石榴枯萎病菌拮抗放线菌对南方根结线虫的毒力
2016-11-24周银丽杨艳丽袁绍杰林丽飞胡先奇
周银丽, 杨艳丽, 袁绍杰, 林丽飞, 胡先奇*
(1. 云南农业大学农业生物多样性与病害控制教育部重点实验室, 昆明 650201;2. 红河学院云南省高校农作物优质高效栽培与安全控制重点实验室, 蒙自 661100)
石榴枯萎病菌拮抗放线菌对南方根结线虫的毒力
周银丽1,2, 杨艳丽1, 袁绍杰1, 林丽飞1, 胡先奇1*
(1. 云南农业大学农业生物多样性与病害控制教育部重点实验室, 昆明 650201;2. 红河学院云南省高校农作物优质高效栽培与安全控制重点实验室, 蒙自 661100)
从云南蒙自万亩石榴园、建水石榴园采集的土样中分离筛选得到对石榴枯萎病菌有较强拮抗活性的放线菌,其中菌株JS1对石榴枯萎病菌的室内抑菌半径为11 mm。测定了菌株JS1发酵液对南方根结线虫的抑杀活性,结果表明,50% JS1菌株发酵液处理南方根结线虫卵6 d,卵孵化率与对照相比降低了51.67%;50% JS1菌株发酵液处理南方根结线虫2龄幼虫72 h,2龄幼虫的校正死亡率增加了88.10%;菌株JS1兼有拮抗石榴枯萎病菌及抑杀南方根结线虫的活性。结合菌株JS1在鉴定培养基上的培养特征、形态学观察及16S rDNA序列分析,将JS1初步鉴定为壮观链霉菌(Streptomycesspectabilis)。
石榴枯萎病菌; 根结线虫; 生物防治; 壮观链霉菌
石榴(PunicagranatumL.),隶属于石榴科(Punicaceae),又名珍珠石榴、安石榴,若榴、山力叶、丹若、海石榴、谢榴、沃丹、天浆、金罂,是我国重要的果树之一。石榴既可观花又可食果,享有“天堂之果”的美誉[1]。云南省蒙自县被誉为“中国石榴之乡”,随着石榴栽培的集约化和树龄的增长,各种病害的发生也日益严重,尤其是石榴枯萎病的发生。石榴枯萎病是由甘薯长喙壳(CeratocystisfimbriataEllisetHalsted)引起的土传真菌病害,而该病原菌主要从甘薯、芋头、木薯及咖啡等多种木本、草本植物的根部伤口侵入,可引起甘薯黑斑病、石榴枯萎病、芋头黑腐病[2-3]等。石榴枯萎病在我国先后发生于云南省蒙自县[4]、四川省攀西地区(主要包括凉山州的会理县、西昌市和攀枝花市仁和区)[5]。 2004年,蒙自的石榴枯萎病的病株率达15.0%[6],近年来该病害呈逐年扩散加重的趋势,被当地果农称为石榴的“癌症”。已有研究表明根结线虫在石榴枯萎病发生过程中起着“打开通道”的作用[7],两者的复合侵染加重了石榴的产量损失。在蒙自十字路、红塘子、三路地、瞭望台附近、板凳桥、蒋台等地的根结线虫发生率已达30%~45%[8]。随着石榴根结线虫的发生日益加剧,石榴枯萎病的危害也日益严重。然而,石榴根结线虫的发生与危害具有隐蔽性,在生产上常被忽略。因此,寻找到防治石榴枯萎病同时能兼治根结线虫的生防菌,将对蒙自石榴产业的可持续绿色防控具有重要意义。
石榴枯萎病的防治目前主要依赖化学药剂的施用,而长期施用单一的化学药剂易使石榴枯萎病菌产生抗药性,加重果实和环境污染。利用生防微生物或其次生代谢产物进行石榴枯萎病的防治,已成为控制其进一步蔓延的有效措施,已有研究表明枯草芽胞杆菌(Bacillussubtilis)和荧光假单胞杆菌(Pseudomonasfluorescens)等土著微生物在石榴枯萎病防治中具有良好的应用前景[9]。本研究筛选了对石榴枯萎病菌及南方根结线虫(Meloidogyneincognita)卵孵化有较强抑制作用的生防放线菌,旨在为石榴枯萎病的综合防控提供参考。
1 材料与方法
1.1 材料
1.1.1 试验材料
健康石榴树根际土壤样本及枯萎病石榴树根际土壤样本,采自蒙自万亩石榴园和建水石榴园,共采集了45份样本;石榴枯萎病菌标准样品由云南农业大学植物病理实验室黄琼教授提供;南方根结线虫采自红河州蒙自市草坝镇大落就村大棚丝瓜地。
1.1.2 供试培养基
放线菌分离培养基为改良高氏1号培养基,平板对峙培养试验采用 PDA 培养基。放线菌鉴定培养基包括:蔡氏培养基,麦芽膏-酵母膏培养基,甘油-天门冬酰胺培养基,无机盐淀粉培养基,PDA培养基,葡萄糖-天门冬酰胺培养基,营养琼脂培养基,燕麦片琼脂培养基[10-11]。
1.2 试验方法
1.2.1 土壤中放线菌的分离与培养
在蒙自万亩石榴园的三角地、十字路、红塘子、三路地、瞭望台附近、板凳桥、蒋台、水井凹、马山冲和建水石榴园的周伍、小营、李伍山分别采集石榴枯萎病树根际土样(采后应将工具用75%乙醇消毒)、健康石榴树根际土样。在每点选取样株3株,从每样株周围选3处不同的位置,用锄头挖至距地表面10~20 cm处将石榴根系暴露出来,轻轻抖掉外围大块土壤,收集根系周围1 cm左右的土壤500 g,将采集的每点3株3个样本混合均匀,取500 g装入无菌自封袋,编号,将土样及时带回实验室后,进行去杂和过0.1 cm筛处理。
放线菌的分离与培养均采用改良高氏一号培养基,土壤中放线菌的分离采用平板稀释法。首先制备土壤样品稀释液,在无菌环境条件下取5 g根际土溶解于45 mL无菌水中,在28℃恒温下振荡30 min使微生物充分分散,静置10 min,即得10-1稀释液,再依次稀释为10-2、10-3、10-4、10-5系列浓度稀释液[12]。用无菌移液器吸取100 μL 浓度为10-3、10-4、10-5土壤稀释液于平板上,用涂布器将其涂均匀。置于28℃恒温下培养5 d,每个土壤稀释液浓度重复3次,将分离到的形态、颜色不同的放线菌进行画线纯化,得到单菌落,将纯化的放线菌菌株保存在改良高氏一号培养基中,置于4℃冰箱保存。
1.2.2 石榴枯萎病菌拮抗菌的筛选
拮抗菌初筛:将培养7 d的石榴枯萎病菌用灭菌水配制成浓度为103个/mL的孢子悬浮液,取100 μL涂布于PDA平板上,静置5 h。将分离得到的放线菌活化,接种到涂有石榴枯萎病病原菌的平板上,每个平板进行十字画线,在十字线距离中心3 cm位置分别接种4种放线菌,置于28℃恒温培养箱培养5 d后,观察并测定其拮抗圈的半径(r)。拮抗性强弱的分级及判定标准为:强拮抗性放线菌,r≥10 mm;中等拮抗性放线菌,7.5 mm ≤r<10 mm;弱拮抗性放线菌,r<7.5 mm;无拮抗性放线菌,r=0 mm[13]。
拮抗菌复筛:根据初筛结果,选择有明显抑菌效果的放线菌,用平板对峙法对拮抗菌进行复筛,抑菌效果的判断标准同上。
拮抗放线菌发酵液及菌丝浸出液对石榴枯萎病菌的抑菌作用:将筛选得到的放线菌在高氏一号培养基上活化后,接种于放线菌发酵培养液中,置于28℃恒温摇床150 r/min发酵培养7 d,将发酵液在10 000 r/min、4℃条件下离心20 min后得菌丝和发酵液两部分,收集所有上清发酵液,在超净工作台上用0.22 μm微孔滤膜过滤,备用[14]。菌丝溶于3倍体积的灭菌水于560 W和4 s/5 s(工作时间/间隔时间)条件下,在超声波破碎仪中冰浴破碎60 min,再以10 000 r/min离心20 min,收集上清液,在超净工作台上用0.22 μm微孔滤膜过滤,即可得菌丝提取液[15]。用孔洞法测定抑菌作用:将石榴枯萎病菌(d=5 mm)置于PDA培养基平板中心,在距中心30 mm处(d=5mm)的直径上打两个孔,每孔加入100 μL菌丝浸出物或发酵液,每种3个重复,以只接病原菌为对照,于28℃恒温箱培养,待对照长满平板,观察并测量抑菌圈半径[16]。
1.2.3 拮抗菌对南方根结线虫的影响
放线菌菌株发酵液的制备:将筛选出的放线菌在高氏一号培养基上活化后,接种于放线菌发酵培养液中,置于28℃恒温摇床150 r/min发酵培养7 d,5 000 r/min离心发酵液10 min获取上清液,取上清液用0.22 μm微孔滤膜过滤,备用。
放线菌发酵液对南方根结线虫卵孵化的影响:南方根结线虫卵悬液的制备采用改良的振荡法分离[17]。将感染南方根结线虫的丝瓜根剪成1~2 cm小段,放入200 mL 1%次氯酸钠溶液中,在豆浆机中搅拌1 min后,将悬浮液快速经过100、300和500目的套筛,上下振荡网筛,500目网筛可收集到游离的卵,快速用无菌水冲洗500目筛网上的卵以除去残留的次氯酸钠。多次冲洗振荡根段以获取更多的卵,过筛收集到离心管中,加入40%蔗糖溶液20 mL,3 500 r/min离心5min,吸取上层溶液过500目筛,用无菌水冲洗去除蔗糖液,将卵收集到三角瓶中加无菌水制成卵悬液,浓度为1 200个/mL,置于4℃冰箱备用。在24孔细胞培养板中加入50 μL浓度为1 200个/mL南方根结线虫卵悬液、放线菌菌株发酵液及灭菌水,总体积均为500 μL,调整放线菌菌株发酵液及灭菌水的加入体积,使24孔细胞培养板中放线菌菌株发酵液的浓度分别为0%、10%、20%、30%、40%、50%[18]。首先测定50%放线菌发酵液抑卵孵化作用,选择抑卵孵化率超过40%的菌株进行重复测定。每个处理3次重复,放入28℃培养箱中,分别在第2、4、6天记录2龄幼虫的孵化数,计算孵化率。孵化率(%)=2龄幼虫孵化数/总卵数×100。所得数据经软件SPSS17.0进行数据处理和方差分析。
放线菌发酵液对南方根结线虫2龄幼虫的抑杀作用测定:在解剖镜下挑取感染南方根结线虫丝瓜根组织上的卵块,经1%次氯酸钠溶液消毒后,置于无菌水中孵化2龄幼虫,定容至浓度为1 000条/mL 2龄幼虫悬液,置于4℃备用。菌株的浓度设置同上,分别向处理和对照中加入50 μL浓度为1 000条/mL南方根结线虫2龄幼虫悬液,每个浓度3次重复,放入28℃培养箱中, 分别于24、48、72 h后记录2龄幼虫死亡数,用尖细针刺激虫体,如果虫体不动就记录为死亡,计算死亡率及校正死亡率。
线虫死亡率(%)=(死亡线虫数/供试线虫数)×100;
校正死亡率(%)=(处理线虫死亡率-对照线虫死亡率)/(1-对照线虫死亡率)×100;
所得数据经软件SPSS17.0进行数据处理和方差分析。
1.2.4 生防放线菌JS1的鉴定
采用插片法培养放线菌菌株,在高氏一号培养基平板上28℃培养6~10 d,在光学显微镜下观察菌丝形态,记录菌落形态,基内菌丝、气生菌丝和孢子丝的形态及颜色,是否产生可溶性色素等。同时配制用于放线菌鉴定的培养基:蔡氏培养基、麦芽膏-酵母膏培养基、甘油-天门冬酰胺培养基、无机盐淀粉培养基、马铃薯培养基、葡萄糖-天门冬酰胺培养基、燕麦片琼脂粉培养基、营养琼脂培养基,观察菌株在其上的生长状况。
采用Promega细菌基因组提取试剂盒提取放线菌菌株JS1的DNA,用16S rDNA通用引物(27F:5′-AGA GTT TGA TCC TGG CTC AG-3′; 1492R: 5′-GGT TAC CTT GTT ACG ACT T-3′)进行PCR扩增,扩增产物切胶回收后送至华大生物技术有限公司测序,获得的序列提交GenBank数据库,用BLAST进行同源性比较,选取相似度高的不同种菌株,用MEGA5.0软件中的Neighbor-joining法构建系统发育树并进行分析。
2 结果与分析
2.1 石榴枯萎病菌拮抗放线菌的筛选
从建水石榴园和蒙自石榴园采集了45个土壤样本,分离得到菌落特征各不相同的放线菌168株,初步筛选得到38株拮抗石榴枯萎病的放线菌,进一步用平板对峙法复筛得到6株石榴枯萎病菌拮抗放线菌,6株放线菌对石榴枯萎病菌的抑菌半径在3.5~11 mm之间,其中JS1菌株对石榴枯萎病菌有较好的拮抗活性,抑菌半径为11 mm,6个菌株对石榴枯萎病菌的拮抗效果见图1,根据复筛的结果,我们选择JS1菌株进一步试验。

图1 生防菌JS1、JS2、MZ11、MZ12、MZ93、MZ1对石榴枯萎病菌的拮抗效果Fig.1 Antifungal plate of JS1, JS2,MZ11, MZ12, MZ93 and MZ1 against Ceratocystis fimbriata
采用孔洞法测定了拮抗放线菌JS1发酵液及菌丝浸出液对石榴枯萎病菌的抑菌作用,其中拮抗放线菌JS1发酵液对石榴枯萎病菌的抑菌半径为10 mm,说明放线菌JS1发酵液中含有某些抑菌物质,抑制了石榴枯萎病菌的生长;拮抗放线菌菌丝浸出液对石榴枯萎病菌没有明显的抑菌作用,说明拮抗放线菌细胞内没有抑制石榴枯萎病菌的活性物质存在。综合以上结果,拮抗放线菌JS1的抑菌成分主要存在于其发酵产物而非菌体本身。
2.2 放线菌JS1发酵液对南方根结线虫卵孵化的抑制活性
石榴枯萎病菌拮抗放线菌JS1发酵液对南方根结线虫卵孵化的抑制活性见表1。不同浓度的JS1发酵液处理南方根结线虫卵2 d,孵化率与对照相比差异不显著;不同浓度的JS1发酵液处理南方根结线虫卵4 d,浓度为30%、40%、50%的处理与对照卵孵化率相比差异显著;不同浓度的JS1发酵液处理南方根结线虫卵6 d,孵化率与对照相比都差异显著,且JS1发酵液浓度为20%、30%、40%、50%的处理与对照卵孵化率相比差异极显著。南方根结线虫卵孵化率随着JS1发酵液浓度升高而降低,50% JS1发酵液处理6 d,南方根结线虫卵孵化率与对照相比降低了51.67%,说明JS1菌株发酵液对南方根结线虫卵孵化有较好的抑制作用。
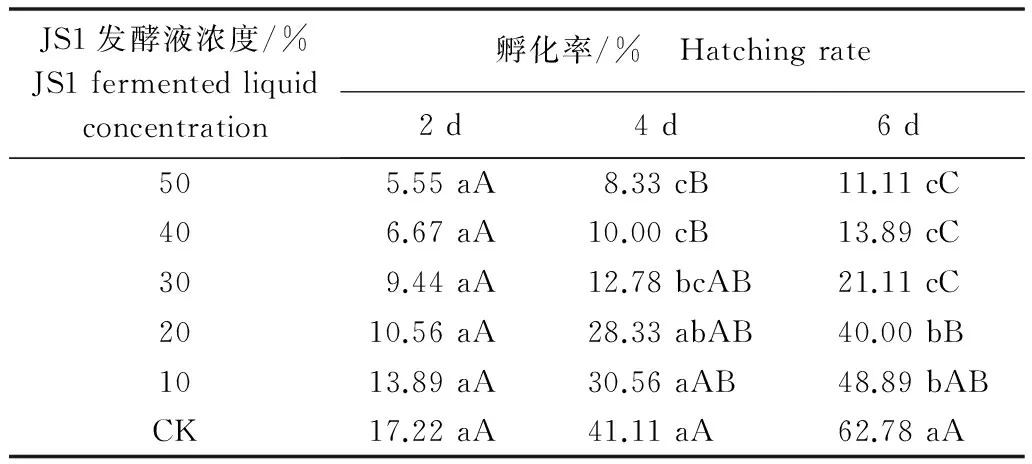
表1 JS1菌株发酵液对南方根结线虫卵孵化的抑制作用1)Table 1 Inhibition effect of JS1 fermented liquid on egg hatching of Meloidogyne incognita
1) 同列数据后不同小写字母表示数据间差异显著 (P<0.05),同列数据后不同大写字母表示数据间差异极显著 (P<0.01)。下同。
Different lowercase letters in the same column indicate significant difference at 0.05 level; different capital letters in the same column indicate significant difference at 0.01 level. The same below.
2.3 放线菌JS1发酵液对南方根结线虫2龄幼虫的抑杀活性
放线菌JS1发酵液对南方根结线虫2龄幼虫的抑杀活性见表2,不同浓度的JS1发酵液处理2龄幼虫24、48 h,发酵液浓度为30%、40%、50%的处理2龄幼虫死亡率与对照相比差异显著;不同浓度的JS1发酵液处理2龄幼虫72 h,所有处理中2龄幼虫的死亡率与对照相比差异极显著。南方根结线虫2龄幼虫的校正死亡率随着JS1发酵液浓度增大而增加,浓度50%的JS1发酵液处理72 h,南方根结线虫2龄幼虫校正死亡率与对照相比增加了88.10%,说明JS1发酵液对南方根结线虫2龄幼虫有较好的抑杀活性。

表2 JS1菌株发酵液对南方根结线虫2龄幼虫的抑杀活性Table 2 Toxicity effect of JS1 fermented liquid on Meloidogyne incognita second-stage juveniles
2.4 石榴枯萎病拮抗放线菌JS1的鉴定
石榴枯萎病拮抗放线菌JS1在高氏一号培养基上,菌丝生长好,基内菌丝秋海棠红,无隔不断裂;气生菌丝舌红,菌苔表面绒状;孢子丝直形或波曲,孢子椭圆形或长柱形,两端整齐,不产生水溶性色素。石榴枯萎病拮抗放线菌JS1在鉴定培养基上的培养特性见表3。

表3 放线菌JS1在各种鉴定培养基上的培养特性Table 3 Cultural characteristics of actinomycetes strain JS1 in different media
利用16S rDNA通用引物对菌株JS1进行PCR扩增,片段经测序后得到16SrDNA近全长序列,GenBank序列号为KT069562。Blast相似性分析表明,放线菌JS116S rDNA序列与壮观链霉菌(Streptomycesspectabilis)的同源性为100%,构建的系统发育树(图2)揭示放线菌JS1与壮观链霉菌处于同一分支。
结合形态学特征和分子特征分析,初步鉴定菌株JS1为壮观链霉菌。
3 讨论
放线菌因种类多样、分布广泛,且可产生丰富的农用抗生素和抗菌活性物质,使其在植物病害的防治及微生物农药的筛选上得到广泛应用[19]。据报道壮观链霉菌能产生大观霉素、曲张链菌素及间环丙菌素等多种抗生素[20];Chaudhary等[21]研究表明壮观链霉菌在氨基糖甙类抗生素的生产中有特殊作用,氨基糖甙类抗生素通过抑制微生物的蛋白质合成而产生快速的杀菌作用。Benallaoua等[22]从土壤中分离得到壮观链霉菌BT352,从该菌的上清液分离纯化得到一种非多烯的大环内酯结构代谢产物有抗真菌活性,研究发现这种代谢物与沙漠霉素有关。赵淑莉等[23]报道壮观链霉菌对玉米大斑病菌具有较强的拮抗作用,室内抑菌半径为15.5 mm;本研究从云南蒙自、建水石榴园采集的土样中分离筛选得到对石榴枯萎病菌有较强拮抗活性的放线菌菌株JS1,鉴定为壮观链霉菌菌株,对石榴枯萎病菌的室内抑菌半径为11 mm,与赵淑莉等的研究结果相比略低,可能是因为拮抗的病原菌不同。另Khamna等[24]的研究表明分离自七叶兰根际土壤的壮观链霉菌CMU-PA101能高效地产生生物活性代谢物质,对多种病原真菌有较好的拮抗作用;壮观链霉菌CMU-PA101对尖孢镰刀菌、齐整小核菌的拮抗作用尤其显著[25]。综合以上结果,壮观链霉菌对多种植物病原真菌有较好的拮抗作用,本研究首次发现壮观链霉菌对石榴枯萎病菌有较强的拮抗活性,其有效成分主要存在于发酵产物中,其对石榴枯萎病的田间防效有待进一步研究。

图2 以16S rDNA为基础构建的JS1系统进化树Fig.2 Phylogenetic tree of JS1 based on 16S rDNA sequences
放线菌对病原生物有很高的寄生性和拮抗作用,是控制植物病原生物的重要微生物资源,在根结线虫的生防菌株开发方面具有巨大的潜能。放线菌中的一些种类能够产生具有杀线虫特性的化合物,已报道的主要有链霉菌和诺卡氏菌[26],链霉菌产生的阿维菌素已成功应用于南方根结线虫的预防与治疗[27]。本研究中分离获得的放线菌JS1,其50%发酵液处理南方根结线虫6 d的卵孵化率与对照相比降低了51.67%,JS1菌株发酵液抑制卵孵化效果优于田阳等[28]报道的海洋放线菌M1D14发酵液,略低于陈立杰等[29]报道的放线菌C49、C25-3、C58对孢囊线虫的孵化相对抑制率。本研究中50% JS1发酵液处理南方根结线虫2龄幼虫72 h,南方根结线虫2龄幼虫校正死亡率增加了88.10%,略低于田阳等[28]报道的海洋放线菌M1D14发酵液的效果。放线菌对根结线虫的抑杀活性有较多的报道,曾庆飞等[30]分离自海泥样品中的链霉菌HA10002发酵原液对2龄幼虫的校正死亡率达100%;黄惠琴等[31]分离的放线菌菌株 HA11097发酵液稀释40倍,抗根结线虫2龄幼虫的校正死亡率为46.7%;魏华等[32]报道从红树林底泥中分离筛选的放线菌HA11090,其发酵液稀释40倍抗根结线虫校正死亡率为65.0%。
本研究分离得到的壮观链霉菌对石榴枯萎病菌有较好的室内拮抗作用,对南方根结线虫卵孵化及2龄幼虫有较强的抑杀活性。蒙自万亩石榴园中石榴根结线虫病的广泛发生加重了石榴枯萎病发生程度及病害的防控难度,寻找对石榴枯萎病及根结线虫都有防效的菌株是环保有效地控制该病害的努力方向,该菌株在石榴枯萎病及根结线虫病复合侵染的防治上有较好的开发利用潜力。
[1] Stover E D, Mercure E W.The pomegranate: a new look at the fruit of paradise [J]. HortScience, 2007, 42(5), 1088-1092.
[2] 李倩, 邓吉, 李健强. 甘薯长喙壳菌产生芳香性气体物质研究进展[J]. 植物保护, 2009, 35(4): 8-14.
[3] 刘云龙, 何永宏, 王新志. 国内一种果树新病害—石榴枯萎病[J]. 植物检疫, 2003, 17(4): 206-208.
[4] Huang Q, Zhu Y Y, Chen H R, et al. First report of pomegranate wilt caused byCeratocystisfimbriatain Yunnan, China [J]. Plant Disease, 2003, 87(9): 1150.
[5] Xu B, Zheng X H, Guo W X, et al. First report of pomegranate wilt caused byCeratocystisfimbriatain Sichuan Province[J]. Plant Disease, 2011, 95(6): 776-777.
[6] 邓吉, 陆进, 李健强, 等. 石榴枯萎病发生危害与防治初步研究[J]. 植物保护, 2006, 32(6): 97-101.
[7] 周银丽, 胡先奇, 王卫疆, 等. 根结线虫在云南石榴枯萎病发生过程中的作用初探[J]. 江苏农业科学, 2010, (1): 149-150.
[8] 周银丽, 杨伟, 余光海, 等. 中国云南省石榴根结线虫的种类初报[J]. 华中农业大学学报, 2005, 24(4): 351-354.
[9] 潘俊, 毛忠顺, 李霞, 等. 利用枯草芽孢杆菌和荧光假单胞杆菌防治石榴枯萎病的初步研究[J]. 云南农业大学学报(自然科学), 2013, 28(1): 27-32.
[10]Shirling E B, Gottlieb D.Methods for characterization ofStreptomycesspecies [J]. International Journal of Systematic and Evolutionary Microbiology, 1966, 16(3): 313-340.
[11]Williams S T, Goodfellow M, Alderson G, et al. Numerical classification ofStreptomycesand related genera [J]. Journal of General Microbiology, 1983, 129: 1743-1813.
[12]刘国生.微生物学实验技术[M]. 北京:科学技术出版社,2007:125-128.
[13]张鸿雁,薛泉宏,唐明, 等. 不同种植年限人参地土壤放线菌生态研究[J]. 西北农林科技大学学报(自然科学版),2010,38(8):151-159.
[14]杨敬辉,文平兰,陈宏州, 等. 放线菌NF0919的鉴定及生防活性[J]. 西北农业学报,2013,22(4):201-206.
[15]张武岗,冯俊涛,张锦恬, 等. 放线菌19G-317菌株发酵产物抑菌活性初步研究[J]. 西北农林科技大学学报(自然科学版),2009,37(4):158-162.
[16]李姝江,朱天辉,余琴, 等. 桑氏链霉菌突变株的抑菌活性及对杨树紫纹羽病的盆栽防效[J]. 植物保护,2015,41(5):61-68.
[17]田阳,李平,张莉,等.海洋放线菌M1D14代谢产物对几种重要植物寄生线虫的抑制作用[J].植物保护,2012,38(4):96-100.
[18]Lee Y S, Nguyen X H, Moon J H.Ovicidal activity of lactic acid produced byLysobactercapsiciYS1215 on eggs of root-knot nematode,Meloidogyneincognita[J]. Journal of Microbiology and Biotechnology, 2014, 24(11): 1510-1515.
[19]刘冰, 宋水林, 刘晓丽, 等. 生防放线菌ML27的鉴定及其抗真菌活性代谢产物的性质研究[J]. 江西农业大学学报, 2014, 36(3): 656-660.
[20]金燕华, 裘娟萍, 何景昌. 壮观链霉菌产生抗生素的多样性[J]. 中国医药工业杂志, 2006, 37(12): 849-855.
[21]Chaudhary H S, Soni B, Shrivastava A R, et al. Diversity and versatility of actinomycetes and its role in antibiotic production[J]. Journal of Applied Pharmaceutical Science, 2013, 3(8 S 1): S83-S94.
[22]Benallaoua S, Nguyen Van P, De Meo M P, et al. Recherches sur le mode d’action d’un antifongique non polyénique (désertomycine) produit par une souche deStreptomycesspectabilis[J]. Canadian Journal of Microbiology, 1990, 36(9): 609-616.
[23]赵淑莉,任飞娥,刘金亮,等.玉米大斑病生防放线菌的筛选鉴定及发酵条件优化[J].微生物学报,2012,52(10):1228-1236.
[24]Khamna S, Yokota A, Peberdy J F, et al. Antifungal activity ofStreptomycesspp. isolated from rhizosphere of Thai medicinal plants[J]. International Journal of Integrative Biology, 2009, 6(3): 143-147.
[25]Khamna S, Yokota A, Lumyong S.Actinomycetes isolated from medicinal plant rhizosphere soils: diversity and screening of antifungal compounds, indole-3-acetic acid and siderophore production [J]. World Journal of Microbiology and Biotechnology, 2009, 25(4): 649-655.
[26]罗红丽, 孙漫红, 谢建平, 等. 根结线虫放线菌及其生物防治活性研究[J]. 微生物学报,2006,46(4):598-601.
[27]朱卫刚, 胡伟群, 陈定花.阿维菌素对南方根结线虫的生物活性[J]. 现代农药,2008,7(4):38-39.
[28]田阳,李平,张莉,等.海洋放线菌M1D14代谢产物对几种重要植物寄生线虫的抑制作用[J].植物保护,2012,38(4):96-100.
[29]陈立杰,陈井生,段玉玺, 等. 防治大豆孢囊线虫的生防放线菌初步筛选[J]. 植物保护,2008,34(3):116-119.
[30]曾庆飞, 黄惠琴, 朱军, 等. 根结线虫拮抗放线菌的筛选及菌株HA10002的鉴定与活性物质分析[J]. 植物保护,2011,37(6):159-163.
[31]黄惠琴, 袁维道, 魏华, 等. 一株抗根结线虫放线菌的筛选与鉴定[J]. 生物技术通报,2013(11):175-179.
[32]魏华,刘敏,鲍时翔,等.1株抗根结线虫红树林放线菌的筛选与鉴定[J].微生物学杂志,2012,32(4):13-16.
(责任编辑:田 喆)
Nematicidal activity of actinomycete antagonistic to pomegranate wilt pathogen
Zhou Yinli1,2, Yang Yanli1, Yuan Shaojie1, Lin Lifei1, Hu Xianqi1
(1. Key Laboratory for Agro-biodiversity and Pest Management of Education Ministry of China,Yunnan Agricultural University, Kunming 650201, China; 2. Yunnan Key Laboratory for High-Quality Crops and Efficient Cultivation and Security Control, Honghe University, Mengzi 661100, China)
This study was conducted to separate actinomycetes with a strong antagonistic activity to pomegranate wilt pathogen from the soil collected from Mengzi megranate garden, Mengzi County and Jianshui Megranate Garden, Jianshui County in Yunnan Province. The indoor bacteriostatic radius of actinomycetes strains JS1 against pomegranate wilt pathogen was 11mm. Nematicidal activity of JS1 supernatant of fermented liquid toMeloidogyneincognitawas measured. The results showed that, compared with the control,M.incognitaegg hatching rate was lowered by 51.67% when treated for 6 d with 50% JS1 supernatant of the fermented liquid, and the corrected mortality rate ofM.incognitasecond-stage juveniles was increased by 88.10% if treated for 72 h with 50% JS1 supernatant of the fermented liquid. The finding suggested that strain JS1 could inhibit pomegranate wilt pathogen and was toxic toM.incognita. Strain JS1 was identified asStreptomycesspectabilisbased on its growth characteristics, morphological characteristics and analysis of 16S rDNA sequence.
pomegranate wilt pathogen; root knot nematode; biological control;Streptomycesspectabilis
2015-10-28
2015-12-10
公益性行业(农业)科研专项 (201103018-7);云南省应用基础研究项目(2013FZ125);云南省教育厅科研基金(2012Y453);红河学院硕士点植物保护一级学科建设项目
S 476
A
10.3969/j.issn.0529-1542.2016.05.009
* 通信作者 E-mail: xqhoo@126.com
