蚂蚁对不同土地利用的物种响应及生物指示
——以云南为例
2016-11-18卢志兴陈又清
卢志兴,陈又清
(中国林业科学研究院资源昆虫研究所)

蚂蚁对不同土地利用的物种响应及生物指示
——以云南为例
卢志兴,陈又清*
(中国林业科学研究院资源昆虫研究所)
为了揭示不同土地利用状态下蚂蚁群落在物种层面的变化,采用陷阱法调查了云南省绿春县天然次生林、桉树林、紫胶林、紫胶-玉米混农林、橡胶林、旱地和农田7种类型样地地表蚂蚁群落。共采集地表蚂蚁16704头,隶属于8亚科44属106种。不同类型样地地表蚂蚁群落结构有差异(ANOSIM Global R=0.786,P=0.01),桉树林、紫胶林和紫胶玉米混农林地表蚂蚁群落结构与天然次生林相似,与橡胶林和农田不相似。不同样地具有各自的特有种,旱地和农田中各为1种,其余样地有5-8种。在多个样地中均有出现的物种在不同类型样地中的多度变化明显。对群落结构相似性贡献率≥5%的特征物种分析显示,天然次生林中的特征物种明显与旱地和农田不同。天然次生林中的指示物种为毛发铺道蚁Tetramoriumciliatum,桉树林中为亮褐举腹蚁Crematogastercontemta,紫胶林中为飘细长蚁Tetraponeraallaborans,旱地中为伊大头蚁Pheidoleyeensis。不同特征物种及指示物种的生物学特性与不同样地特征相关联。地表蚂蚁通过物种组成及多度变化响应土地利用变化,蚂蚁特征物种可以作为指示物种的补充。
土地利用;蚂蚁;群落结构;特征物种;指示物种
土地利用变化一般被认为是一种区域性的环境问题,但是其重要性越来越受到全球关注。这种由人类主导的实践活动正导致热带雨林消失,强化农林复合系统及农田的产出,已经改变了地球上很大比例的地表特征(Foleyetal.,2005)。土地利用变化是一种影响生物多样性的重要的全球性压力(Salaetal.,2000;UNEP,2002;Zebischetal.,2004)。大部分的研究只是关注最重要的压力,即原始森林清除的影响,而极少关注农业系统土地利用强度对生物多样性的影响(Salaetal.,2000)。然而,随着农业用地利用强度的增加,已经导致日益增长的生物多样性压力,而且这种趋势可能增强(Tilmanetal.,2001),在今后的几十年里,这种对生物多样性的压力可能演变成一种最重要的形式(Petitetal.,2001)。
生物多样性受到影响后最终以物种的变化形式体现,目前研究多从物种多样性、物种组成、多度、结构相似性等层面研究土地利用变化对群落的影响(Gómezetal.,2003;Dauberetal.,2006;Yeoetal.,2011;Rizalietal.,2013)。群落受到影响后,群落中的哪些物种受到影响以及如何受影响,才是理解群落响应栖境变化的机制的基础(Lavoreletal.,2011)。
蚂蚁在陆地生态系统中几乎均有分布,具有较高的生物多样性和重要的生态系统功能(Hölldobler and Wilson,1990),其多度、物种丰富度及群落结构变化被认为能够较好指示生态系统运行状况(Philpottetal.,2008;Andersenetal.,2012;Lukeetal.,2014)。已有研究显示,土壤和植被是影响蚂蚁群落的主要因素,其次是干扰导致大部分蚂蚁多度降低,改变蚂蚁群落结构(Hoffmann and Anderson,2003;Philpottetal.,2008;Hoffmann,2010)。在不同利用强度下,蚂蚁物种组成、群落结构及动态能够反映出干扰强度变化(Hoffmann,2000)。在低利用强度下,少数物种竞争优势地位,排斥其它物种,而在高利用强度下,仅少量压力耐受物种和特化物种能够存活(Bestelmeyer and Wiens,1996;Andersonetal.,2004)。在同等干扰条件下,对环境变化敏感的物种受到环境变化的影响最大,通过研究这些物种的变化,可以快速获知生态系统状态。
本研究通过调查不同土地类型地表蚂蚁群落,比较不同样地中物种有无及其多度变化,分析不同样地的特征蚂蚁及指示物种,在物种层面探讨1)土地利用差异导致的群落结构变化中哪些物种在变化?2)物种如何变化?以期为今后在山地土地利用变化中有针对性地保护蚂蚁多样性提供理论指导。
1 材料与方法
1.1 研究地概况
在云南省绿春县大兴镇、牛孔乡和大黑山乡3个地点选取天然次生林(NF)、桉树林(EP)、紫胶林(LP)、紫胶-玉米混农林(MP)、橡胶林(RP)、旱地(DL)和农田(FL) 7种类型样地调查蚂蚁群落,每种类型各选取两块面积大于1 hm2的样地作为重复样地,间距1 km以上。7种类型样地均处于阳坡,坡度基本一致,土壤条件差异不大,同一类型两块重复样地的植被、郁闭度、枯落物等情况基本一致。天然次生林位于牛孔乡海拔1150-1235 m地段,植被保护较好,主要乔木有麻栎Quercusacutissima、栓皮栎Quercusvariabilis、云南松Pinusyunnanensis等,林下地表覆盖大量枯落物,郁闭度65%以上;紫胶林位于牛孔乡海拔1000-1065 m地段,主要乔木为紫胶虫寄主植物南岭黄檀Dalbergiabalansae,林下草本层发达,样地内长期放养云南紫胶虫Kerriayunnanensis,地表有少量枯落物,郁闭度约50%;紫胶-玉米混农林位于牛孔乡海拔980-1000 m地段,主要乔木为南岭黄檀Dalbergiabalansae,密度为紫胶林1/3,林下在雨季种植玉米Zeamays,旱季则撂荒,郁闭度约40%;旱地位于牛孔乡海拔900-1000 m地段,雨季种植玉米,旱季则撂荒,撂荒期草本层发达;农田位于牛孔乡海拔950-1030 m地段,长期从事水稻生产,采样前种植过水稻,土壤板结,地表有少量草本植物;桉树林位于大兴镇海拔1400-1420 m地段,以6年生桉树eucalyptusgrandis为主,灌草层发达,地表有枯落物,郁闭度约50%;橡胶林为30年生橡胶Heveabrasiliensis树林,地面有固定的收胶路线,橡胶树行间有草本和灌木植物生长,地表有枯落物,郁闭度75%以上。
1.2 调查方法
分别于2012年10月和2013年4月进行了7种类型样地两次蚂蚁群落调查。使用直径6 cm,高8 cm的塑料杯作为陷阱调查蚂蚁群落,每个样地设置5×3网格状的15个陷阱,间距10 m,每个陷阱内倒入50 mL 50%乙二醇作为陷阱液,放置48 h后收集蚂蚁置于含有75%酒精的离心管中带回实验室。根据相关资料将蚂蚁鉴定到种,不能鉴定到种的,按形态种对待(吴坚和王常禄,2000;徐正会,2002)。
1.3 数据分析
为了避免陷阱处于蚂蚁活动路径或蚁巢旁时出现单个陷阱个体数偏大的情况,对蚂蚁多度数据进行6级评分转换,1级1头,2级2-5头,3级6-10头,4级11-20头,5级21-50头,6级大于50头(Hoffmann and Andersen,2003;卢志兴等,2013),利用统计软件PRIMER v7中的层次聚类方法(Hierarchical Cluster analysis)比较各样地蚂蚁群落结构相似性(基于Bray-Curtis similarity),并使用相似性分析(ANOSIM,analysis of similarities)检验7种类型样地蚂蚁群落结构相似性差异显著性,以平方根转换后的多度数据绘制各物种在不同了些样地中的阴影图(Shade plot),比较不同样地蚂蚁群落中的物种组成及多度变化(Clarke and Gorley,2006)。同时,使用PRIMER v7中的相似性百分比(SIMPER,Similarity Percentages)方法分析不同类型样地中的特征蚂蚁,选取对群落结构相似性贡献率≥5%的蚂蚁作为主要特征种类(Clarke and Warwick,2001)。利用统计软件R语言中的Labdsv软件包计算不同类型样地地表蚂蚁指示物种(R Team,2012),以IndVal值≥0.7作为指示物种的判定标准(Nakamuraetal.,2007)。
2 结果与分析
2.1 不同类型样地地表蚂蚁群落结构及物种组成变化
共采集地表蚂蚁16704头,隶属于8亚科44属106种。其中天然次生林37属86种;桉树林中32属76种;紫胶林36属79种;橡胶林34属67种;紫胶-玉米混农林34属68种;旱地22属41种;农田11属22种。
不同类型样地地表蚂蚁群落结构有差异(ANOSIM Global R=0.786,P=0.01),桉树林地表蚂蚁群落结构与天然次生林相似,紫胶林与紫胶-玉米混农林相似,旱地与紫胶林及紫胶-玉米混农林相似。上述5种类型样地地表蚂蚁群落结构与橡胶林和农田不相似,橡胶林地表蚂蚁群落结构与农田不相似(图1)。7种类型样地均有各自的特有物种(见附表)。弯刺角腹蚁Recurvidrisrecurvispinosa等8种蚂蚁仅出现在桉树林中,多毛厚结猛蚁Pachycondylapilosior等7种蚂蚁仅出现在天然次生林中,光胫多刺蚁Polyrhachistibialis等5种蚂蚁仅出现在紫胶林中,克氏锯猛蚁Prionopeltakraepelini等7种蚂蚁仅出现在紫胶-玉米混农林中,大隐猛蚁Cryptoponegigas仅出现在旱地中,云南细蚁Leptanillasp.1等5种仅出现在橡胶林中,费氏臭蚁Dolichoderusfeae仅出现在农田中。在多个样地中均有出现的物种中,不同样地间该物种的多度有差异。其中,毛发铺道蚁Tetramoriumciliatum和舒尔盘腹蚁Aphaenogasterschurri在天然次生林中的多度较高,卡泼林大头蚁Pheidolecapellini、立毛举腹蚁Crematogasterferrarii和平和弓背蚁Camponotusmitis在紫胶林中的多度较高,女娲角腹蚁Recurvidrisnuwa、棒刺大头蚁Pheidolespathifera、东方小家蚁Monomoriumorientale和中华小家蚁Monomoriumchinensis在紫胶-玉米混农林中的多度较高,伊大头蚁Pheidoleyeensis在旱地中的多度较高,皮氏大头蚁Pheidolepieli和沃森大头蚁Pheidolewatsoni在橡胶林中的多度较高,罗氏心结蚁Cardiocondylawroughtonii和扁平虹臭蚁Iridomyrmexanceps在农田中的多度较高。(见图1和附表)
2.2 不同类型样地的特征物种
进一步分析不同类型样地中对群落结构相似性贡献累积超过70%的物种,结果显示地表蚂蚁群落结构在天然次生林中变化较大(该类型样地的群落平均相似性为39.87%),在桉树林和紫胶-玉米混农林中变化相对较小(该类型样地的群落平均相似性分别为65.73%和64.66%),紫胶林、橡胶林、旱地和农田则超过50%。在多个类型样地中出现的特征蚂蚁物种为横纹齿猛蚁Odontoponeratransversa(5个类型样地均出现)、棒刺大头蚁(5个类型样地均出现)和来氏大头蚁Pheidolelighti(4个类型样地均出现),其中,横纹齿猛蚁在天然次生林中具有最高的群落相似性贡献比例,在紫胶林中最低(见表1);棒刺大头蚁在紫胶林和旱地中具有较高的群落相似性贡献比例(见表1);来氏大头蚁在桉树林中具有较高的群落相似性贡献比例。
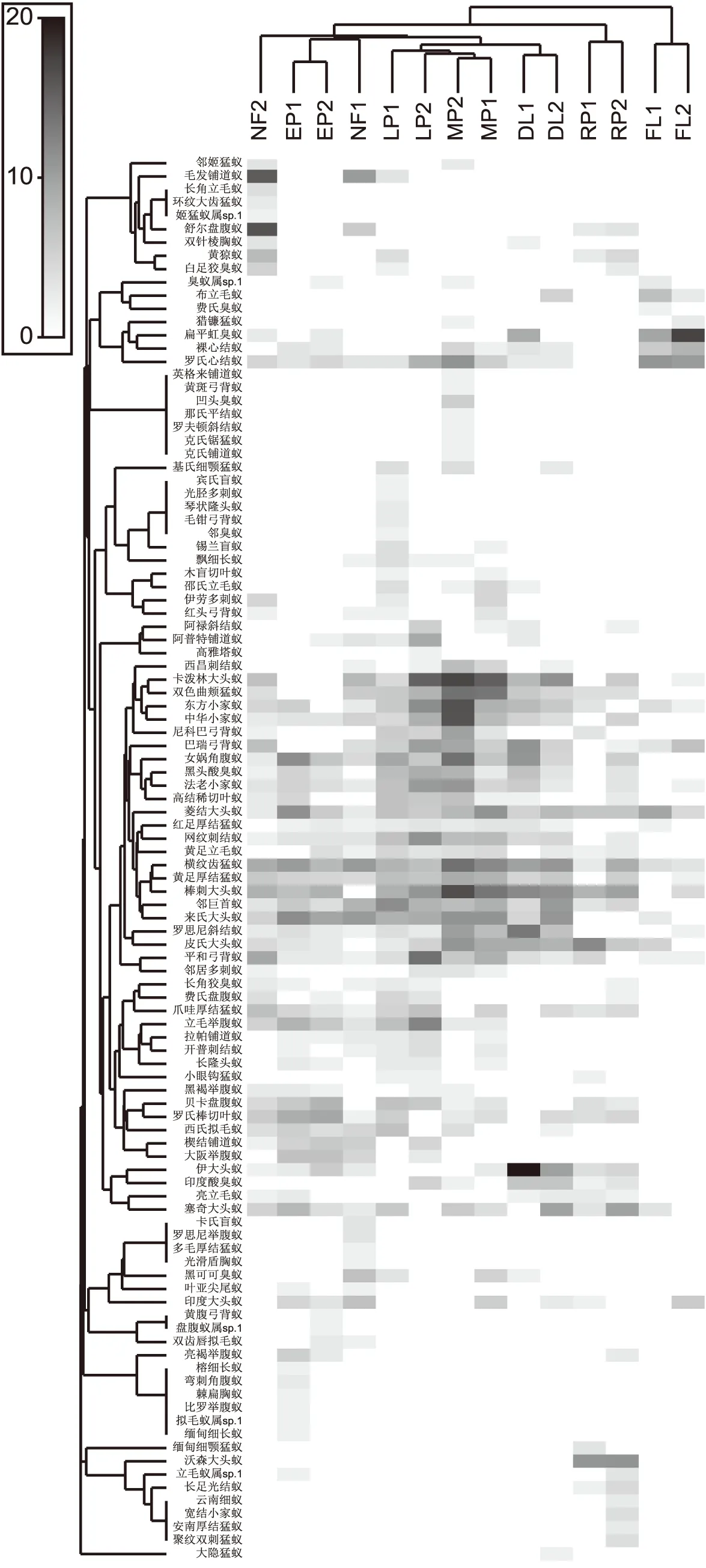
图1 不同样地地表蚂蚁群落结构层次聚类阴影图Fig.1 Shade plot of ant communities among different plots based on hierarchical cluster analysis

表1 不同样地地表蚂蚁群落相似性平均贡献率
注:平均相似性是同一类型样地重复样地的相似性。NF,天然次生林;EP,桉树林;LP,紫胶林;RP,橡胶林;MP,紫胶玉米混农林;DL,旱地;FL,农田。附表同。Note: Average similarity means similarity of repeated sample in same site. NF, Secondary natural forest; EP, Eucalyptus plantation; LP, Lac insect plantation; RP, Rubber plantation; MP, Lac-corn agroforest; DL, Dry land; FL,Farmland. Same in the appendix。
不同类型样地特征物种存在差异,其中天然次生林中的特征蚂蚁为毛发铺道蚁和舒尔盘腹蚁;桉树林中为大阪举腹蚁Camponotusparius和罗氏棒切叶蚁Rhoptromyrmexwroughtonii;紫胶林中为邻巨首蚁Pheidologetonaffinis和黑头酸臭蚁Tapinomamelanocephalum;紫胶混农林中为双色曲颊猛蚁Gnamptogenysbicolor;橡胶林中为菱结大头蚁Pheidolenoda和沃森大头蚁;旱地中为伊大头蚁、印度酸臭蚁Tapinomaindicum和罗思尼斜结蚁Plagiolepisrothneyi;农田中为罗氏心结蚁、裸心结蚁Cardiocondylanuda和扁平虹臭蚁。
2.3 不同类型样地地表蚂蚁指示物种
不同类型样地地表蚂蚁指示物种明显不同(表2)。旱地中为喜欢开放栖境的伊大头蚁;紫胶林中指示物种为飘细长蚁Tetraponeraallaborans;天然次生林中指示物种为毛发铺道蚁;桉树林中指示物种为亮褐举腹蚁Crematogastercontemta。橡胶林、紫胶-玉米混农林和农田中无指示物种。

表2 不同土地类型地表蚂蚁指示值分析
注:IndValij=Aij×Bij,Aij表示物种i在样地j中的特异性,Bij表示物种i在样地j中的保真度。Note:Aijis the proportion of sites of typejwith speciesi,Bijis the proportion of the number of abundance of speciesithat are in ajtype of site.
3 结论与讨论
物种是生态系统功能实现的载体,基于物种组成的群落结构分析可以较好揭示出生态系统运行状态(Lavoreletal.,2011),土地利用变化直接改变了蚂蚁群落结构。整体上看,天然次生林、人工林、混农林及农耕系统中地表蚂蚁群落结构不相似(图1)。首先,不同类型样地中特有物种数量不一致,旱地和农田中各有1个特有物种,其余类型样地中有5至8种特有物种(表1)。不同土地利用的生境与特有蚂蚁物种的生态学特性密切相关,天然次生林中的环纹大齿猛蚁、卡氏盲蚁和光滑盾胸蚁特有种被认为是分布在植被相对较好、有枯落物层的栖境中(徐正会,2002);紫胶林和紫胶-玉米混农林中的光胫多刺蚁、黄斑弓背蚁和凹头臭蚁喜欢在植物上活动且喜食蜜露资源(卢志兴等,2013);农田中的费氏臭蚁多分布在开放有干扰的栖境中(徐正会,2002)。其次,多度发生变化的物种也明显响应了土地利用变化导致的栖境变化。毛发铺道蚁和舒尔盘腹蚁在植被条件较好的天然次生林中的多度较高,在其余样地中多度较低或没有,同时,毛发铺道蚁也是天然次生林的指示物种,表明这两种蚂蚁需要依赖较好质量栖境生存,与前人研究一致(Chenetal.,2011);伊大头蚁、扁平虹臭蚁、裸心结蚁和罗氏心结蚁在旱地和农田中的多度较高,伊大头蚁也是旱地的指示物种,表明这些蚂蚁趋向于开放有干扰的栖境,与国外研究一致(Hoffmann,2003)。第三,蚂蚁群落结构相似性反映了不同土地利用下生境条件的相似性。天然次生林和桉树林特征物种毛发铺道蚁和舒尔盘腹蚁为依赖具枯落物层复杂栖境的物种(张智英等,2005;Chenetal.,2011),大阪举腹蚁和罗氏棒切叶蚁则需要依赖植物空间结构较为复杂的栖境生存(陈友等,2013),紫胶林的黑头酸臭蚁喜食蜜露(Chenetal.,2011,卢志兴等,2013),旱地中的伊大头蚁和农田中的扁平虹臭蚁则喜欢有干扰开放的栖境(Hoffmann,2003)。
土地利用转变导致蚂蚁群落多样性降低,物种组成及群落结构随着干扰及利用强度发生改变(Hoffmann,2010;de Castro Solaretal.,2016),一方面,这种差异是由于样地土壤、植被组成及结构变化造成,另一方面则是由于不同土地利用强度差异造成(Hoffmann and Andersen,2003;Armbrechtetal.,2006)。本研究结果显示,不同类型样地蚂蚁群落组成及结构差异明显,在林分条件好、干扰少的天然次生林或部分人工林中(如桉树林),蚂蚁物种组成及群落结构不同于利用强度高、干扰多的农业系统,与前人研究一致。人工林由于树种单一被认为不利于多样性保护,实际上,在保护相对较好,干扰少的桉树林、具有提供蜜露食物资源的紫胶林以及复合型的紫胶-玉米混农林系统中,蚂蚁物种组成及群落结构与天然次生林具有一定的相似性,这对西南脆弱山地生态系统中的生物多样性保护具有积极作用。
已有研究多通过分析蚂蚁指示物种,以这些物种的生物学习性和生态学特点来指示栖境变化或栖境质量情况(Hoffmann and Andersen,2003;Hoffmann,2010;Menkeetal.,2015)。由于很多蚂蚁的生态学研究仍然是空白,可能会缺乏对应指示物种的生态学资料;同时,对一些栖境数据的分析并不能获得指示物种,使得指示物种方法在运用中受到一定的限制(Nakamuraetal.,2007;Hoffmann,2010)。基于SIMPER的特征物种分析说明,物种在某类型样地中对群落相似性贡献率越高,则在该种栖境中的存在越稳定,与该栖境的关联程度较可靠(Nakamuraetal.,2007;Delabieetal.,2009)。本文通过对云南蚂蚁群落结构在不同土地利用方式下的变化分析,筛选出在蚂蚁群落变化中起关键作用的物种,也能较好指示栖境变化和栖境质量情况,可作为指示物种的补充加以利用。
致谢:感谢西南林业大学徐正会教授帮助核实标本。
References)
Armbrecht I, Perfecto I, Silverman E. Limitation of nesting resources for ants in Colombian forests and coffee plantations[J].EcologicalEntomology, 2006, 31(5): 403-410.
Andersen AN, Fisher A, Hoffmann BD,etal. Use of terrestrial invertebrates for biodiversity monitoring in Australian rangelands, with particular reference to ants[J].AustralEcology, 2004, 29(1): 87-92.
Andersen AN, Woinarski JCZ, Parr CL. Savanna burning for biodiversity: Fire management for faunal conservation in Australian tropical savannas[J].AustralEcology, 2012, 37(6): 658-667.
Bestelmeyer BT, Wiens JA. The effects of land use on the structure of ground-foraging ant communities in the Argentine Chaco[J].ecologicalApplications, 1996: 1225-1240.
Chen YQ, Li Q, Chen YL,etal. Ant diversity and bio-indicators in land management of lac insect agroecosystem in Southwestern China[J].BiodiversityandConservation, 2011, 20(13): 3017-3038.
Chen Y, Luo CW, Li HW,etal. Investigation of ant species and distribution on Wuliang mountain[J].JournalofHenanAgriculturalSciences, 2013, 42(5): 118-122. [陈友, 罗长维, 李宏伟, 等. 无量山蚂蚁种类及其分布[J].河南农业科学, 2013, 42(5): 118-122]
Clarke KR, Gorley RN. PRIMER Version 6: User Manual/Tutorial[M]. Plymouth:PRIMER-E, 2006.
Clarke KR,Warwick RM. An Approach to Statistical Analysis and Interpretation.Change in Marine Communities[M]. Plymouth:PRIMER-E, 2001.
Dauber J, Bengtsson JAN, Lenoir L. Evaluating effects of habitat loss and land-use continuity on ant species richness in seminatural grassland remnants[J].ConservationBiology, 2006, 20(4): 1150-1160.
Delabie JHC, Céréghino R, Groc S,etal.Ants as biological indicators of Wayana Amerindian land use in French Guiana[J].Comptes.Rendus.Biologies, 2009, 332(7): 673-684.
Foley JA, DeFries R, Asner GP,etal.Global consequences of land use[J].Science,2005, 309(5734): 570-574.
Gómez C, Casellas D, Oliveras J,etal. Structure of ground-foraging ant assemblages in relation to land-use change in the northwestern Mediterranean region[J].BiodiversityandConservation, 2003, 12(10): 2135-2146.
Hoffmann BD, Andersen AN. Responses of ants to disturbance in Australia, with particular reference to functional groups[J].AustralEcology, 2003, 28(4): 444-464.
Hoffmann BD. Responses of ant communities to experimental fire regimes on rangelands in the Victoria River District of the Northern Territory[J].AustralEcology, 2003, 28(2):182-195.
Hoffmann BD. Using ants for rangeland monitoring: Global patterns in the responses of ant communities to grazing[J].EcologicalIndicators, 2010, 10(2):105-111.
Hölldobler B, Wilson EO. The Ants[M]. Harvard University Press, 1990.
Lavorel S, Grigulis K, Lamarque P,etal. Using plant functional traits to understand the landscape distribution of multiple ecosystem services[J].JournalofEcology,2011, 99:135-147.
Lu ZX, Chen YQ, Zhang W,etal. Effects of facultative mutualism between ants and lac insects on the diversity of ant communities[J].BiodiversityScience, 2013, 21(3): 343-351. [卢志兴, 陈又清, 张威, 等. 蚂蚁-紫胶虫兼性互利关系对蚂蚁群落多样性的影响[J]. 生物多样性, 2013, 21(3): 343-351]
Luke SH, Fayle TM, Eggleton P,etal. Functional structure of ant and termite assemblages in old growth forest, logged forest and oil palm plantation in Malaysian Borneo[J].BiodiversityandConservation, 2014, 23(11): 2817-2832.
Menke SB, Gaulke E, Hamel A,etal. The effects of restoration age and prescribed burns on grassland ant community structure[J].EnvironmentalEntomology, 2015,44(5):1336-1347.
Nakamura A, Catterall CP, House APN,etal. The use of ants and other soil and litter arthropods as bio-indicators of the impacts of rainforest clearing and subsequent land use[J].JournalofInsectConservation, 2007, 11(2): 177-186.
Petit S, Firbank L, Wyatt B,etal. MIRABEL: Models for integrated review and assessment of biodiversity in European landscapes[J].AMBIO:AJournaloftheHumanEnvironment, 2001, 30(2): 81-88.
Philpott SM, Arendt WJ, Armbrecht I,etal.Biodiversity loss in Latin American coffee landscapes: Review of the evidence on ants, birds, and trees[J].ConservationBiology, 2008, 22(5): 1093-1105.
Rizali A, Clough Y, Buchori D,etal. Long-term change of ant community structure in cacao agroforestry landscapes in Indonesia[J].InsectConservationandDiversity, 2013, 6(3): 328-338.
Sala OE, Chapin FS, Armesto JJ,etal.Global biodiversity scenarios for the year 2100[J].Science, 2000,287(5459):1770-1774.
Team RC. R:A Language and Environment for Statistical Computing[M]. Vienna: R Foundation for Statistical Computing,2012.
Tilman D, Fargione J, Wolff B,etal. Forecasting agriculturally driven global environmental change[J].Science, 2001, 292(5515): 281-284.
United Nations Environment Programme. Global Environmental Outlook: 3. Past, Present and Future Perspectives[M]. London: Earthscan Publications, 2002.
Wu J, Wang CL. The Ants of China[M].Beijing:China Forestry Publishing House, 1995. [吴坚, 王常禄. 中国蚂蚁[M]. 北京:中国林业出版社, 1995]
Xu ZH. A Study on the Biodiversity of Formicidae Ants of Xishuangbanna Nature Reserve[M]. Kunming: Yunnan Science and Technology Press, 2002.[徐正会. 西双版纳自然保护区蚁科昆虫生物多样性研究[M]. 昆明: 云南科技出版社, 2002]
Yeo K, Konate S, Tiho S,etal. Impacts of land use types on ant communities in a tropical forest margin (Oumé-Cte d’Ivoire)[J].AfricanJournalofAgriculturalResearch, 2011, 6(2): 260-274.
Zebisch M, Wechsung F, Kenneweg H. Landscape response functions for biodiversity-assessing the impact of land-use changes at the county level[J].LandscapeandUrbanPlanning, 2004, 67(1): 157-172.
Zhang ZY, Li YH, Chai DM,etal. Ant biodiversity in different habitats in Shilin Park, Yunnan Province[J].BiodiversityScience, 2005, 13(4): 357-362. [张智英, 李玉辉, 柴冬梅, 等. 云南石林公园不同生境蚂蚁多样性研究[J]. 生物多样性, 2005, 13(4): 357-362]

附表 不同样地蚂蚁名录及多度
续上表

物种名SpeciesNFEPLPRPMPDLFL 聚纹双刺猛蚁Diacammarugosum4 横纹齿猛蚁Odontoponeratransversa67565426167878盲蚁亚科Aenictinae 锡兰盲蚁Aenictusceylonicus41 宾氏盲蚁Aenictusbinghami1 卡氏盲蚁Aenictuscamposi7细蚁亚科Leptanillinae 云南细蚁Leptanillasp13 伪切叶蚁亚科Pseudomyrmecinae 飘细长蚁Tetraponeraallaborans151 榕细长蚁Tetraponeramicrocarpa1 缅甸细长蚁Tetraponerabirmana1切叶蚁亚科Myrmicinae 罗思尼举腹蚁Crematogasterrothneyi7 黑褐举腹蚁Crematogasterrogenhoferi2411 立毛举腹蚁Crematogasterferrarii16521656 亮褐举腹蚁Crematogastercontemta142 比罗举腹蚁Crematogasterbiroi1 大阪举腹蚁Crematogasterosakensis9332 琴状隆头蚁Strumigenyslyroessa2 长隆头蚁Strumigenysexilirhina461 木盲切叶蚁Carebaralignata11 高雅塔蚁Pyramicaelegantula1 高结稀切叶蚁Oligomyrmexaltinodus361202141 邻巨首蚁Pheidologetonaffinis65441635481115 光滑盾胸蚁Meranopluslaeviventris1 双针棱胸蚁Pristomyrmexpungens41 女娲角腹蚁Recurvidrisnuwa8117421015666 弯刺角腹蚁Recurvidrisrecurvispinosa4 法老小家蚁Monomoriumpharaonis524171782241 东方小家蚁Monomoriumorientale16312176438456 宽结小家蚁Monomoriumlatinode19 中华小家蚁Monomoriumchinensis1261101053318 棘扁胸蚁Vollenhoviaacanthina1 罗氏棒切叶蚁Rhoptromyrmexwroughtonii1065206522726 英格来铺道蚁Tetramoriuminglebyi1
续上表

物种名SpeciesNFEPLPRPMPDLFL 拉帕铺道蚁Tetramoriumlaparum2271 毛发铺道蚁Tetramoriumciliatum3263 阿普特铺道蚁Tetramoriumaptum31362 克氏铺道蚁Tetramoriumkraepelini1 楔结铺道蚁Tetramoriumcuneinode102111 棒刺大头蚁Pheidolespathifera43661116546618813 来氏大头蚁Pheidolelighti120142372158542 卡泼林大头蚁Pheidolecapellini121216208671651 伊大头蚁Pheidoleyeensis3412352858 菱结大头蚁Pheidolenoda712039371233573 印度大头蚁Pheidoleindica3221112226 沃森大头蚁Pheidolewatsoni142 塞奇大头蚁Pheidolesagei14372322216377 皮氏大头蚁Pheidolepieli247121401827813 罗氏心结蚁Cardiocondylawroughtonii121138935150 裸心结蚁Cardiocondylanuda79842 贝卡盘腹蚁Aphaenogasterbeccarii5303494 费氏盘腹蚁Aphaenogasterfeae41121 舒尔盘腹蚁Aphaenogasterschurri3865 盘腹蚁属Aphaenogastersp11臭蚁亚科Dolichoderinae 白足狡臭蚁Technomyrmexalbipes5213 长角狡臭蚁Technomyrmexantennus3131 黑头酸臭蚁Tapinomamelanocephalum11117342547 印度酸臭蚁Tapinomaindicum179132 费氏臭蚁Dolichoderusfeae1 黑可可臭蚁Dolichoderusthoracicus42671 凹头臭蚁Dolichoderusincisus11 邻臭蚁Dolichoderusaffinis1 臭蚁属Dolichoderussp1112 扁平虹臭蚁Iridomyrmexanceps34134312蚁亚科Formicinae 叶亚尖尾蚁Acropygayaeyamensis11 开普刺结蚁Lepisiotacapensis1193 西昌刺结蚁Lepisiotaxichangensis11551 网纹刺结蚁Lepisiotareticulate477423713
续上表
The species-level responses of ant communities to land use and its bio-indicators: A case study in Yunnan Province
LU Zhi-Xing, CHEN You-Qing*
(Research Institute of Resources Insects, Chinese Academy of Forestry, Kunming 650224, China)
In order to reveal the species-level change of ant communities in land use changing process, investigations were conducted on ground-dwelling ant communities of secondary natural forest, eucalyptus plantation, lac insect plantation, lac-corn agroforest, rubber plantation, dry land and farmland in Lüchun County,Yunnan Province. 16704 ant individuals were collected, representing 106 species 44 genera in 8 subfamilies of Formicidae. Community structures of ground-dwelling ants had significantly differences among different sites (ANOSIM Global R=0.786,P=0.01). Ant community structures of eucalyptus plantation, lac insect plantation and lac-corn agroforest were similar with secondary natural forest, and these sites were dissimilar with rubber plantation and farmland. Different sites had their endemic species. There was only one endemic species in dry land and farmland, and other sites had five to eight endemic species, respectively. Abundance of species appeared in most sites had a significantly change within different sites. Typical species which had contributions over 5% to community similarities showed typical species belong to secondary natural forest were different form dry land and farmland.Tetramoriumciliatumis the indicator of secondary natural forest.Crematogastercontemtais the indicator of eucalyptus plantation.Tetraponeraallaboransis the indicator of lac insect plantation.Pheidoleyeensisis the indicator of dry land. Biology and ecology of typical species and indicators were associated with each habitat. Ground-dwelling ants had responses to land use change by species composition and abundance change. Typical species can use as a complement to the indicator species application.
Land-use; ants; community structure; typical species; indicators

物种名SpeciesNFEPLPRPMPDLFL 阿禄斜结蚁Plagiolepisalluaudi10112 罗思尼斜结蚁Plagiolepisrothneyi51061512351 罗夫顿斜结蚁PlagiolepisWroughtoni1 长足光结蚁Anoplolepisgracilipes6 西氏拟毛蚁Pseudolasiussilvestrii14141561 双齿唇拟毛蚁Pseudolasiusbidenticlypeus12 拟毛蚁属sp1Pseudolasiussp11 邵氏立毛蚁Paratrechinasauteri5101 长角立毛蚁Paratrechinalongicornis5 亮立毛蚁Paratrechinavividula1244 黄足立毛蚁Paratrechinaflavipes1493123 布立毛蚁Paratrechinabourbonica1119 立毛蚁属sp1Paratrechinasp113 那氏平结蚁Prenolepisnaraojii1 黄猄蚁Oecophyllasmaragdina4678 光胫多刺蚁Polyrhachistibialis1 邻居多刺蚁Polyrhachisproxima136 伊劳多刺蚁Polyrhachisillaudata736 毛钳弓背蚁Camponotuslasiselene2 巴瑞弓背蚁Camponotusparius33591759916 尼科巴弓背蚁Camponotusnicobarensis1634201 平和弓背蚁Camponotusmitis56427574862 黄斑弓背蚁Camponotusalbosparsus1 黄腹弓背蚁Camponotushelvus1 红头弓背蚁Camponotussingularis213
国家自然科学基金项目(31270561,31470493)
卢志兴,男,1986年生,云南华宁人,博士,研究方向为昆虫生态学,E-mail:endeavou@163.com
*通讯作者Author for correspondence,E-mail: cyqcaf@126.com
Received:2015-11-25;接受日期Accepted:2016-06-03
Q968.1
A
1674-0858(2016)05-0950-11
