝脏、睾周脂肪蛋白,经分光光度仪定量后在垂直电泳仪(BIO-RAD)上用等量(30 μg)蛋白质样品经7.5%SDS-PAGE分离后,转移于PVDF膜(Millipore)上(110 V,1 h)。PVDF膜经5%脱脂奶粉封闭1 h,再用5%牛血清白蛋白(BSA)按1∶1 000(Santa Cruz,Rabbit anti-visfatin)稀释一抗,4℃孵育过夜,经1×TBST充分洗涤,再用5%脱脂奶粉按照1∶5 000(Invitrogen,HRP 标记羊抗兔多克隆IgG)稀释二抗,室温下将PVDF膜用二抗孵育1 h,1×TBST充分洗涤后,使用E-CL Plus试剂盒发光显影,胶片曝光,扫描定量各信号条带的相对灰度值。

2 结果
2.1 IR动物模型的建立 通过8周的高脂饲料喂养,高脂饮食组空腹胰岛素水平显著高于普通饮食组(t=6.32,P<0.05),见图1;高脂饮食组OGTT后血糖曲线下面积较普通饮食组增加54.31%(t=17.44,P<0.05),见图2。提示8周的高脂饮食喂养已经成功地建立了IR动物模型。

注:FINS:空腹胰岛素图1 喂养8周后小鼠的FINS水平
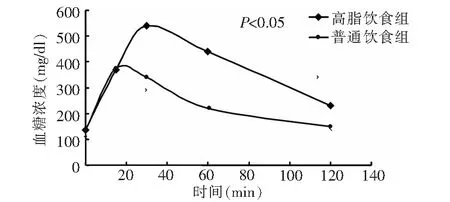
注:OGTT:口服葡萄糖耐量试验图2 喂养8周后小鼠的OGTT曲线图
2.2 各组小鼠血脂水平比较 如表1所示,高脂饮食运动组甘油三酯和LDL-C水平均较高脂饮食对照组显著降低(P均<0.01),而HDL-C水平无明显变化(P>0.05);同时,普通饮食对照组及运动组甘油三酯和LDL-C水平均低于高脂饮食对照组(P均<0.05);普通饮食运动组甘油三酯水平较普通饮食对照组降低(P<0.01)。普通饮食运动组LDL-C水平较普通饮食对照组无明显改变(P>0.05),但HDL-C水平降低(P<0.05)。
2.3 各组小鼠血清visfatin含量变化 高脂饮食运动组visfatin水平[(400.34±15.83)μg/L]较高脂饮食对
照组[(450.11±16.09)μg/L]降低,而普通饮食运动组[(331.09±5.48)μg/L]较普通饮食对照组[(283.42±43.89) μg/L]升高(F=43.59,P=0.015)。
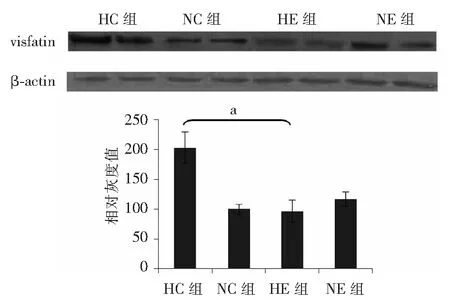
2.4 各组小鼠骨骼肌visfatin表达变化 由图3可知, 高脂饮食运动组较高脂饮食对照组骨骼肌中visfatin的表达下降(F=52.79,P<0.05);普通饮食运动组visfatin的表达比普通饮食对照组增加(F=52.79,P<0.05)。

注:HC组:高脂饮食对照组;HE组:高脂饮食运动组;NC组:普通饮食对照组;NE组:普通饮食运动组;与普通饮食对照组相比,aP<0.05;与高脂饮食对照组相比, bP<0.05图3 各组小鼠骨骼肌visfatin表达的比较
2.5 各组小鼠肝脏visfatin表达变化 由图4可知,高脂饮食运动组较高脂饮食对照组肝脏中visfatin的表达下降(F=29.33,P<0.05);普通饮食运动组与普通饮食对照组之间visfatin表达差别无统计学意义(P>0.05)。
2.6 各组小鼠内脏脂肪visfatin表达变化 由图5可知, 高脂饮食运动组较高脂饮食对照组内脏脂肪中visfatin的表达下降(F=85.18,P<0.05);其在普通饮食运动组中的表达与普通饮食对照组相比,差异无统计学意义(P>0.05)。
表1 各组小鼠血脂水平比较
注:TG:甘油三酯;LDL-C:低密度脂蛋白-胆固醇;HDL-C:高密度脂蛋白-胆固醇;与普通饮食对照组相比,aP<0.05;与高脂饮食对照组相比,bP<0.05
注:HC组:高脂饮食对照组;HE组:高脂饮食运动组;NC组:普通饮食对照组;NE组:普通饮食运动组;与高脂饮食对照组相比,aP<0.05图4 各组小鼠肝脏visfatin表达的比较

注:HC组:高脂饮食对照组;HE组:高脂饮食运动组;NC组:普通饮食对照组;NE组:普通饮食运动组;与高脂饮食对照组相比,aP<0.05图5 各组小鼠内脏脂肪visfatin表达的比较
3 讨论
本研究通过OGTT证实,C57BL/6小鼠经过8周的高脂饮食喂养,反映糖代谢情况的空腹胰岛素水平及血糖的曲线下面积,都可以说明动物模型出现了IR,提示8周的高脂饮食喂养是制备动物模型的有效方法。
研究显示,visfatin能促进葡萄糖合成甘油三酯并诱导其在前脂肪细胞积聚,还能诱导多种脂肪标志物的基因表达,促进前脂肪细胞分化及脂质储存。另外,visfatin可以通过旁分泌途径作用于内脏脂肪组织,促进脂肪组织的分解、合成与积聚。本研究中,小鼠经过8周的高脂饮食喂养,血清LDL-C和甘油三酯水平显著高于普通饮食组,提示此时机体已经出现了脂代谢异常。在6周有氧运动干预后,血清LDL-C和甘油三酯水平也明显降低,表明有氧运动可改善机体的脂代谢异常;但是HDL-C在运动干预前、后变化趋势不明显,提示在动物体内甘油三酯和LDL-C水平可能更容易受到运动干预的调节。另外,visfatin水平与HDL-C呈正相关,与LDL-C、甘油三酯呈负相关,且这种相关性独立于肥胖和IR之外[3]。说明visfatin可能在脂代谢中发挥重要作用。但本研究中没有观察到,visfatin的改变与血脂谱变化的相关性,对于本研究结果与既往研究存在差异的原因,此次研究还难以直接确定,但是前述研究是在新发2型糖尿病患者和糖耐量减低者中观察到的特点,本研究的对象是动物模型,可能是动物与人体存在一定的差异导致了这样的结果,也可能是干预周期相对较短,还不能完全模拟人体具体的变化特点。
本研究结果显示,高脂饮食对照组血清visfatin水平显著高于其他组,说明通过8周的高脂饮食诱导小鼠IR模型建立的过程中,血清visfatin水平呈现升高趋势。
这可能是机体在IR发生后,由于代谢平衡的需要,通过升高血清visfatin来代偿。有研究证实,visfatin不但可以促进胰岛素分泌、抑制胰岛β细胞凋亡,也可以促进胰岛素受体的磷酸化,进而改善糖代谢[4]。
Visfatin的主要功能是具有类胰岛素作用,是NAD+合成限速酶,参与炎性反应和免疫过程的调节[5-7]。它在脂肪组织中高表达,因而可能是与肥胖、IR、糖尿病等代谢疾病相关的一个候选因子[8]。本研究中,随着高脂饮食诱导的IR的形成,visfatin的水平也发生相应改变,由此提示,visfatin可能与肥胖、IR及糖代谢相关。
有氧运动已被证实是预防肥胖、IR等行之有效的方法,能够持续增加肥胖和IR者骨骼肌对胰岛素的敏感性[9]。另外,有氧运动可以增加脂肪酸氧化及增强线粒体氧化酶活性,使骨骼肌对胰岛素的敏感性增强,从而改善IR[10-11]。本研究发现,有氧运动干预可以降低高脂饮食诱导的IR动物模型血清visfatin的水平,但却可以升高普通饮食小鼠的血清visfatin水平。由于血清visfatin可来源于脂肪组织、肝脏、肌肉等,所以血清visfatin的变化来源还难以定位于某一特定组织。此外,经过有氧运动干预,骨骼肌中visfatin的表达变化与血清中的变化趋势一致,而对于高脂饮食组,有氧运动会下调肝脏与内脏脂肪组织中visfatin的表达,但是对于普通饮食组,有氧运动对小鼠肝脏、内脏脂肪中visfatin的表达无明显的调节变化,提示骨骼肌中visfatin表达的变化可能更容易受到有氧运动的调节,或者本研究有氧运动干预的强度和时间长度可以影响骨骼肌visfatin的表达,但还不足以影响肝脏及内脏脂肪组织visfatin的表达,还需进一步完善实验设计,进一步明确其中的原因。
综上所述,高脂饮食可以诱导IR的形成,同时引起visfatin水平的升高。有氧运动能够有效减轻IR,改善机体的脂代谢紊乱,但不是直接通过升高visfatin蛋白表达水平实现的。由此推测,有氧运动可能不是通过提高骨骼肌、肝脏和内脏脂肪的visfatin表达水平,而是通过改善visfatin的作用效能来实现的。基于现有的实验结果,visfatin水平的升高更像是机体应对IR状态的代偿性改变,有氧运动在减轻实验动物IR状态的同时,导致了visfatin表达水平的下调,但visfatin在IR形成过程中具体扮演的角色,还需要进一步研究。
[1] Kurose S,Tsutsumi H,Yamanaka Y,et al. Improvement in endothelial function by lifestyle modification focused on exercise training is associated withinsulin resistance in obese patients[J].Obes Res Clin Pract,2014,8(1):e106-e114. DOI: 10.1016/j.orcp.2012.10.005.
[3] Dogru T,Sonmez A,Tasci I,et al. Plasma visfatin levels in patients with newly diagnosed and untreated type 2 diabetes mellitus and impairedglucose tolerance[J].Diabetes Res Clin Pract,2007,76(1):24-29.
[4] Xiang RL,Mei M,Su YC,et al. Visfatin protects rat pancreatic β-cells against IFN-γ-induced apoptosis through AMPK and ERK1/2 signaling pathways[J].Biomed Environ Sci,2015,28(3):169-177. DOI:10.3967/bes2015.023.
[5] Fukuhara A,Matsuda M,Nishizawa M,et al. Visfatin: a protein secreted by visceral fat that mimics the effects of insulin[J].Science,2005,307(5708):426-430.
[6] Wang T,Zhang X,Bheda P,et al. Structure of Nampt/PBEF/visfatin, a mammalian NAD+biosynthetic enzyme[J].Nat Struct Mol Biol,2006,13(7):661-662.
[7] Jacques C,Holzenberger M,Mladenovic Z,et al. Proinflammatory actions of visfatin/nicotinamide phosphoribosyltransferase (Nampt) involve regulation of insulinsignaling pathway and Nampt enzymatic activity[J].J Biol Chem,2012,287(18):15100-15108. DOI:10.1074/jbc.M112.350215.
[8] Stofkova A. Resistin and visfatin: regulators of insulin sensitivity, inflammation and immunity[J].Endocr Regul,2010,44(1):25-36.
[9] Trevellin E,Scorzeto M,Olivieri M,et al. Exercise training induces mitochondrial biogenesis and glucose uptake in subcutaneous adipose tissue through eNOS-dependent mechanisms[J].Diabetes,2014,63(8):2800-2811. DOI:10.2337/db13-1234.
[10] Liu X,Niu Y,Yuan H,et al. AMPK binds to Sestrins and mediates the effect of exercise to increase insulin-sensitivity through autophagy[J].Metabolism,2015,64(6):658-665.DOI:10.1016/j.metabol.2015.01.015.
[11] Li J,Zhang W,Guo Q,et al. Duration of exercise as a key determinant of improvement in insulin sensitivity in type 2 diabetes patients[J].Tohoku J Exp Med,2012,227(4):289-296.
Effectsofaerobicexerciseontheexpressionofvisfatininmicewithinsulinresistance
LiJing,DongRongna,YuDemin.KeyLaboratoryofHormonesandDevelopment
(MinistryofHealth),TheMetabolicDiseaseHospital&TianjinInstituteofEndocrinology,TianjinMedicalUniversity,Tianjin300070,China
YuDemin,Email:yudemintij@126.com
ObjectiveTo explore the expression of visfatin in mice with insulin resistance (IR) induced by high-fat diet and the effects of aerobic exercise on the expression of visfatin and lipid metabolism.MethodsA total of 24 male C57BL/6 mice were randomly divided into normal diet group (n=12) and high-fat diet group (n=12) according to the table of random number. Mice in high-fat diet group were fed with high-fat diet for 8 weeks to induce IR. Both groups were divided into normal diet control group (n=6),normal diet exercise group(n=6)and high-fat diet control group (n=6),high-fat diet exercise group (n=6),respectively.Mice in both exercise groups underwent treadmill exercise for 6 weeks. All mice did not change diet. After 14 weeks, serum visfatin concentration was evaluated by ELISA. And Western blotting was applied to detect the expression of visfatin in gastrocnemius muscle, liver and epididymal tissue.ResultsEight weeks later, compared with normal diet group, fasting insulin of high-fat diet group was elevated, and blood glucose area under curve was increased (t=6.32,17.44,allP<0.01). Fourteen weeks later, the level of serum visfatin in high-fat diet exercise group was lower than that in high-fat diet control group, but the level of serum visfatin in normal diet exercise group was higher than that in normal diet control group(F=43.59,P<0.05). Compared with high-fat diet control group, the expression of visfatin in skeletal muscle, liver and adipose tissue were reduced in high-fat diet exercise group(F=29.33-85.17, allP<0.05). On the contrary,the expression of visfatin in skeletal muscle in normal diet exercise group was higher than that in normal diet control group(F=52.79,P<0.05), but there were no differences in the expression of visfatin in liver and adipose tissue between normal diet groups(allP>0.05).ConclusionHigh-fat diet increases the level of visfatin, whereas aerobic exercise reduces the expression of visfatin and improves lipid metabolism in IR models.
Aerobic exercise;Insulin resistance;Visfatin;Lipid metabolism
10.3760/cma.j.issn.1673-4157.2016.01.003
300070 天津医科大学代谢病医院内分泌研究所,卫生部激素与发育重点实验室
于德民,Email:yudemintij@126.com
2015-06-15)
