基于Ecopath模型估算莱州湾朱旺人工鱼礁区日本蟳、脉红螺捕捞策略和刺参增殖生态容量*
2016-11-17杨超杰吴忠鑫刘鸿雁张沛东李文涛曾晓起张秀梅
杨超杰, 吴忠鑫, 刘鸿雁, 张沛东, 李文涛, 曾晓起, 张秀梅
(1.中国海洋大学水产学院,山东 青岛266003; 2.青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛266235)
基于Ecopath模型估算莱州湾朱旺人工鱼礁区日本蟳、脉红螺捕捞策略和刺参增殖生态容量*
杨超杰1, 吴忠鑫1, 刘鸿雁1, 张沛东1, 李文涛1, 曾晓起1, 张秀梅**
(1.中国海洋大学水产学院,山东 青岛266003; 2.青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛266235)
于2010—2011年在莱州湾朱旺人工鱼礁区(建礁3年)采用地笼网进行周年渔业资源调查,根据调查结果,使用Ecopath with Ecosim 6.4.3软件,构建了莱州湾朱旺人工鱼礁区生态系统食物网模型,提出了人工鱼礁区中日本蟳(Charybdisjaponica)、脉红螺(Rapanavenosa)和刺参(Apostichopusjaponucus)的管理策略。研究表明:功能组主要占据3个营养级;系统总流量为3 390.131 t·km-2·a-1,总消耗量为1 839.502 t·km-2·a-1,总呼吸流动量为991.909 t·km-2·a-1,流向碎屑的总流量为523.729 t·km-2·a-1;总能流转化效率为12.8%,来自初级生产者和碎屑的能流转化效率分别为13%和12.3%。研究结果显示:莱州湾朱旺人工鱼礁区主要以底栖生物为主,日本蟳和脉红螺为绝对优势种,中上层鱼类的种类和生物量较少,整个生态系统的成熟度和稳定性较低,食物网连接相对简单且趋于线性结构,系统相对不稳定,抵抗外界干扰能力较差。人工鱼礁区的日本蟳和脉红螺生态容量分别为4.038和2.482 t·km-2,以每年约1.17和0.96 t的捕捞量可持续捕捞10年保持系统平衡;刺参生态容量为50.80 t·km-2,以每年22.38~29.85 t·km-2的放流量并从第3年以每年8.95~11.94 t·km-2规模采捕,4~5年达到其生态容量后停止放流,可继续按原计划采捕5年仍能维持系统稳定。
莱州湾;捕捞策略;人工鱼礁;Ecopath模型;增殖放流;生态容量
随着人类活动干扰和渔业资源衰退加剧,人工鱼礁建设已成为解除近岸海域生态压力及恢复渔业资源的重要手段。为科学评价人工鱼礁建设的生态效果,众多学者通过对人工鱼礁区建设前后或礁区与对照区水文环境、水化学环境、生物环境及游泳生物资源等方面的比较分析,从渔业生境改善、饵料生物增加、经济生物资源量提高等方面探讨了人工鱼礁区的生态效益[1-5]。然而人工鱼礁生态系统作为一种人为引导下的特殊生态系统,其演替过程趋于复杂,并具有特定的物质循环和能量流动特点,因此,有必要引入生态系统特征参数来分析其成熟度及稳定性[6]。
生态通道模型(Ecopath)是一种利用营养动力学原理直接构造水域生态系统结构、描述能量流动以及确定生态参数的能量平衡模式,可以快速反映某一特定水域生态系统实时的状态、特征及营养关系等,从而评价该生态系统的成熟状况,被誉为新一代水域生态系统研究的核心工具[6-7]。近年来,我国学者亦利用Ecopath模型研究了湖泊[8-11]、海洋[7, 12-13]等不同水域生态系统的结构与功能,而国内外利用Ecopath模型研究人工鱼礁生态系统的报道尚不多见。2002年哥伦比亚大学渔业中心首次采用Ecopath模型构建了香港人工鱼礁区生态系统模型[14];2007年李永刚等运用Ecopath模型研究了建礁1年的嵊泗人工鱼礁区生态系统能量流动[15];2009年吴忠鑫等利用Ecopath模型评价了建礁4年的荣成俚岛人工鱼礁区生态系统结构与功能[6]。而利用Ecopath模型对莱州湾人工鱼礁生态系统的研究还未见报道。
莱州湾位于山东半岛西北部,渤海南部,是渤海最大的半封闭性海湾,湾岸属淤泥质平原海岸,海底地形单调平缓,平均水深小于10 m,沿岸有黄河、小清河等多条地表河流入海,带入大量营养盐,饵料生物丰富,是渤海重要渔业资源的主要产卵场、育幼场和索饵场。随着捕捞强度和生态压力增加,莱州湾渔业资源衰退不断加剧,对其重要经济生物的补充恢复及其渔业生境的修复迫在眉睫。2008—2010年山东东方海洋科技股份有限公司在莱州朱旺海域进行了人工鱼礁投放,开展海珍品增养殖和修复渔业资源。本文根据2010—2011年莱州朱旺人工鱼礁区的年际间水化学环境和生物资源逐月调查数据,构建该特定生态系统的Ecopath模型,探讨浅滩平原型海底人工鱼礁生态系统的能量流动规律,分析其成熟度和稳定性,依据模型生态容量估算礁区优势渔获生物日本蟳(Charybdisjaponica)、脉红螺(Rapanavenosa)的生态容量和刺参(Apostichopusjaponucus)的增殖容量,并给出合理的捕捞管理策略,为科学评价人工鱼礁的生态效益,适度捕捞日本蟳、脉红螺和合理增殖刺参等提供理论依据,也为类似海域人工鱼礁建设提供借鉴。
1 礁区概况与研究方法
1.1 礁区概况
莱州朱旺人工鱼礁区位于山东省莱州市莱州湾朱旺港附近海域,该海域盐度全年保持在30~32之间变化较小,水温变化范围为6~28 ℃,水深6~8 m,pH变化范围在7.4~8.1之间。2008年由东方海洋科技股份有限公司开始投放海珍品增殖礁。2008—2010年,共投放石块礁4×106m3、水泥管构件1 000个、扇贝壳礁2×104m3,形成由近岸向远岸的6块人工鱼礁区(S1-S6),礁区面积合计约670 hm2(见图1)。
依据投礁地点的远近将人工鱼礁区分为近岸鱼礁区(S1、S2和S3)和远岸鱼礁区(S4、S5和S6)2个调查区域,每个区域设置3个站位(见表1)。调查网具采用地笼网(规格:网长5 m,网高0.4 m,网宽0.4 m,网目尺寸20 mm),每调查站位放置地笼网3套,放置时间为2 d;渔获物经冷冻保存,带回实验室,进行种类鉴定,并测量各种类的体长、体重等生物学指标;调查期间海区平均水温为17 ℃,平均水深为7 m,平均盐度为31,平均pH为7.75,泥沙质底,各区域间水深无明显差异。由于大规模围海养殖对潮滩的侵占,加之本区域以粉砂潮滩为主,因此历史上仅大叶藻(Zosteramarina)等浅海水生植物繁盛,大型藻类的生物多样性和生物量较少且分布不均匀,生产力极低。
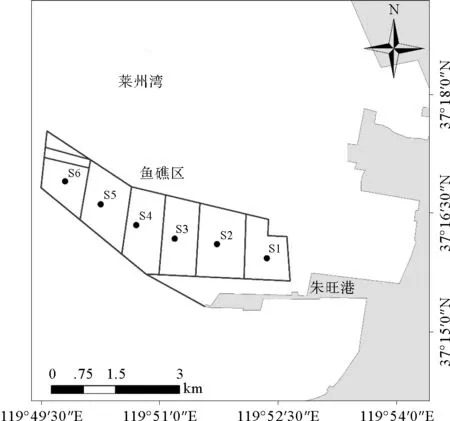
图1 莱州朱旺人工鱼礁区示意图

站点Locality经度Longitude纬度LatitudeS1119°52.35883′E37°15.93567′NS2119°51.72995′E37°16.11413′NS3119°51.19455′E37°16.18212′NS4119°50.71015′E37°16.35209′NS5119°50.25973′E37°16.61554′NS6119°49.80932′E37°16.90448′N
1.2 Ecopath模型建立
1.2.1 功能组划分和数据来源 Ecopath模型由一系列生态关联的功能组组成,所有功能组必须覆盖生态系统能量流动全过程。根据不同生物种类的食性、个体大小和生长特性以及人工鱼礁区海域物种调查情况,并参考同一海域莱州湾相关Ecopath模型的功能组划分标准[16],将莱州湾朱旺人工鱼礁区生态系统定义为14个功能组,基本覆盖了该生态系统能量流动的全过程。依据生态(优势种)和经济(捕捞种)重要性,将许氏平鲉(Sebastesschlegelii)、大泷六线鱼(Hexagrammosotakii)、斑尾复鰕虎鱼(Synechogobiusommaturus)、日本蟳和脉红螺具有重要经济价值和生态功能的单独划分为一个功能组,中上层鱼类、底层鱼类各划分为一个功能组,具体如表2所示。
有机碎屑的生物量参考林群等[12]、张明亮等[16]和杨纪明[17]的报道获得。样品采集严格按照我国《海洋调查规范》(GB12763—2007)进行,浮游生物分别以表层采水法(采水器)和网采法(浅Ⅲ型浮游生物网)定量采集各站位的浮游植物和浮游动物样品,并现场固定(分别用Lugol’s液和Formalin);采集后的样品经浓缩处理后,通过镜检鉴定种类并定量。单位捕捞努力量(CPUE)为每天每套网笼单元的渔获量,单位为g/(net·d),每调查站位放置3套地笼网。根据2010年5月—2011年10月逐月调查数据计算游泳动物生物量[6],其中2010年12月—2011年2月期间由于调查海区结冰未进行取样,胃含物分析获得斑尾复鰕虎鱼食性组成,各功能组的P/B和Q/B值(P为生产量, Q为消耗量, B为生物量)及部分食性分析矩阵参考林群等[12]、张明亮等[16]和杨纪明[ 17-19]的报道获得。
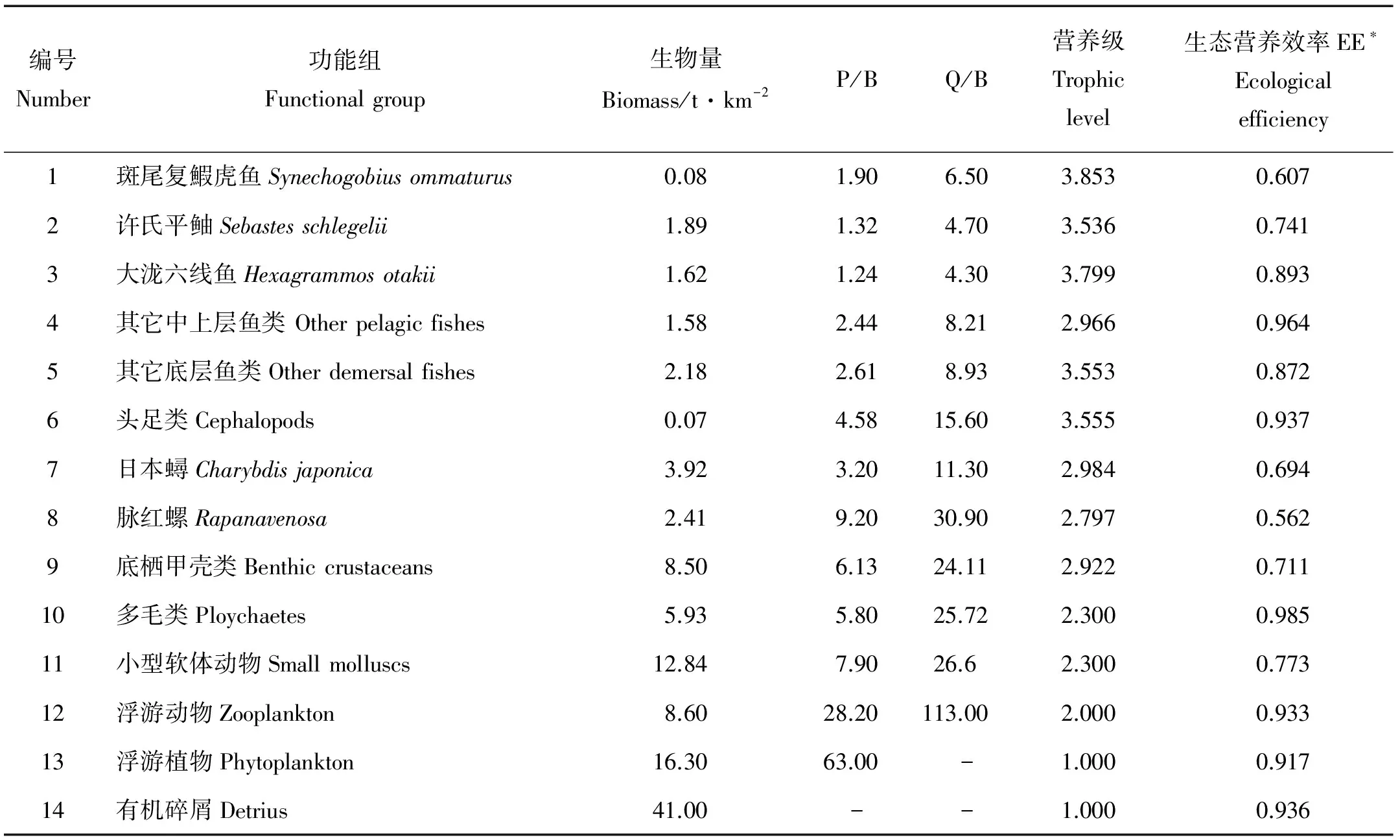
表2 朱旺人工鱼礁区生态系统Ecopath模型功能组估算参数
* 为模型估算值 Estimated parameters; -相应值不存在 No value.
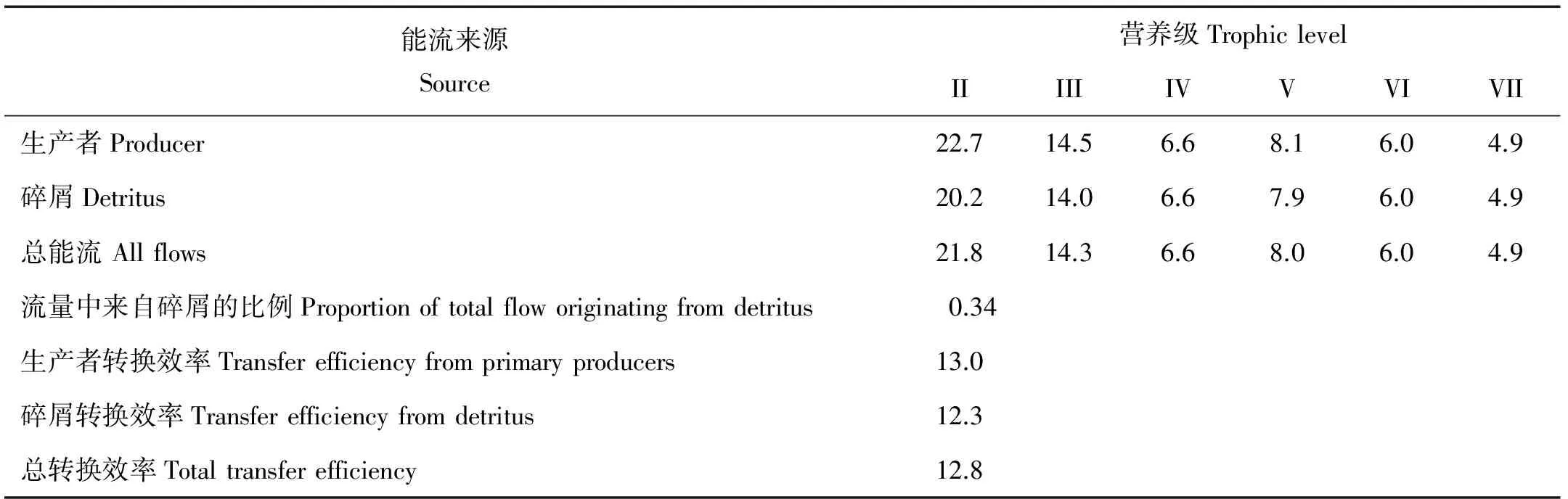
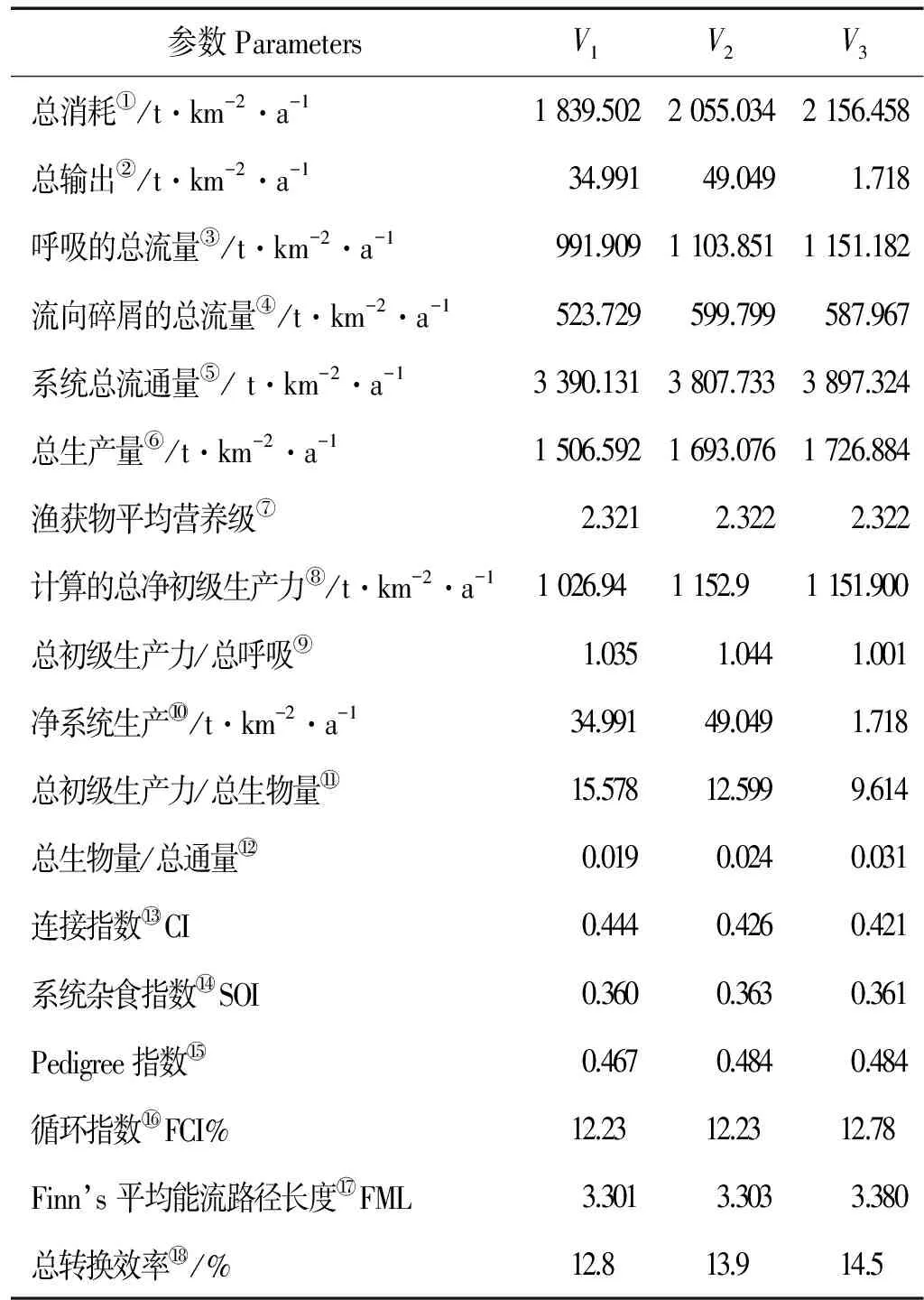
1.2.2 模型调试和总体特征参数 Ecopath模型要求生态系统的输入和输出保持平衡,其基本条件是:0 Pedigree指数是模型敏感性分析的重要指标,用于分析Ecopath模型的数据来源和质量,量化模型输入参数的不确定性[21]。新版的EwE 6.4.3软件在不同参数的数据来源方面有所增加,且0与1的定义根据不同参数有一些变动,不同数据来源置信区间也有所不同,并新增了生态系统关键种的分析模块[22]。混合营养效应 (Mixed trophic impact, MTI)反映功能组之间的相互关系[23]。 总体特征参数是Ecopath模型用来描述系统规模、稳定性和成熟度等特征的指标,主要包括系统总流量(Total system throughput, TST)、总消耗(Total consumption, TC)、净系统生产量(Net system production, NSP)、总呼吸(Total respiratory flows, TR)、连接指数(Connectance index, CI) 和系统杂食指数(System omnivory index, SOI)等[23]。 1.3 日本蟳、脉红螺和刺参生态容量和最低生物量估算方法 生态容量的计算参考Ecopath模型关于贝类养殖容量的估算方法[24]。生态系统中各营养级生物的生物量呈金字塔结构,营养级越高生物量越低,不断提高或降低模型中某种生物的生物量,饵料生物等其它功能组EE随之发生变化,当出现任意功能组EE> 1时,模型失去平衡,则模型不平衡前的生物量即为该种的生态容量;假设该生物EE=1,估算系统平衡条件下该生物的最低生物量。 2.1 莱州朱旺人工鱼礁区生态系统营养级特征 朱旺人工鱼礁区Ecopath模型输入参数和估算参数如表2所示。14个功能组营养级范围为1.000~3.853,占据3个营养级,其中斑尾复鰕虎鱼的营养级最高,达到3.853,其次是大泷六线鱼(3.799)。生态营养效率(EE)为0.562~0.985,其中中上层鱼类和多毛类的EE较高,位于0.96~0.99之间,而脉红螺、日本蟳和斑尾复鰕虎鱼的EE较低,仅为0.56~0.70。所有功能组的P/Q范围为0.226~0.297,运算结果较合理。 2.2 营养级的能量流动和转换效率 图2显示了朱旺人工鱼礁区营养层级间的能量流动。礁区初级生产者的生产量为1 026.94 t·km-2·a-1,其中941.3 t·km-2·a-1被II营养级消耗,占初级生产量的91.7%,85.64 t·km-2·a-1流入碎屑再循环,仅占8.3%。碎屑中来自各个营养级的能量总和为522.8 t·km-2·a-1,来自生产者的能量为85.64 t·km-2·a-1,被摄食的碎屑能量为490.3 t·km-2·a-1,占碎屑总量的80.58%,其余因矿化沉积而脱离系统。整个流入II、III、IV营养级的能量分别占系统总流量的42.84%、9.329%、1.323%。朱旺人工鱼礁生态系统能量流动主要以牧食食物链为主(22.7%)。优势种类脉红螺、日本蟳等位于II、III营养级,使能量流动主要集中于3个营养级之间。总能流转换效率为12.8%,来自初级生产者、碎屑的能流转换效率分别为13.0%和12.3%;来自初级初级生产者和碎屑的能量传输到II、III营养级转换效率都高于其它营养级,且2个营养级总能流分别达到21.8%和14.3%(见表3)。 (P:生产者 Producer,D:有机碎屑 Detritus)图2 朱旺人工鱼礁区生态系统营养层级间能量流动示意图 2.3 功能组间的营养交互关系 混合营养效应(Mixed trophic impact, MTI)用于研究系统内各功能组之间直接和间接的相互作用,估计某一功能组生物量变化对其他功能组生物量的影响。图3为莱州朱旺人工鱼礁区生态系统各功能组间的营养关系,捕食者会直接或间接地消极影响其他功能组,而被捕食者会积极影响其他功能组。有机碎屑、浮游动植物对大部分功能组产生积极效应,如有机碎屑会对底栖甲壳类产生直接的积极效应,浮游动物对中上层鱼类积极作用较强。优势种类许氏平鲉和大泷六线鱼等营养级较高,其生物量增加对底层鱼类和底栖甲壳类等产生消极作用,日本蟳、脉红螺生物量会对底栖甲壳类、浮游动物、小型软体动物等产生较大影响。 表3 朱旺人工鱼礁区生态系统各营养级的转换效率 注:空白:相应值不存在Blank space: no value. (基线上方灰块代表积极影响,基线下方黑块代表消极影响,块状大小表示影响的强弱。Positive impacts were shown above each baseline in grey bar, while negative impacts were shown below the baselines in dark bar, and the rectangular frame size showed the strength of the impact.a:有机碎屑Detrius; b:浮游植物Phytoplankton; c:浮游动物Zooplankton; d:小型软体动物Small mollusca; e:多毛类Ploychaetes; f:底栖甲壳类Benthic crustaceans; g:脉红螺Rapanavenosa;h:日本蟳Charybdisjaponica; i:头足类Cephalopods; j:其他底层鱼类Other demersal fishes; k:其他中上层鱼类Other pelagic fishes; l:大泷六线鱼Hexagrammosotakii; m:许氏平鲉Sebastesschlegelii; n:斑尾复鰕虎鱼Synechogobiusommaturus;Fleet:捕捞船队) 图3 朱旺人工鱼礁区生态系统营养关系图 Fig.3 Mixed trophic impacts analysis for Zhuwang artificial reef ecosystem 利用总体效应(Overall effect,εi)与关键指数(Keystoneness,KSi)对莱州湾朱旺人工鱼礁区生态系统功能群进行排序,如图4所示。关键种是指具有较高总体效应和关键指数(值接近或者大于0)的功能群。许氏平鲉对应的总体效应和关键指数最高,其次是底层鱼类,在维持生态系统稳定性方面发挥重要作用,这也与人工鱼礁区底层游泳动物生物量高于对照区的调查结果相吻合。 2.4 日本蟳、脉红螺生态容量和最低生物量估算 日本蟳和脉红螺是朱旺人工鱼礁区的绝对优势种类,根据调查数据估算两者生物量为3.92和2.41 t·km-2,远高于其他功能组,也是人工鱼礁区的重要捕捞对象。根据模型生态系统容量计算方法,以1%的比例在当前生物量基础上逐步提高日本蟳和脉红螺的生物量,其它功能组生物量维持不变,随着两者生物量的增加,系统各功能组生态效率相应的变动,当两者同时提高到3%时,日本蟳由3.92 t·km-2增加到4.076 8 t·km-2,脉红螺由2.41 t·km-2增加到2.506 4 t·km-2,多毛类能量转化效率EE=1.001>1,模型失去平衡,认为2%比例时两者达到生态容量。因此,日本蟳的生态容量为4.037 6 t·km-2,脉红螺的生态容量为2.482 3 t·km-2。假设日本蟳和脉红螺的生态营养转化效率为EE=1时,估算不影响系统平衡时两者最低生物量为2.464和1.265 t·km-2。 (图中数字为表2所示的功能组编号。Numbers in the graph show the IDs of functional groups in the Table 2.) 图4 朱旺人工鱼礁区生态系统功能群总体效应(εi)与关键指数(KSi)的关系 Fig.4 Relationship between overall effect (εi) and keystoneness (KSi) of functional groups in Zhuwang artificial reef ecosystem 2.5 刺参的增殖容量及其对系统特征的影响 莱州湾朱旺人工鱼礁区主要用于刺参的增养殖,模型构建时未将刺参作为独立功能组,为了反映刺参对生态系统的影响和礁区刺参的增殖生态容量,固定海区刺参每年的投放量,其它功能组相关数据保持不变,将刺参单独作为一个功能组重新加入模型中,采用生态容量方法计算刺参增殖生态容量,结果表明当刺参的生物量为50.80 t·km-2时达到增殖生态容量。 表4为莱州湾朱旺人工鱼礁区在未投放刺参之前(V1)、投放刺参后(V2)和刺参达到增殖生态容量(V3)时生态系统的总体特征参数。对比3个生态系统的总体特征参数,大部分特征参数变化较小,个别参数呈先升高后降低变化,如V1~V3系统总流通量提升了14.99%,V3系统总消耗量分别是V1、V2的1.17和1.05倍。TPP/TR、FCI和FML、CI和SOI表示生态系统成熟度、稳定性和复杂程度,在刺参达到最高增殖生态容量时,TPP/TR由1.035降为1.001,FCI和FML由12.23和3.301提升为12.79和3.381,成熟度和稳定性指标变化幅度较小;CI和SOI由0.444和0.36变为0.421和3.381,复杂程度指标并无明显变化,总能量转化效率逐渐升高。P指数分别为 0.467、0.484和0.484。 表4 刺参投放对莱州湾朱旺人工鱼礁区生态系统总体特征参数的影响 V1:为目前未投放刺参时的系统状态The status of ecosystem have noApostichopusjaponicus;V2:为预测投放刺参后系统状态The predicted status of ecosystem haveApostichopusjaponicus;V3:为预测刺参达到生态容量时系统状态The predicted status at ecological carrying capacity ofApostichopusjaponicas. 3.1 莱州湾朱旺人工鱼礁区生态系统特征 通过调查和分析莱州朱旺人工鱼礁区的渔业资源状况,构建了该礁区的Ecopath模型。并通过新增刺参功能组,探讨了刺参投放对礁区生态系统总体特征参数的影响。模型运行结果表明,未投放刺参、投放刺参和刺参达到生态容量后3个模型P指数位于0.464~0.484之间,与Morissette等[25]给出的全球 150 个Ecopath模型P 指数在0.16~0.68之间相比,表明该模型输入数据的可靠性较好,模型的可信度较高。当前礁区生态系统总能量转化效率为12.8%,高于Linderman关于48个水生群落的平均传递效率(10.1%)以及林群、张明亮关于莱州湾生态系统能量转化效率报道,低于Ryther的研究结果(15%)。系统通过牧食食物链传递的能量高于有机碎屑食物链,总能量转化效率高,流入系统的能量多,各营养级间能量转化效率低;系统稳定性和成熟度指标TPP/TR、FCI、FML、CI和SOI表明系统不成熟稳定;由于莱州湾水深较浅,水温受气温影响较大以及泥沙底质的影响,初级生产力较低,位于同一营养级生物的种类多样性和生物量少,以日本蟳、许氏平鲉、大泷六线鱼和脉红螺等底栖岩礁型生物为主,高营养级的中上层鱼类种类和生物量较少,食物网结构单一,整个系统处于生长发育初期的“幼态”阶段,成熟度低,稳定性差,抗干扰能力弱,易受过度捕捞、环境污染等外界因素的干扰。 3.2 日本蟳和脉红螺的捕捞管理策略 日本蟳和脉红螺是莱州湾朱旺人工鱼礁区主要优势种类和重要经济渔获物,对其采取合理的捕捞管理策略,能够使生态效益和经济效益最大化,且不破坏生态系统平衡。王春琳等[26-27]研究发现日本蟳雌雄比约为1∶1,每年繁殖2次,4—5月产卵量高,8—9月只有部分雌体产卵,平均抱卵量约为8.5×104粒/只;刘洪军等[28]对日本蟳人工育苗和养殖技术研究表明,不同时期成活率不同,从蚤状Ⅰ期幼体到第Ⅱ期幼蟹的成活率为7.62%,从第Ⅴ期幼期到成体的成活率为16.76%。本研究礁区日本蟳生物量为3.92 t·km-2,模型估算日本蟳的最低生物量为2.464 t·km-2,参照日本蟳繁殖生物学研究时的个体体重[28],将性成熟和捕捞体重设为80 g/只,并依据日本蟳平均抱卵量、雌雄比例、性成熟比例以及孵化率和成活率相关研究结果[26-31],制定其10年捕捞管理策略:礁区每年新增日本蟳生物量约为0.03 t·km-2,保持系统平衡条件下,目前日本蟳的可捕捞生物量为1.456 t·km-2,以每年约0.175 6 t·km-2的捕捞量,即每年礁区日本蟳的捕捞量约为1.17 t,可连续捕捞10年不影响系统平衡。 魏利平等[32]研究了脉红螺繁殖方式、繁殖季节、繁殖力等,结果表明脉红螺为雌雄异体,其比例为1∶1,平均每个亲螺在繁殖期内有2.1次交尾活动并产卵;山东沿海脉红螺每年只有一个繁殖期,即6~8月(水温19~26 ℃),产卵盛期在7月(水温22~24 ℃);壳高80~100 mm的亲螺,每个雌螺平均产卵量为75.4万粒。毕进红等[33]对山东沿海脉红螺的性腺发育和生殖周期研究表明,脉红螺配子发育适于9月,分别有14.2%和12.5%的雌雄个体处于性成熟期。本研究礁区脉红螺生物量为2.41 t·km-2,模型估算最低生物量为1.265 t·km-2,参照脉红螺繁殖生物学研究的个体体重[34],将脉红螺个体性成熟和捕捞体重设为≥185 g/个,并依据脉红螺繁殖和生长相关研究数据[32-35],制定其10年捕捞策略:礁区脉红螺的年平均种群自然增殖量约为0.002 t·km-2,礁区系统平衡条件下,目前脉红螺可捕捞的生物量为1.145 t·km-2,每年以0.144 5 t·km-2的捕捞量,即每年礁区脉红螺的捕捞量约为0.96 t,可连续捕捞10年不影响系统平衡。但据东方海洋公司的捕捞日志,2010年日本蟳和脉红螺的捕捞量约为6.567 5和1 t,2011年约为1.033 5和12.12 t,至2012年则骤降为0.325 5和0.385 5 t,日本蟳和脉红螺均已经处于过度捕捞状态,导致2013、2014年日本蟳的捕捞量仅为0.446和0.120 5 t,脉红螺已忽略不计。因此,必须停止捕捞或借助人工增殖放流的手段才能使礁区生态系统重新恢复。 3.3 刺参的底播增殖管理策略 刺参为莱州朱旺人工鱼礁区主要海珍品底播增殖放流对象,具有很高的经济价值。底播增殖放流时,不同规格刺参种苗放流后的成活率差异较大,放流规格越大成活率越高。日本学者认为体长5 cm的幼参,底播增殖放流成活率可达70%左右,回捕率约为40%;山东省海水养殖研究所试验表明,放流体长0.9 cm的稚参,成活率约为0.4%~0.6%,放流体长1cm的参苗,成活率约为12.8%~18.5%;黄海水产研究所试验表明,放流体长2.5 cm的幼参,成活率为20%以上[36]。于东祥等[37]建议放流刺参苗种规格在体长3 cm以上,年成活率可达60%以上,且体长2 cm以上的苗种,海区生长2年即可达到采捕规格。目前莱州朱旺礁区刺参每年的放流量为22.38~29.85 t·km-2,投放规格约3 cm,刺参的生态容量为50.80 t·km-2,根据刺参底播增殖放流的量以及成活率、回捕率相关研究[36-39],按当前的刺参投放量,以70%的成活率和40%的回捕率,制定了9~10年的采捕策略,即每年底播放流量保持不变,从第3年以每年8.95~11.94 t·km-2的生产量开始采捕,4~5年达到其生态容量后停止放流,并可在保持系统平衡的状态下继续采捕5年,然后可根据海域生态系统的实际状况重新制定放流量和采捕量。 随着建礁时间的延长,礁区游泳动物的生物量会逐渐增加,调查礁区资源量时,建议在原有站位的基础上增加定置刺网调查,扩大捕捞对象,获取不同水层游泳动物生物量,丰富调查数据。由于海区地理位置不同以及海区渔业资源结构组成也会随季节发生相应变化,不同海域和不同季节同种游泳动物的饵料并不完全相同,因此,今后针对功能组食性方面,建议利用稳定同位素法对主要功能组进行食性分析,以全面反映功能组的食物来源信息。 [1] 汪振华, 章守宇, 王凯, 等. 三横山人工鱼礁区鱼类和大型无脊椎动物诱集效果初探 [J]. 水产学报, 2010, 34(5): 751-759. Wang Z H, Zhang S Y, Wang K, et al. A preliminary study on fish and macroinvertebrate enhancement artificial reef area around Sanheng Isle, Shengsi, China [J]. Journal of Fisheries of China, 2010, 34(5): 751-759. [2] 吴忠鑫, 张磊, 张秀梅, 等. 荣成俚岛人工鱼礁区游泳动物群落特征及其与主要环境因子的关系 [J]. 生态学报, 2012, 32(21): 6737-6746. Wu Z X, Zhang L, Zhang X M, et al. Nekton community structure and its relationship with main environmental variables in Lidao artificial reef zones of Rongcheng [J]. Acta Ecologica Sinica, 2012, 32(21): 6737-6746. [3] 王伟定, 梁君, 章守宇. 人工鱼礁建设对浙江嵊泗海域营养盐与水质的影响 [J]. 水生生物学报, 2010, 34(1): 78-87. Wang W D, Liang J, Zhang S Y. Influence of artificial reef consrtuction on nutrition and water quality in off-shore area of Shengsi, Zhenjiang [J]. Acta Hydrobiologica Sinica, 2010, 34(1): 78-87. [4] 刘国山, 佟飞, 蔡星媛, 等. 威海双岛海域人工鱼礁区大型海藻夏季群落结构变化 [J]. 中国水产科学, 2014, 21(5): 1010-1019. Liu G S, Tong F, Cai X Y, et al. Variation in the macroalgae community structure during summer in the artificial reef zones of Shuangdao Bay, Weihai, Shandong Province, China [J]. Journal of Fishery Sciences of China, 2014, 21(5): 1010-1019. [5] 佟飞, 张秀梅, 吴忠鑫, 等. 荣成俚岛人工鱼礁区生态系统健康的评价 [J]. 中国海洋大学学报(自然科学版), 2014, 44(4): 29-36. Tong F, Zhang X M, Wu Z X, et al. The ecosystem health assement of artificial reef area in Lidao Island, Rongcheng [J]. Periodical of Ocean University of China, 2014, 44(4): 29-36. [6] 吴忠鑫, 张秀梅, 张磊, 等. 基于Ecopath模型的荣成俚岛人工鱼礁区生态系统结构和功能评价 [J]. 应用生态学报, 2012, 23(10): 2878-2886. Wu Z X, Zhang X M, Zhang L, et al. Structure and function of Lidao artificial reef ecosystem in Rongcheng of Shandong Province, East China: An evaluation based on Ecopath model [J]. Chinese Journal of Applied Ecology, 2012, 23(10): 2878-2886. [7] 陈作志, 邱永松, 贾晓平, 等. 基于Ecopath模型的北部湾生态系统结构和功能 [J]. 中国水产科学, 2008, 15(3): 460-467. Chen Z Z, Qiu Y S, Jia X P, et al. Structure and function of Beibu Gulf ecosystem based on Ecopath model [J]. Journal of Fishery Sciences of China, 2008, 15(3): 460-467. [8] 武震, 贾佩峤, 胡忠军, 等. 基于Ecopath模型分析分水江水库生态系统结构和功能 [J]. 应用生态学报, 2012, 23(3): 812-818. Wu Z, Jia P Q, Hu Z J, et al.Structure and function of Fenshuijiang Reservoir ecosystem based on the analysis with Ecopath model [J]. Chinese Journal of Applied Ecology, 2012, 23(3): 812-818. [9] 李云凯, 刘恩生, 王辉, 等. 基于Ecopath模型的太湖生态系统结构和功能分析 [J]. 应用生态学报, 2014, 25(7): 2033-2040. Li Y K, Liu E S, Wang H, et al. Analysis on the ecosystem structure and function of Lake Taihu based on Ecopath model [J]. Chinese Journal of Applied Ecology, 2014, 25(7): 2033-2040. [10] 刘恩生, 李云凯, 臧日伟, 等. 基于Ecopath模型的巢湖生态系统结构和功能初步分析 [J]. 水产学报, 2014, 38(3): 417-425. Liu E S, Li Y K, Zang R W, et al. A preliminary analysis of the ecosystem structure and functioning of Lake Chaohu based on Ecopath model [J]. Journal of Fisheries of China, 2014, 38(3): 417-425. [11] 黄孝峰, 邴旭文, 陈家长. 基于Ecopath模型的五里湖生态系统营养结构和能量流动研究 [J]. 中国水产科学, 2012, 19(3): 471-481. Huang X F, Bing X W, Chen J C. Energy flow and trophic structure of the Wuli Lake ecosystem based on an ecopath model [J]. Journal of Fishery Sciences of China, 2012, 19(3): 471-481. [12] 林群, 李显森, 李忠义, 等. 基于Ecopath模型的莱州湾中国对虾增殖生态容量 [J]. 中国水产科学, 2013, 24(4): 1131-1140. Lin Q, Li X S, Li Z Y, et al.Ecological carrying capacity of Chinese shrimp stock enhancement in Laizhou Bay of East China based on Ecopath model [J]. Chinese Journal of Applied Ecology, 2013, 24(4): 1131-1140. [13] 李睿, 韩震, 程和秦, 等. 基于Ecopath模型的东海区生物资源能量流动规律的初步研究 [J]. 资源科学, 2010, 32(4): 600-605. Li R, Han Z, Cheng H Q, et al. Apreliminary study on biology resources energy flows based on Ecopath model in the East China Sea [J]. Resources Science, 2010, 32(4): 600-605. [14] Pitcher T, Buchary E, Trujillo P. Spatial simulations of Hong Kong’s marine ecosystem: Ecological and economicforecasting of marine protected areas with human-made reefs[R].//Fisheries CentreResearch Reports. Canada: University of British Columbia, 2002: 1-168. [15] 李永刚, 汪振华, 章守宇. 嵊泗人工鱼礁海区生态系统能量流动模型初探 [J]. 海洋渔业, 2007, 29(3): 226-234. Li Y G, Wang Z H, Zhang S Y. A preliminary approach on the ecosystem model of the artificial reef in Shengsi [J]. Marine Fisheries, 2007, 29(3): 226-234. [16] 张明亮, 冷悦山, 吕振波, 等. 莱州湾三疣梭子蟹生态容量估算 [J]. 海洋渔业, 2013, 35(3): 303-308. Zhang M L, Leng Y S, Lv Z B, et al. Estimating the ecological carrying capacity ofPortunustrituberculatusin Laizhou Bay [J]. Marine Fisheries, 2013, 35(3): 303-308. [17] 杨纪明. 渤海鱼类的食性和营养级研究 [J]. 现代渔业信息, 2001, 16(10): 10-19. Yang J M. A study on food and trophic leves of Bohai Sea Fish [J]. Modern Fisheries Information, 2001, 16(10): 10-19. [18] 杨纪明. 渤海无脊椎动物的食性和营养级研究 [J]. 现代渔业信息, 2001, 16(9): 8-16. Yang J M. A study on food and trophic leves of Bohai Sea Invertebrates [J]. Modern Fisheries Information, 2001, 16(9): 8-16. [19] 杨纪明. 渤海挠足类(Copepoda)的食性和营养级研究 [J]. 现代渔业信息, 2001, 16(6): 6-10. Yang J M. A study on food and trophic leves of Bohai Sea Copepoda [J]. Modern Fisheries Information, 2001, 16(6): 6-10. [20] Christensen V, Walters C J, Pauly D. Ecopath with Ecosim: A user’s guide[M]. Vancouver: Fisheries Centre, University of British Columbia, 2005: 154. [21] Christensen V, Walters C J. Ecopath with Ecosim: Methods, capabilities and limitations [J]. Ecological Mmodelling, 2004, 172(2): 109-139. [22] Libralato S, Christensen V, Pauly D. A method for identifying keystone species in food web models [J]. Ecological modelling, 2006, 195(3): 153-171. [23] Odum E P. The strategy of ecosystem development [J]. Science, 1969, 164: 262-270. [24] Jiang W, Gibbs M T. Predicting the carrying capacity of bivalve shellfish culture using a steady, linear food web model [J]. Aquaculture, 2005, 244(1): 171-185. [25] Morissette L. Complexity, cost and quality of ecosystem models and their impact on resilience: A comparative analysis, with emphasis on marine mammals and the Gulf of St[D]. Lawrence: University of British Columbia, 2007. [26] 王春琳, 薛良义, 刘凤燕, 等. 日本蟳Charybdis(charybdis)Japonica(A. Milne-Edwards) 繁殖生物学的初步研究Ⅰ [J]. 浙江水产学报, 1996, 15(4): 261-266. Wang C L, Xue L Y, Liu F Y, et al. The preliminary study on reproductive biology ofCharybdis(charybdis)Japonica(A. Milne-Edwards)Ⅰ [J]. Journal of Zhejiang College of Fisheries, 1996, 15(4): 261-266. [27] 王春琳, 蒋霞敏, 陈慧群, 等. 日本蟳繁殖生物学的初步研究Ⅱ. 雄性繁殖习性及胚胎发育 [J]. 东海海洋, 2000, 18(1): 43-49. Wang C L, Jiang X M, Chen H Q, et al. A preliminary study on reproductive biology ofCharybdisjaponica(A. Milne-Edwards) Ⅱ. Male breeding characteristics and Embryo growth [J]. Donghai Maring Science, 2000, 18(1): 43-49. [28] 刘洪军, 戴玉蓉, 张富君, 等. 日本蟳人工育苗及养殖技术研究 [J]. 海洋科学, 2000, 24(8): 23-27. Liu H J, Dai Y R, Zhang F J, et al. Studies on the technique for seed rearing and the culture ofCharybdis(charybdis)japonicus[J]. Marine Sciences, 2000, 24(8): 23-27. [29] 姜玉声, 刘庆坤, 李岺, 等. 日本蟳抱卵量与形态参数的相关性分析 [J]. 大连海洋大学学报, 2011, 26(6): 488-492. Jiang Yu S, Liu Q K, Li L, et al. Relationships between morphologic parameters and egg carrying capacity in Asian swimming crabCharybdisjaponica[J]. Journal of Dalian Ocean University, 2011, 26(6): 488-492. [30] 许星鸿, 阎斌伦, 郑家声, 等. 日本蟳的性腺发育和生殖周期 [J]. 海洋湖沼通报, 2010(2): 29-36. Xu X H, Yan B L, Zheng J S, et al. Study on the development of sex gland and reproductive cycle ofCharybdisjaponica(A. Milne-Edwards) [J]. Transactions of Oceanology and Limnology, 2010(2): 29-36. [31] 阎愚, 孙颖民, 宋志乐. 日本蟳幼体发育的研究 [J]. 水产学报,1989, 13(1): 74-79. Yan Y, Sun Y M, Song Z L. The larval development ofCharybdis(charybdis)japonica(A. Milne-Edwards) [J]. Journal of Fisheries of China, 1989, 13(1): 74-79. [32] 魏利平, 邱盛尧, 王宝钢, 等. 脉红螺繁殖生物学的研究 [J]. 水产学报, 1999, 23(2): 150-155. Wei L P, Qiu S Y, Wang B G, et al. Studies on the reproductive biology ofRapanavenosa[J]. Journal of Fisheries of China, 1999, 23(2): 150-155. [33] 毕进红, 李琪, 张志新, 等. 山东沿海脉红螺的性腺发育和生殖周期 [J]. 中国水产科学, 2015, 22(6): 1243-1252. Bi J H, Li Q, Zhang Z X, et al. Gonadal maturation and the reproductive cycle ofRapanavenosaalong the coast of Shandong Province [J]. Journal of Fishery Sciences of China, 2015, 22(6): 1243-1252. [34] 安俊庭. 脉红螺人工苗种繁育与微卫星标记筛选[D]. 青岛: 中国海洋大学, 2014. An J T. Studies on Artifical Breeding and Development of Microsatellite Loci in the Veined Rapa WhelkRapanavenosa[D].Qingdao: Oceea University of China, 2014. [35] 吴耀泉. 莱州湾脉红螺的分布及其壳高与体重的初步分析 [J]. 海洋科学, 1988(6): 39-40. Wu Y Q. Distribution and shell height-weight relation ofRapanavenosavalenciennes in the Laizhou Bay [J]. Marine Sciences, 1988(6): 39-40. [36] 隋锡林. 刺参的放流增殖 [J]. 齐鲁渔业, 2004, 21(6): 16-18. Sui X L. Releasing propagation ofApostichopusjaponicus[J]. Shandong Fisheries, 2004, 21(6): 16-18. [37] 于东祥, 张岩, 陈四清, 等. 刺参增殖的理论和技术(2) [J]. 齐鲁渔业, 2005, 22(7): 54-56. Yu D X, Zhang Y, Chen S Q, et al. The theory and techonlogy of releasing propagation ofApostichopusjaponicusⅡ [J]. Shandong Fisheries, 2005, 22(7): 54-56. [38] 吴耀泉, 崔玉珩, 孙道远, 等. 灵山岛浅海岩礁区刺参增殖放流研究 [J]. 齐鲁渔业, 1996, 13(1): 22-24. Wu Y Q, Cui Y H, Sun D Y, et al. Study on enhancement release ofApostichopusjaponicusin shallow sea rocky area of Lingshandao [J]. Shandong Fisheries, 1996, 13(1): 22-24. [39] 牟绍敦, 李庆彪, 张晓燕. 刺参人工苗种的放流增殖 [J]. 海洋湖沼通报, 1986(3): 44-50. Mu S D, Li Q B, Zhang X Y. Releasing propagation for the artifical seeding ofApostichopusjaponicus(SELENKA) [J]. Transactions of Oceanology and Limnology, 1986(3): 44-50. 责任编辑 朱宝象 The Fishing Strategy of Charybdis japonica and Rapana venosa and the Carrying Capacity ofApostichopusjaponucusin Zhuwang,Laizhou Artificial Reef Ecosystem Based on Ecopath Model YANG Chao-Jie, WU Zhong-Xin, LIU Hong-Yan, ZHANG Pei-Dong,LI Wen-Tao, ZENG Xiao-Qi, ZHANG Xiu-Mei (1.College of Fisheries, Ocean University of China, Qingdao 266003, China; 2.Functional Laboratory of Marine Fisheries Science and Food Production Process, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266235, China) Based on the investigation data of fishery resources in Zhuwang, Laizhou artificial reef zone, from 2010 to 2011 (1 year after deployment), a mass-balanced ecosystem model of an artificial reef zone in Laizhou, Shandong, was established using Ecopath and Ecosim 6.4.3 to develop a management policy forCharybdisjaponica,RapanavenosaandApostichopusjaponucus. Results showed that the functional groups were divided into 3 trophic levels. The total system throughput was 3 390.131 t·km-2·a-1, of which 1 839.502 t·km-2·a-1was for consumption, 523.729 t·km-2·a-1became detritus, and 991.909 t·km-2·a-1was for respiration. The total energy transfer efficiency was 12.8%, and the proportions of the total flow originated from primary producers and detritus, respectively, were 13% and 12.3%. Due to its sandy shoal feature, the primary productivity was extremely low and there was fewer pelagic fish species and lower biomass. The study showed that Zhuwang, Laizhou artificial reef ecosystem, had a relatively low maturity and stability, with a relatively simple food web connection which tended to be linear. The ecosystem was relatively unstable and subjected to external interference. The ecological carrying capacity ofC.japonicaandR.venosawere 4.038 and 2.482 t·km-2·a-1, and sustainable catches of these two species in 10 years was 1.17 t·km-2·a-1and 0.96 t·km-2·a-1in the ecosystem. The ecological carrying capacity ofA.japonucuswas 50.80 t·km-2, which can be released at rates ranging from 22.38 to 29.85 t·km-2·a-1and an annual catch of 8.95-11.94 t·km-2·a-1can be taken two years later. In four to five years, the ecological carrying capacity will be reached and then stop the release. The ecosystem can be kept stable with the catch rate in the next five years. Zhuwang, Laizhou artificial reef ecosystem, was dominated by benthic organisms, of themCharybdisjaponicaandRapanavenosawere the most dominant species. Laizhou Bay; fishing policy; artificial reef;Ecopath mass-balance model; stock enhancement; ecological carrying capacity 海洋公益行业科研专项(201305043;201405010);青岛海洋科学与技术国家实验室鳌山科技创新计划项目(2015ASKJ02)资助 2016-04-06; 2016-05-26 杨超杰(1986-), 男,博士生, 主要从事渔业资源修复研究。 E-mail: yangchaojie1986@163.com ** 通讯作者:E-mail:gaozhang@ouc.edu.cn S931.1 A 1672-5174(2016)11-168-10 10.16441/j.cnki.hdxb.20160112 杨超杰, 吴忠鑫, 刘鸿雁,等. 基于Ecopath模型估算莱州湾朱旺人工鱼礁区日本蟳、脉红螺捕捞策略和刺参增殖生态容量[J]. 中国海洋大学学报(自然科学版), 2016, 46(11): 168-177. YANG Chao-Jie, WU Zhong-Xin, LIU Hong-Yan, et al. The fishing strategy ofCharybdisjaponicaandRapanavenosaand the carrying capacity ofApostichopusjaponucusin Zhuwang, Laizhou artificial reef ecosystem based on Ecopath model [J]. Periodical of Ocean University of China, 2016, 46(11): 168-177. Supported by Special Fund for Scientifc Research in the Public Interest(201305043;201405010);Qingdao National Laboratory for Marine Science and Technology(2015ASKJ02)2 结果与分析



3 讨论
4 结语
